






Abstract
Bispecific antibody MEDI3902, targeting the Pseudomonas aeruginosa type 3 secretion system (PcrV) and Psl exopolysaccharide, is currently in phase 2b development for prevention of nosocomial pneumonia in patients undergoing mechanical ventilation. We surveyed a diverse collection of isolates to study MEDI3902 epitope conservation and protective activity.
P. aeruginosa clinical isolates (n = 913) were collected from diverse patients and geographic locations during 2003–2014. We conducted whole-genome sequencing; performed PcrV and Psl expression analyses via immunoblotting and enzyme-linked immunosorbent assay, respectively; performed crystallography to determine the MEDI3902 PcrV epitope, using anti-PcrV Fab and PcrV components (resolved at 2.8 Å); and evaluated MEDI3902 protective activity against select isolates in vitro and in vivo.
Intact psl operon and pcrV genes were present in 94% and 99% of isolates, respectively, and 99.9% of isolates contained at least one of the genetic elements. Anti-Psl binding was confirmed in tested isolates harboring a complete Psl operon or lacking nonessential psl genes. We identified 46 PcrV variant sequences, and MEDI3902-PcrV contact residues were preserved. MEDI3902 maintained potent in vivo activity against various strains, including strains expressing only a single target.
Psl and PcrV are highly prevalent in global clinical isolates, suggesting MEDI3902 can mediate broad coverage against P. aeruginosa.
The continued spread of antimicrobial resistance among gram-negative bacteria and the now widely accepted recognition of damage to the beneficial microbiome by broad-spectrum antibiotics underscores the need for developing novel strategies for the prevention or treatment of serious bacterial infections. Given their safety profile and extended half-life attributes, pathogen-specific monoclonal antibodies (mAbs) are promising alternatives for the clinician’s armamentarium [1]. Immunotherapeutics could preserve antibiotic use or serve as adjuncts to boost the efficacy of broad-spectrum antibiotics.
Pseudomonas aeruginosa is an opportunistic pathogen that is often antibiotic resistant, making infections with this bacterium difficult to treat. Antibiotic resistance in P. aeruginosa is largely driven by various efflux pumps, reduced permeability of the outer membrane and extraordinary genetic plasticity [2, 3]. Pneumonia due to this pathogen is one of the most clinically relevant manifestations as it is the most frequent cause of gram-negative ICU pneumonia in subjects not undergoing (approximately 13%) or those undergoing (approximately 24%) mechanical ventilation [4–6].
Over the last several decades, many virulence factors and/or surface targets have been sought in the development of a vaccine or immunotherapeutic agent against P. aeruginosa [7–11]. Two surface targets that show considerable promise are PcrV of the type-3 secretion system (T3S) and the Psl exopolysaccharide. PcrV sits at the apex of the T3S needle, which resembles a molecular syringe, and upon contact with host cells is capable of transporting multiple virulence factors directly into host cells [12]. T3S is a major virulence determinant, and its expression in clinical isolates is associated with poor clinical outcomes [13–15]. Psl is an abundantly expressed exopolysaccharide composed of mannose, rhamnose, and glucose, which is important in formation and maintenance of antibiotic recalcitrant biofilms [16–18]. In addition, Psl mediates attachment to epithelial cells and prevents complement deposition on the bacterial surface, allowing P. aeruginosa to escape immune surveillance by host effector cells [19, 20].
Both PcrV and Psl targets are surface accessible and serotype independent, and some mAbs binding to either target have exhibited promising protective activity in multiple murine infection models [21–23]. Binding to PcrV prevents T3S-mediated cytotoxicity, while some mAbs targeting Psl prevent P. aeruginosa attachment to host cells and promote complement-dependent opsonophagocytic killing (OPK) by host effector cells. These complementary mechanisms of action are combined within the bispecific mAb MEDI3902, which was selected based on its enhanced protective activity in comparison to individual parental mAbs or a mAb mixture in a lethal pneumonia model [24].
To better understand the prevalence of the psl operon (expression of Psl is mediated via a 20-kb operon) and the pcrV gene in P. aeruginosa clinical isolates, we performed whole-genome sequencing on 913 isolates collected between 2003–2014 (45 countries) from diverse patient populations and geographical locations. In addition, maintenance of MEDI3902 binding specificity, PcrV epitope conservation and protective activity is reported.
MATERIALS AND METHODS
P. aeruginosa Clinical Isolates
Isolates of P. aeruginosa were analyzed as part of a collection from an international antibiotic resistance surveillance program. Basic demographic data (age, sex, hospital location, sample type, and length of stay) were provided for each isolate by International Health Management Associates (IHMA), using a unique study number that was delinked from any patient identification. Multidrug resistance assignments were performed for 616 isolates from this study as described previously [25], according to breakpoints established by the Clinical and Laboratory Standards Institute (CLSI) and the European Committee on Antimicrobial Susceptibility Testing (EUCAST) [26, 27]. Susceptibility for the remaining 297 isolates was reported previously [28].
Whole-Genome Sequencing and Genetic Analysis
DNA was purified from bacterial cultures via bead beating followed by extraction using a PureLink Genomic DNA Mini Kit (ThermoFisher). Sequencing libraries were prepared by Nextera (Illumina) or Covaris mechanical shearing followed by a NEBNext Ultra DNA Library Prep Kit for Illumina (New England BioLabs). Sequencing was performed via HiSeq 2 for 150 runs or via MiSeq 2 for 250 runs (Illumina). Sequences were de novo assembled in CLC Genomics Workbench, version 9.5.3 (Qiagen). pcrV gene sequences were translated into protein sequences, and amino acid substitutions were identified by using PAO1 NC_002516 pcrV as a reference sequence [3]. Core-genome multialignments were performed using Parsnp and the PAO1 reference NC_002516 [29]. Phylogenetic trees were visualized and annotated using ITOL, version 3 [30].
Psl Operon Gene Content
Psl operon gene content was determined by read mapping to the psl loci from reference NC_002516, using SRST2 [31], and manual inspection of BAM files was performed to confirm gene-null instances.
Detection of PcrV Expression From Clinical Isolates
PcrV expression was evaluated from whole-cell lysates by Western immunoblot analysis after in vitro induction of T3S as described elsewhere [23, 32].
Detection of Psl Expression in Clinical Isolates
Psl expression in clinical isolates was measured by indirect enzyme-linked immunosorbent assays (ELISAs) as previously described [24, 32], using anti-Psl mAb Psl0096 [33], which binds the class I Psl epitope.
Cytotoxicity Assay
Assays were performed as described previously [24]. Briefly, MEDI3902 was added to A549 cells seeded in white 96-well plates (Nunc Nunclon Delta) in Dulbecco’s modified Eagle’s medium plus 10% fetal bovine serum (4 × 104 cells/mL). Log-phase P. aeruginosa clinical isolates capable of expressing ExoU were added at a multiplicity of infection of 10 and incubated for 2 hours at 37° and 5% CO2, and the level of lactate dehydrogenase released from lysed cells was measured. T3S-injectisome and ExoU-positive P. aeruginosa strain 6077 was used as a positive control.
Opsonophagocytosis Assay
Clinical isolates were made luminescent, and assays were performed as previously described [7, 21]. Briefly, assays were performed in 96-well plates, using 0.025 mL of each component: luminescent P. aeruginosa strains from log-phase cultures diluted to approximately 26 cells/mL, diluted baby rabbit serum (1:10), differentiated HL-60 cells (2 × 107 cells/mL), and 11 × 3-fold serial dilutions of MEDI3902. Following incubation for 2 hours at 37°C with shaking at 250 RPM, luminescence was acquired using an Envision Multilabel plate reader (Perkin Elmer) and plotted as percentage killing as compared to a control lacking antibody.
Acute Pneumonia Mouse Model
All procedures were performed in accordance with federal, state, and institutional guidelines and were approved by the MedImmune Institutional Animal Care and Use Committee in an Association for Assessment and Accreditation of Laboratory Animal Care International–accredited facility. Guidelines for humane end points were strictly followed. The acute pneumonia model was performed as described elsewhere [24, 34]. Briefly, 7–8-week-old female BALB/c mice were anesthetized with ketamine and xylazine, followed by intranasal infection with the designated P. aeruginosa strain and infectious inoculum. Animals were monitored closely for up to 5 days after infection. MEDI3902 or an isotype control immunoglobulin G (IgG) were administered 24 hours before infection.
Protein Expression, Purification, Crystallization, Data Collection, and Structure Determination
The Fab fragment of the anti-PcrV arm of MEDI3902 was prepared by papain digestion of parental fully human IgG1ƙ V2L2MD antibody [23]. The digestion mixture was passed over a protein A column (GE Healthcare), and the Fab containing fraction was collected in the flow through. The Fab was further purified to homogeneity by cation exchange and size exclusion chromatography, using HiTrap SP HP and Superdex 200 10/300GL columns (GE Healthcare), respectively. Crystals did not form with full-length PcrV (amino acids 1–294); therefore, we generated a truncated PcrV fragment (PcrVfrag) comprising PcrV amino acids 1–17 fused to amino acids 149–236, which was stably expressed in Escherichia coli and exhibited V2L2MD binding characteristics identical to those of full-length PcrV. Crystallization, data collection, and structure determination are described in the Supplementary Materials. Coordinates and structure factors have been deposited in the Protein Data Bank (accession code 6CYF) and will be released upon publication.
Statistical Analyses
Sample sizes for all animal studies were estimated using the log-rank test with a 5% type I error rate and 80% power and were calculated using nQuery Advisor software. Randomization of animals and treatment groups was determined using a program implemented in Excel. Statistical relationships involving Kaplan-Meier survival curves were performed with GraphPad Prism, version 7.
RESULTS
Patient Demographic Characteristics and P. aeruginosa Characterization
To better understand the global prevalence of PcrV and Psl exopolysaccharide, the molecular targets of MEDI3902, we performed whole-genome sequencing on 913 P. aeruginosa strains collected between 2003 and 2014 from geographically diverse locations. All isolates were derived from body fluids, which included endotracheal aspirates, bronchoalveolar brush samples, lavage samples, sputum, specimens, or urine specimens. The isolates originated from 9 different departments across participating hospitals (Table 1). Approximately 25% or 33% of tested isolates were multidrug resistant according to CLSI or EUCAST breakpoints, respectively.
Strain Demographic Characteristics and Isolate Attributes
| Variable | Overall (n = 913) | Asia (n = 169) | Africa (n = 24) | Europe (n = 322) | Latin America (n = 123) | Middle East (n = 36) | North America (n = 214) | South Pacific (n = 25) |
|---|---|---|---|---|---|---|---|---|
| Year collected, % | ||||||||
| 2003 | 1 (0.1) | … | … | 1 (0.3) | … | … | … | … |
| 2004 | 20 (2.2) | 5 (3) | … | 11 (3.4) | 4 (3.3) | … | … | … |
| 2005 | 13 (1.4) | … | … | 3 (0.9) | 6 (4.9) | … | 4 (1.9) | … |
| 2006 | 17 (1.9) | 7 (4.1) | … | 7 (2.2) | … | … | 3 (1.4) | … |
| 2007 | 35 (3.8) | 6 (3.6) | … | 15 (4.7) | 7 (5.7) | … | 7 (3.3) | … |
| 2008 | 42 (4.6) | 10 (5.9) | … | 24 (7.5) | 8 (6.5) | … | … | … |
| 2009 | 38 (4.2) | 7 (4.1) | … | 21 (6.5) | 6 (4.9) | … | 4 (1.9) | … |
| 2010 | 58 (6.4) | 15 (8.9) | … | 36 (11.2) | … | … | 7 (3.3) | … |
| 2011 | 42 (4.6) | 2 (1.2) | … | 28 (8.7) | 6 (4.9) | … | 6 (2.8) | … |
| 2012 | 61 (6.7) | 6 (3.6) | … | 32 (9.9) | 12 (9.8) | 5 (13.9) | 6 (2.8) | … |
| 2013 | 164 (18) | 13 (7.7) | 10 (41.7) | 39 (12.1) | 21 (17.1) | 8 (22.2) | 68 (31.8) | 5 (20) |
| 2014 | 422 (46.2) | 98 (58) | 14 (58.3) | 105 (32.6) | 53 (43.1) | 23 (63.9) | 109 (50.9) | 20 (80) |
| Facility, % | ||||||||
| Clinic/office | 35 (3.8) | 6 (3.6) | … | 12 (3.7) | 2 (1.6) | … | 15 (7) | … |
| Emergency department | 51 (5.6) | 11 (6.5) | … | 12 (3.7) | 14 (11.4) | 1 (2.8) | 12 (5.6) | 1 (4) |
| General unspecified ICU | 25 (2.7) | 5 (3) | 5 (20.8) | 9 (2.8) | 1 (0.8) | … | 5 (2.3) | … |
| Medicine general | 334 (36.6) | 78 (46.2) | 5 (20.8) | 118 (36.6) | 29 (23.6) | 16 (44.4) | 73 (34.1) | 15 (60) |
| Medicine ICU | 191 (20.9) | 19 (11.2) | 9 (37.5) | 59 (18.3) | 52 (42.3) | 10 (27.8) | 38 (17.8) | 4 (16) |
| Nursing home/rehab | 5 (0.5) | … | … | 1 (0.3) | … | … | 4 (1.9) | … |
| Pediatric general | 27 (3) | 4 (2.4) | 2 (8.3) | 6 (1.9) | 3 (2.4) | 1 (2.8) | 10 (4.7) | 1 (4) |
| Pediatric ICU | 32 (3.5) | 3 (1.8) | … | 7 (2.2) | 2 (1.6) | 2 (5.6) | 17 (7.9) | 1 (4) |
| Surgery general | 86 (9.4) | 20 (11.8) | … | 42 (13) | 8 (6.5) | 2 (5.6) | 14 (6.5) | … |
| Surgery ICU | 92 (10.1) | 18 (10.7) | 3 (12.5) | 35 (10.9) | 10 (8.1) | 3 (8.3) | 20 (9.3) | 3 (12) |
| Other | 3 (0.3) | 2 (1.2) | … | … | … | 1 (2.8) | … | … |
| Missing | 32 (3.5) | 3 (1.8) | … | 21 (6.5) | 2 (1.6) | … | 6 (2.8) | … |
| Body location, % | ||||||||
| Bodily fluids | 82 (9) | 15 (8.9) | … | 42 (13) | 14 (11.4) | … | 10 (4.7) | 1 (4) |
| GI tract | 35 (3.8) | 12 (7.1) | … | 20 (6.2) | … | … | 3 (1.4) | … |
| GU system | 81 (8.9) | 11 (6.5) | … | 42 (13) | 17 (13.8) | … | 11 (5.1) | … |
| Respiratory | ||||||||
| Overall | 715 (78.3) | 131 (77.5) | 24 (100) | 218 (67.7) | 92 (74.8) | 36 (100) | 190 (88.8) | 24 (96) |
| Bronchiole brushing | 23 (2.5) | 1 (0.6) | … | 9 (2.8) | 11 (8.9) | 1 (2.8) | 1 (0.5) | … |
| Bronchioles | 60 (6.6) | 4 (2.4) | … | 34 (10.6) | 6 (4.9) | … | 16 (7.5) | … |
| BAL | 42 (4.6) | … | … | 9 (2.8) | 14 (11.4) | … | 18 (8.4) | 1 (4) |
| Endotracheal aspirate | 129 (14.1) | 15 (8.9) | 8 (33.3) | 27 (8.4) | 29 (23.6) | 14 (38.9) | 24 (11.2) | 12 (48) |
| Lungs | 13 (1.4) | 1 (0.6) | … | 6 (1.9) | 5 (4.1) | … | 1 (0.5) | … |
| Other | 12 (1.3) | 3 (1.8) | 2 (8.3) | 2 (0.6) | 1 (0.8) | 1 (2.8) | 3 (1.4) | … |
| Sinuses | 3 (0.3) | … | … | … | … | … | 3 (1.4) | … |
| Sputum | 351 (38.4) | 104 (61.5) | 10 (41.7) | 92 (28.6) | 20 (16.3) | 15 (41.7) | 99 (46.3) | 11 (44) |
| Trachea | 82 (9) | 3 (1.8) | 4 (16.7) | 39 (12.1) | 6 (4.9) | 5 (13.9) | 25 (11.7) | … |
| Variable | Overall (n = 913) | Asia (n = 169) | Africa (n = 24) | Europe (n = 322) | Latin America (n = 123) | Middle East (n = 36) | North America (n = 214) | South Pacific (n = 25) |
|---|---|---|---|---|---|---|---|---|
| Year collected, % | ||||||||
| 2003 | 1 (0.1) | … | … | 1 (0.3) | … | … | … | … |
| 2004 | 20 (2.2) | 5 (3) | … | 11 (3.4) | 4 (3.3) | … | … | … |
| 2005 | 13 (1.4) | … | … | 3 (0.9) | 6 (4.9) | … | 4 (1.9) | … |
| 2006 | 17 (1.9) | 7 (4.1) | … | 7 (2.2) | … | … | 3 (1.4) | … |
| 2007 | 35 (3.8) | 6 (3.6) | … | 15 (4.7) | 7 (5.7) | … | 7 (3.3) | … |
| 2008 | 42 (4.6) | 10 (5.9) | … | 24 (7.5) | 8 (6.5) | … | … | … |
| 2009 | 38 (4.2) | 7 (4.1) | … | 21 (6.5) | 6 (4.9) | … | 4 (1.9) | … |
| 2010 | 58 (6.4) | 15 (8.9) | … | 36 (11.2) | … | … | 7 (3.3) | … |
| 2011 | 42 (4.6) | 2 (1.2) | … | 28 (8.7) | 6 (4.9) | … | 6 (2.8) | … |
| 2012 | 61 (6.7) | 6 (3.6) | … | 32 (9.9) | 12 (9.8) | 5 (13.9) | 6 (2.8) | … |
| 2013 | 164 (18) | 13 (7.7) | 10 (41.7) | 39 (12.1) | 21 (17.1) | 8 (22.2) | 68 (31.8) | 5 (20) |
| 2014 | 422 (46.2) | 98 (58) | 14 (58.3) | 105 (32.6) | 53 (43.1) | 23 (63.9) | 109 (50.9) | 20 (80) |
| Facility, % | ||||||||
| Clinic/office | 35 (3.8) | 6 (3.6) | … | 12 (3.7) | 2 (1.6) | … | 15 (7) | … |
| Emergency department | 51 (5.6) | 11 (6.5) | … | 12 (3.7) | 14 (11.4) | 1 (2.8) | 12 (5.6) | 1 (4) |
| General unspecified ICU | 25 (2.7) | 5 (3) | 5 (20.8) | 9 (2.8) | 1 (0.8) | … | 5 (2.3) | … |
| Medicine general | 334 (36.6) | 78 (46.2) | 5 (20.8) | 118 (36.6) | 29 (23.6) | 16 (44.4) | 73 (34.1) | 15 (60) |
| Medicine ICU | 191 (20.9) | 19 (11.2) | 9 (37.5) | 59 (18.3) | 52 (42.3) | 10 (27.8) | 38 (17.8) | 4 (16) |
| Nursing home/rehab | 5 (0.5) | … | … | 1 (0.3) | … | … | 4 (1.9) | … |
| Pediatric general | 27 (3) | 4 (2.4) | 2 (8.3) | 6 (1.9) | 3 (2.4) | 1 (2.8) | 10 (4.7) | 1 (4) |
| Pediatric ICU | 32 (3.5) | 3 (1.8) | … | 7 (2.2) | 2 (1.6) | 2 (5.6) | 17 (7.9) | 1 (4) |
| Surgery general | 86 (9.4) | 20 (11.8) | … | 42 (13) | 8 (6.5) | 2 (5.6) | 14 (6.5) | … |
| Surgery ICU | 92 (10.1) | 18 (10.7) | 3 (12.5) | 35 (10.9) | 10 (8.1) | 3 (8.3) | 20 (9.3) | 3 (12) |
| Other | 3 (0.3) | 2 (1.2) | … | … | … | 1 (2.8) | … | … |
| Missing | 32 (3.5) | 3 (1.8) | … | 21 (6.5) | 2 (1.6) | … | 6 (2.8) | … |
| Body location, % | ||||||||
| Bodily fluids | 82 (9) | 15 (8.9) | … | 42 (13) | 14 (11.4) | … | 10 (4.7) | 1 (4) |
| GI tract | 35 (3.8) | 12 (7.1) | … | 20 (6.2) | … | … | 3 (1.4) | … |
| GU system | 81 (8.9) | 11 (6.5) | … | 42 (13) | 17 (13.8) | … | 11 (5.1) | … |
| Respiratory | ||||||||
| Overall | 715 (78.3) | 131 (77.5) | 24 (100) | 218 (67.7) | 92 (74.8) | 36 (100) | 190 (88.8) | 24 (96) |
| Bronchiole brushing | 23 (2.5) | 1 (0.6) | … | 9 (2.8) | 11 (8.9) | 1 (2.8) | 1 (0.5) | … |
| Bronchioles | 60 (6.6) | 4 (2.4) | … | 34 (10.6) | 6 (4.9) | … | 16 (7.5) | … |
| BAL | 42 (4.6) | … | … | 9 (2.8) | 14 (11.4) | … | 18 (8.4) | 1 (4) |
| Endotracheal aspirate | 129 (14.1) | 15 (8.9) | 8 (33.3) | 27 (8.4) | 29 (23.6) | 14 (38.9) | 24 (11.2) | 12 (48) |
| Lungs | 13 (1.4) | 1 (0.6) | … | 6 (1.9) | 5 (4.1) | … | 1 (0.5) | … |
| Other | 12 (1.3) | 3 (1.8) | 2 (8.3) | 2 (0.6) | 1 (0.8) | 1 (2.8) | 3 (1.4) | … |
| Sinuses | 3 (0.3) | … | … | … | … | … | 3 (1.4) | … |
| Sputum | 351 (38.4) | 104 (61.5) | 10 (41.7) | 92 (28.6) | 20 (16.3) | 15 (41.7) | 99 (46.3) | 11 (44) |
| Trachea | 82 (9) | 3 (1.8) | 4 (16.7) | 39 (12.1) | 6 (4.9) | 5 (13.9) | 25 (11.7) | … |
Abbreviations: BAL, bronchoalveolar lavage; GI, gastrointestinal; GU, genitourinary; ICU, intensive care unit.
Strain Demographic Characteristics and Isolate Attributes
| Variable | Overall (n = 913) | Asia (n = 169) | Africa (n = 24) | Europe (n = 322) | Latin America (n = 123) | Middle East (n = 36) | North America (n = 214) | South Pacific (n = 25) |
|---|---|---|---|---|---|---|---|---|
| Year collected, % | ||||||||
| 2003 | 1 (0.1) | … | … | 1 (0.3) | … | … | … | … |
| 2004 | 20 (2.2) | 5 (3) | … | 11 (3.4) | 4 (3.3) | … | … | … |
| 2005 | 13 (1.4) | … | … | 3 (0.9) | 6 (4.9) | … | 4 (1.9) | … |
| 2006 | 17 (1.9) | 7 (4.1) | … | 7 (2.2) | … | … | 3 (1.4) | … |
| 2007 | 35 (3.8) | 6 (3.6) | … | 15 (4.7) | 7 (5.7) | … | 7 (3.3) | … |
| 2008 | 42 (4.6) | 10 (5.9) | … | 24 (7.5) | 8 (6.5) | … | … | … |
| 2009 | 38 (4.2) | 7 (4.1) | … | 21 (6.5) | 6 (4.9) | … | 4 (1.9) | … |
| 2010 | 58 (6.4) | 15 (8.9) | … | 36 (11.2) | … | … | 7 (3.3) | … |
| 2011 | 42 (4.6) | 2 (1.2) | … | 28 (8.7) | 6 (4.9) | … | 6 (2.8) | … |
| 2012 | 61 (6.7) | 6 (3.6) | … | 32 (9.9) | 12 (9.8) | 5 (13.9) | 6 (2.8) | … |
| 2013 | 164 (18) | 13 (7.7) | 10 (41.7) | 39 (12.1) | 21 (17.1) | 8 (22.2) | 68 (31.8) | 5 (20) |
| 2014 | 422 (46.2) | 98 (58) | 14 (58.3) | 105 (32.6) | 53 (43.1) | 23 (63.9) | 109 (50.9) | 20 (80) |
| Facility, % | ||||||||
| Clinic/office | 35 (3.8) | 6 (3.6) | … | 12 (3.7) | 2 (1.6) | … | 15 (7) | … |
| Emergency department | 51 (5.6) | 11 (6.5) | … | 12 (3.7) | 14 (11.4) | 1 (2.8) | 12 (5.6) | 1 (4) |
| General unspecified ICU | 25 (2.7) | 5 (3) | 5 (20.8) | 9 (2.8) | 1 (0.8) | … | 5 (2.3) | … |
| Medicine general | 334 (36.6) | 78 (46.2) | 5 (20.8) | 118 (36.6) | 29 (23.6) | 16 (44.4) | 73 (34.1) | 15 (60) |
| Medicine ICU | 191 (20.9) | 19 (11.2) | 9 (37.5) | 59 (18.3) | 52 (42.3) | 10 (27.8) | 38 (17.8) | 4 (16) |
| Nursing home/rehab | 5 (0.5) | … | … | 1 (0.3) | … | … | 4 (1.9) | … |
| Pediatric general | 27 (3) | 4 (2.4) | 2 (8.3) | 6 (1.9) | 3 (2.4) | 1 (2.8) | 10 (4.7) | 1 (4) |
| Pediatric ICU | 32 (3.5) | 3 (1.8) | … | 7 (2.2) | 2 (1.6) | 2 (5.6) | 17 (7.9) | 1 (4) |
| Surgery general | 86 (9.4) | 20 (11.8) | … | 42 (13) | 8 (6.5) | 2 (5.6) | 14 (6.5) | … |
| Surgery ICU | 92 (10.1) | 18 (10.7) | 3 (12.5) | 35 (10.9) | 10 (8.1) | 3 (8.3) | 20 (9.3) | 3 (12) |
| Other | 3 (0.3) | 2 (1.2) | … | … | … | 1 (2.8) | … | … |
| Missing | 32 (3.5) | 3 (1.8) | … | 21 (6.5) | 2 (1.6) | … | 6 (2.8) | … |
| Body location, % | ||||||||
| Bodily fluids | 82 (9) | 15 (8.9) | … | 42 (13) | 14 (11.4) | … | 10 (4.7) | 1 (4) |
| GI tract | 35 (3.8) | 12 (7.1) | … | 20 (6.2) | … | … | 3 (1.4) | … |
| GU system | 81 (8.9) | 11 (6.5) | … | 42 (13) | 17 (13.8) | … | 11 (5.1) | … |
| Respiratory | ||||||||
| Overall | 715 (78.3) | 131 (77.5) | 24 (100) | 218 (67.7) | 92 (74.8) | 36 (100) | 190 (88.8) | 24 (96) |
| Bronchiole brushing | 23 (2.5) | 1 (0.6) | … | 9 (2.8) | 11 (8.9) | 1 (2.8) | 1 (0.5) | … |
| Bronchioles | 60 (6.6) | 4 (2.4) | … | 34 (10.6) | 6 (4.9) | … | 16 (7.5) | … |
| BAL | 42 (4.6) | … | … | 9 (2.8) | 14 (11.4) | … | 18 (8.4) | 1 (4) |
| Endotracheal aspirate | 129 (14.1) | 15 (8.9) | 8 (33.3) | 27 (8.4) | 29 (23.6) | 14 (38.9) | 24 (11.2) | 12 (48) |
| Lungs | 13 (1.4) | 1 (0.6) | … | 6 (1.9) | 5 (4.1) | … | 1 (0.5) | … |
| Other | 12 (1.3) | 3 (1.8) | 2 (8.3) | 2 (0.6) | 1 (0.8) | 1 (2.8) | 3 (1.4) | … |
| Sinuses | 3 (0.3) | … | … | … | … | … | 3 (1.4) | … |
| Sputum | 351 (38.4) | 104 (61.5) | 10 (41.7) | 92 (28.6) | 20 (16.3) | 15 (41.7) | 99 (46.3) | 11 (44) |
| Trachea | 82 (9) | 3 (1.8) | 4 (16.7) | 39 (12.1) | 6 (4.9) | 5 (13.9) | 25 (11.7) | … |
| Variable | Overall (n = 913) | Asia (n = 169) | Africa (n = 24) | Europe (n = 322) | Latin America (n = 123) | Middle East (n = 36) | North America (n = 214) | South Pacific (n = 25) |
|---|---|---|---|---|---|---|---|---|
| Year collected, % | ||||||||
| 2003 | 1 (0.1) | … | … | 1 (0.3) | … | … | … | … |
| 2004 | 20 (2.2) | 5 (3) | … | 11 (3.4) | 4 (3.3) | … | … | … |
| 2005 | 13 (1.4) | … | … | 3 (0.9) | 6 (4.9) | … | 4 (1.9) | … |
| 2006 | 17 (1.9) | 7 (4.1) | … | 7 (2.2) | … | … | 3 (1.4) | … |
| 2007 | 35 (3.8) | 6 (3.6) | … | 15 (4.7) | 7 (5.7) | … | 7 (3.3) | … |
| 2008 | 42 (4.6) | 10 (5.9) | … | 24 (7.5) | 8 (6.5) | … | … | … |
| 2009 | 38 (4.2) | 7 (4.1) | … | 21 (6.5) | 6 (4.9) | … | 4 (1.9) | … |
| 2010 | 58 (6.4) | 15 (8.9) | … | 36 (11.2) | … | … | 7 (3.3) | … |
| 2011 | 42 (4.6) | 2 (1.2) | … | 28 (8.7) | 6 (4.9) | … | 6 (2.8) | … |
| 2012 | 61 (6.7) | 6 (3.6) | … | 32 (9.9) | 12 (9.8) | 5 (13.9) | 6 (2.8) | … |
| 2013 | 164 (18) | 13 (7.7) | 10 (41.7) | 39 (12.1) | 21 (17.1) | 8 (22.2) | 68 (31.8) | 5 (20) |
| 2014 | 422 (46.2) | 98 (58) | 14 (58.3) | 105 (32.6) | 53 (43.1) | 23 (63.9) | 109 (50.9) | 20 (80) |
| Facility, % | ||||||||
| Clinic/office | 35 (3.8) | 6 (3.6) | … | 12 (3.7) | 2 (1.6) | … | 15 (7) | … |
| Emergency department | 51 (5.6) | 11 (6.5) | … | 12 (3.7) | 14 (11.4) | 1 (2.8) | 12 (5.6) | 1 (4) |
| General unspecified ICU | 25 (2.7) | 5 (3) | 5 (20.8) | 9 (2.8) | 1 (0.8) | … | 5 (2.3) | … |
| Medicine general | 334 (36.6) | 78 (46.2) | 5 (20.8) | 118 (36.6) | 29 (23.6) | 16 (44.4) | 73 (34.1) | 15 (60) |
| Medicine ICU | 191 (20.9) | 19 (11.2) | 9 (37.5) | 59 (18.3) | 52 (42.3) | 10 (27.8) | 38 (17.8) | 4 (16) |
| Nursing home/rehab | 5 (0.5) | … | … | 1 (0.3) | … | … | 4 (1.9) | … |
| Pediatric general | 27 (3) | 4 (2.4) | 2 (8.3) | 6 (1.9) | 3 (2.4) | 1 (2.8) | 10 (4.7) | 1 (4) |
| Pediatric ICU | 32 (3.5) | 3 (1.8) | … | 7 (2.2) | 2 (1.6) | 2 (5.6) | 17 (7.9) | 1 (4) |
| Surgery general | 86 (9.4) | 20 (11.8) | … | 42 (13) | 8 (6.5) | 2 (5.6) | 14 (6.5) | … |
| Surgery ICU | 92 (10.1) | 18 (10.7) | 3 (12.5) | 35 (10.9) | 10 (8.1) | 3 (8.3) | 20 (9.3) | 3 (12) |
| Other | 3 (0.3) | 2 (1.2) | … | … | … | 1 (2.8) | … | … |
| Missing | 32 (3.5) | 3 (1.8) | … | 21 (6.5) | 2 (1.6) | … | 6 (2.8) | … |
| Body location, % | ||||||||
| Bodily fluids | 82 (9) | 15 (8.9) | … | 42 (13) | 14 (11.4) | … | 10 (4.7) | 1 (4) |
| GI tract | 35 (3.8) | 12 (7.1) | … | 20 (6.2) | … | … | 3 (1.4) | … |
| GU system | 81 (8.9) | 11 (6.5) | … | 42 (13) | 17 (13.8) | … | 11 (5.1) | … |
| Respiratory | ||||||||
| Overall | 715 (78.3) | 131 (77.5) | 24 (100) | 218 (67.7) | 92 (74.8) | 36 (100) | 190 (88.8) | 24 (96) |
| Bronchiole brushing | 23 (2.5) | 1 (0.6) | … | 9 (2.8) | 11 (8.9) | 1 (2.8) | 1 (0.5) | … |
| Bronchioles | 60 (6.6) | 4 (2.4) | … | 34 (10.6) | 6 (4.9) | … | 16 (7.5) | … |
| BAL | 42 (4.6) | … | … | 9 (2.8) | 14 (11.4) | … | 18 (8.4) | 1 (4) |
| Endotracheal aspirate | 129 (14.1) | 15 (8.9) | 8 (33.3) | 27 (8.4) | 29 (23.6) | 14 (38.9) | 24 (11.2) | 12 (48) |
| Lungs | 13 (1.4) | 1 (0.6) | … | 6 (1.9) | 5 (4.1) | … | 1 (0.5) | … |
| Other | 12 (1.3) | 3 (1.8) | 2 (8.3) | 2 (0.6) | 1 (0.8) | 1 (2.8) | 3 (1.4) | … |
| Sinuses | 3 (0.3) | … | … | … | … | … | 3 (1.4) | … |
| Sputum | 351 (38.4) | 104 (61.5) | 10 (41.7) | 92 (28.6) | 20 (16.3) | 15 (41.7) | 99 (46.3) | 11 (44) |
| Trachea | 82 (9) | 3 (1.8) | 4 (16.7) | 39 (12.1) | 6 (4.9) | 5 (13.9) | 25 (11.7) | … |
Abbreviations: BAL, bronchoalveolar lavage; GI, gastrointestinal; GU, genitourinary; ICU, intensive care unit.
psl Operon Genes Are Conserved in Diverse P. aeruginosa Isolates
Sequence analyses revealed that 94% of isolates contained a complete psl operon, while only 28 of 913 isolates lacked all psl A-O genes. Twenty-six isolates were null in ≥1 psl operon gene (Figure 1A and 1B). Representative isolates were tested in a whole-cell Psl ELISA to identify which psl operon genes are required for Psl synthesis and binding by the anti-Psl mAb Psl0096 (complementarity-determining regions [CDRs] contained within MEDI3902). As expected, isolates that were either completely operon null, deficient for pslA, or lacking in genes for pslC, pslH, pslI, and pslE-pslO were negative for Psl expression (Figure 1B), indicating the requirement of these genes for Psl expression. Selected isolates that were null for either of the Psl operon genes pslM or pslN-pslO were positive for Psl expression, indicating that these genes are not required for Psl expression.
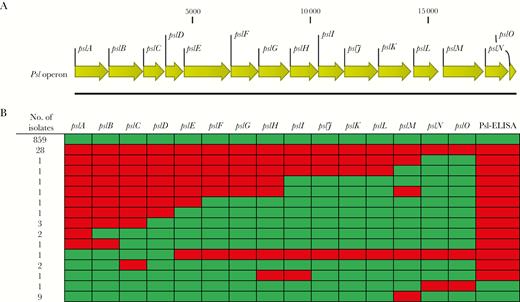
The presence of the psl operon was determined by analysis of whole-genome sequencing data. A, Schematic representation of the psl operon. B, Genotypic overview of the psl operon among the 913 Pseudomonas aeruginosa clinical isolates. Green squares mark the presence and red squares the absence of a full-length gene. Similarly, green and red squares mark the presence and absence, respectively, of anti-Psl reactivity from representative isolates that were evaluated by an enzyme-linked immunosorbent assay.
The pcrV Gene and PcrV Protein Are Conserved in Diverse P. aeruginosa Isolates
A full-length pcrV gene was present in the majority of isolates (99% [902 of 913]), demonstrating high prevalence among P. aeruginosa strains. Using PAO1 as a reference strain, PcrV amino acid sequence analyses of the clinical isolates revealed substitutions at 35 positions, which corresponded to 46 distinct PcrV full-length protein subtypes (Table 2). The most divergent PcrV protein sequences (containing S21P, K118E, D119A, S146G, V172L, D188G, and S225R substitutions) were 96.3% identical (288 of 299 residues) to the PAO1 PcrV reference amino acid sequence (Table 2).
Pseudomonas aeruginosa Type 3 Secretion System (PcrV) Amino Acid Variation
| Isolates, No. | Subtype | Percentage | Amino Acid Position, Reference Sequence | ||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 5, N | 6, L | 9, A | 10, R | 21, S | 27, A | 30, E | 33, L | 53, A | 68, S | 69, A | 88, P | 106, H | 112, D | 118, K | 119, D | 123, T | 143, K | 146, S | 154, A | 172, V | 181, V | 182, G | 184, P | 188, D | 199, D | 209, D | 220, E | 225, S | 229, P | 236, P | 237, V | 256, N | 275, A | 285, S | |||
| 431 | 1 | 47.8 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 141 | 2 | 15.6 | - | F | G | - | P | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | R | - | - | - | - | - | - |
| 62 | 3 | 6.9 | - | F | - | - | P | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | K | - | - | - | - | - | - |
| 59 | 4 | 6.5 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | R | - | - | - | - | - | - |
| 50 | 5 | 5.5 | - | F | G | - | P | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | K | - | - | - | - | - | - |
| 27 | 6 | 3.0 | - | - | - | - | - | - | - | - | - | - | - | - | Y | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 19 | 7 | 2.1 | - | F | G | - | P | - | - | M | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | R | - | - | - | - | - | - |
| 15 | 8 | 1.7 | - | F | - | - | P | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | R | - | - | - | - | - | - |
| 12 | 9 | 1.3 | - | F | G | - | P | - | K | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | R | - | - | - | - | - | - |
| 10 | 10 | 1.1 | - | - | G | - | P | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 9 | 11 | 1.0 | - | F | G | - | P | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 9 | 12 | 1.0 | - | F | G | - | P | - | K | - | - | - | - | - | R | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | R | - | - | - | - | - | - |
| 6 | 13 | 0.7 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | G | - | - | - | - | - | - |
| 6 | 14 | 0.7 | - | - | - | - | P | - | - | - | - | - | - | - | - | - | E | A | - | - | G | - | L | - | - | - | G | G | N | - | R | - | - | - | - | - | - |
| 5 | 15 | 0.6 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | I |
| 3 | 16 | 0.3 | - | F | S | - | P | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | R | - | - | - | - | - | - |
| 2 | 17 | 0.2 | - | - | - | - | - | G | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 2 | 18 | 0.2 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | V | I |
| 2 | 19 | 0.2 | - | - | - | - | P | - | - | - | - | - | - | - | - | N | - | - | - | - | - | - | - | - | - | - | - | - | - | - | K | - | - | - | - | - | - |
| 2 | 20 | 0.2 | - | F | S | - | Q | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | R | - | - | - | - | - | - |
| 2 | 21 | 0.2 | - | F | G | - | P | - | K | - | - | - | - | - | Y | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 2 | 22 | 0.2 | - | F | G | - | P | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | S | - | - | - | - | R | - | - | - | - | - | - |
| 2 | 23 | 0.2 | - | - | - | - | P | - | - | - | - | - | - | - | - | - | E | A | - | - | G | - | L | - | - | - | G | - | - | - | R | - | - | - | - | - | - |
| 2 | 24 | 0.2 | - | - | - | - | P | - | - | - | - | - | - | - | - | - | E | A | - | - | G | - | L | - | - | - | G | G | - | - | R | - | - | - | - | - | - |
| 1 | 25 | 0.1 | - | - | V | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 1 | 26 | 0.1 | - | - | - | S | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 1 | 27 | 0.1 | - | - | - | - | - | - | - | - | - | - | P | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 1 | 28 | 0.1 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | R | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 1 | 29 | 0.1 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | S | - | - | - | - | - |
| 1 | 30 | 0.1 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | S | - | - | - | - |
| 1 | 31 | 0.1 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | D | - | - | - | - | - | - | - | - | - | - | - | - |
| 1 | 32 | 0.1 | - | - | - | - | - | - | - | - | - | - | - | L | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 1 | 33 | 0.1 | - | - | - | - | - | - | - | - | - | - | - | - | Y | - | - | - | A | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 1 | 34 | 0.1 | - | F | - | - | P | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 1 | 35 | 0.1 | - | - | G | - | P | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | R | - | - | - | - | - | - |
| 1 | 36 | 0.1 | S | F | - | - | P | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | R | - | - | - | - | - | - |
| 1 | 37 | 0.1 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | L | A | - | - | G | - | - | - | R | - | - | - | - | - | - |
| 1 | 38 | 0.1 | - | F | G | - | P | - | - | M | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 1 | 39 | 0.1 | - | F | - | - | P | - | - | - | V | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | K | - | - | - | - | - | - |
| 1 | 40 | 0.1 | - | F | - | - | P | - | - | - | - | T | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | R | - | - | - | - | - | - |
| 1 | 41 | 0.1 | - | F | - | - | P | - | - | - | - | - | - | - | R | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | R | - | - | - | - | - | - |
| 1 | 42 | 0.1 | - | F | - | - | P | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | R | - | - | I | - | - | - |
| 1 | 43 | 0.1 | - | F | - | - | P | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | K | - | - | - | S | - | - |
| 1 | 44 | 0.1 | - | F | G | - | P | - | - | - | - | - | - | - | - | - | - | - | - | - | - | V | - | - | - | - | - | - | - | - | R | - | - | - | - | - | - |
| 1 | 45 | 0.1 | - | F | G | - | P | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | D | - | - | - | - | - | R | - | - | - | - | - | - |
| 1 | 46 | 0.1 | - | F | G | - | P | - | K | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | K | R | - | - | - | - | - | - |
| Isolates, No. | Subtype | Percentage | Amino Acid Position, Reference Sequence | ||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 5, N | 6, L | 9, A | 10, R | 21, S | 27, A | 30, E | 33, L | 53, A | 68, S | 69, A | 88, P | 106, H | 112, D | 118, K | 119, D | 123, T | 143, K | 146, S | 154, A | 172, V | 181, V | 182, G | 184, P | 188, D | 199, D | 209, D | 220, E | 225, S | 229, P | 236, P | 237, V | 256, N | 275, A | 285, S | |||
| 431 | 1 | 47.8 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 141 | 2 | 15.6 | - | F | G | - | P | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | R | - | - | - | - | - | - |
| 62 | 3 | 6.9 | - | F | - | - | P | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | K | - | - | - | - | - | - |
| 59 | 4 | 6.5 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | R | - | - | - | - | - | - |
| 50 | 5 | 5.5 | - | F | G | - | P | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | K | - | - | - | - | - | - |
| 27 | 6 | 3.0 | - | - | - | - | - | - | - | - | - | - | - | - | Y | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 19 | 7 | 2.1 | - | F | G | - | P | - | - | M | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | R | - | - | - | - | - | - |
| 15 | 8 | 1.7 | - | F | - | - | P | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | R | - | - | - | - | - | - |
| 12 | 9 | 1.3 | - | F | G | - | P | - | K | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | R | - | - | - | - | - | - |
| 10 | 10 | 1.1 | - | - | G | - | P | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 9 | 11 | 1.0 | - | F | G | - | P | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 9 | 12 | 1.0 | - | F | G | - | P | - | K | - | - | - | - | - | R | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | R | - | - | - | - | - | - |
| 6 | 13 | 0.7 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | G | - | - | - | - | - | - |
| 6 | 14 | 0.7 | - | - | - | - | P | - | - | - | - | - | - | - | - | - | E | A | - | - | G | - | L | - | - | - | G | G | N | - | R | - | - | - | - | - | - |
| 5 | 15 | 0.6 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | I |
| 3 | 16 | 0.3 | - | F | S | - | P | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | R | - | - | - | - | - | - |
| 2 | 17 | 0.2 | - | - | - | - | - | G | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 2 | 18 | 0.2 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | V | I |
| 2 | 19 | 0.2 | - | - | - | - | P | - | - | - | - | - | - | - | - | N | - | - | - | - | - | - | - | - | - | - | - | - | - | - | K | - | - | - | - | - | - |
| 2 | 20 | 0.2 | - | F | S | - | Q | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | R | - | - | - | - | - | - |
| 2 | 21 | 0.2 | - | F | G | - | P | - | K | - | - | - | - | - | Y | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 2 | 22 | 0.2 | - | F | G | - | P | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | S | - | - | - | - | R | - | - | - | - | - | - |
| 2 | 23 | 0.2 | - | - | - | - | P | - | - | - | - | - | - | - | - | - | E | A | - | - | G | - | L | - | - | - | G | - | - | - | R | - | - | - | - | - | - |
| 2 | 24 | 0.2 | - | - | - | - | P | - | - | - | - | - | - | - | - | - | E | A | - | - | G | - | L | - | - | - | G | G | - | - | R | - | - | - | - | - | - |
| 1 | 25 | 0.1 | - | - | V | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 1 | 26 | 0.1 | - | - | - | S | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 1 | 27 | 0.1 | - | - | - | - | - | - | - | - | - | - | P | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 1 | 28 | 0.1 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | R | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 1 | 29 | 0.1 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | S | - | - | - | - | - |
| 1 | 30 | 0.1 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | S | - | - | - | - |
| 1 | 31 | 0.1 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | D | - | - | - | - | - | - | - | - | - | - | - | - |
| 1 | 32 | 0.1 | - | - | - | - | - | - | - | - | - | - | - | L | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 1 | 33 | 0.1 | - | - | - | - | - | - | - | - | - | - | - | - | Y | - | - | - | A | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 1 | 34 | 0.1 | - | F | - | - | P | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 1 | 35 | 0.1 | - | - | G | - | P | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | R | - | - | - | - | - | - |
| 1 | 36 | 0.1 | S | F | - | - | P | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | R | - | - | - | - | - | - |
| 1 | 37 | 0.1 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | L | A | - | - | G | - | - | - | R | - | - | - | - | - | - |
| 1 | 38 | 0.1 | - | F | G | - | P | - | - | M | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 1 | 39 | 0.1 | - | F | - | - | P | - | - | - | V | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | K | - | - | - | - | - | - |
| 1 | 40 | 0.1 | - | F | - | - | P | - | - | - | - | T | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | R | - | - | - | - | - | - |
| 1 | 41 | 0.1 | - | F | - | - | P | - | - | - | - | - | - | - | R | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | R | - | - | - | - | - | - |
| 1 | 42 | 0.1 | - | F | - | - | P | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | R | - | - | I | - | - | - |
| 1 | 43 | 0.1 | - | F | - | - | P | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | K | - | - | - | S | - | - |
| 1 | 44 | 0.1 | - | F | G | - | P | - | - | - | - | - | - | - | - | - | - | - | - | - | - | V | - | - | - | - | - | - | - | - | R | - | - | - | - | - | - |
| 1 | 45 | 0.1 | - | F | G | - | P | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | D | - | - | - | - | - | R | - | - | - | - | - | - |
| 1 | 46 | 0.1 | - | F | G | - | P | - | K | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | K | R | - | - | - | - | - | - |
The sequence of pcrV was determined by whole-genome sequencing followed by translation of amino acid protein sequences. PcrV amino acid sequences of clinical isolates were compared to data from reference strain PAO1 published by Stover et al [3].
Pseudomonas aeruginosa Type 3 Secretion System (PcrV) Amino Acid Variation
| Isolates, No. | Subtype | Percentage | Amino Acid Position, Reference Sequence | ||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 5, N | 6, L | 9, A | 10, R | 21, S | 27, A | 30, E | 33, L | 53, A | 68, S | 69, A | 88, P | 106, H | 112, D | 118, K | 119, D | 123, T | 143, K | 146, S | 154, A | 172, V | 181, V | 182, G | 184, P | 188, D | 199, D | 209, D | 220, E | 225, S | 229, P | 236, P | 237, V | 256, N | 275, A | 285, S | |||
| 431 | 1 | 47.8 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 141 | 2 | 15.6 | - | F | G | - | P | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | R | - | - | - | - | - | - |
| 62 | 3 | 6.9 | - | F | - | - | P | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | K | - | - | - | - | - | - |
| 59 | 4 | 6.5 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | R | - | - | - | - | - | - |
| 50 | 5 | 5.5 | - | F | G | - | P | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | K | - | - | - | - | - | - |
| 27 | 6 | 3.0 | - | - | - | - | - | - | - | - | - | - | - | - | Y | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 19 | 7 | 2.1 | - | F | G | - | P | - | - | M | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | R | - | - | - | - | - | - |
| 15 | 8 | 1.7 | - | F | - | - | P | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | R | - | - | - | - | - | - |
| 12 | 9 | 1.3 | - | F | G | - | P | - | K | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | R | - | - | - | - | - | - |
| 10 | 10 | 1.1 | - | - | G | - | P | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 9 | 11 | 1.0 | - | F | G | - | P | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 9 | 12 | 1.0 | - | F | G | - | P | - | K | - | - | - | - | - | R | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | R | - | - | - | - | - | - |
| 6 | 13 | 0.7 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | G | - | - | - | - | - | - |
| 6 | 14 | 0.7 | - | - | - | - | P | - | - | - | - | - | - | - | - | - | E | A | - | - | G | - | L | - | - | - | G | G | N | - | R | - | - | - | - | - | - |
| 5 | 15 | 0.6 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | I |
| 3 | 16 | 0.3 | - | F | S | - | P | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | R | - | - | - | - | - | - |
| 2 | 17 | 0.2 | - | - | - | - | - | G | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 2 | 18 | 0.2 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | V | I |
| 2 | 19 | 0.2 | - | - | - | - | P | - | - | - | - | - | - | - | - | N | - | - | - | - | - | - | - | - | - | - | - | - | - | - | K | - | - | - | - | - | - |
| 2 | 20 | 0.2 | - | F | S | - | Q | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | R | - | - | - | - | - | - |
| 2 | 21 | 0.2 | - | F | G | - | P | - | K | - | - | - | - | - | Y | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 2 | 22 | 0.2 | - | F | G | - | P | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | S | - | - | - | - | R | - | - | - | - | - | - |
| 2 | 23 | 0.2 | - | - | - | - | P | - | - | - | - | - | - | - | - | - | E | A | - | - | G | - | L | - | - | - | G | - | - | - | R | - | - | - | - | - | - |
| 2 | 24 | 0.2 | - | - | - | - | P | - | - | - | - | - | - | - | - | - | E | A | - | - | G | - | L | - | - | - | G | G | - | - | R | - | - | - | - | - | - |
| 1 | 25 | 0.1 | - | - | V | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 1 | 26 | 0.1 | - | - | - | S | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 1 | 27 | 0.1 | - | - | - | - | - | - | - | - | - | - | P | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 1 | 28 | 0.1 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | R | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 1 | 29 | 0.1 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | S | - | - | - | - | - |
| 1 | 30 | 0.1 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | S | - | - | - | - |
| 1 | 31 | 0.1 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | D | - | - | - | - | - | - | - | - | - | - | - | - |
| 1 | 32 | 0.1 | - | - | - | - | - | - | - | - | - | - | - | L | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 1 | 33 | 0.1 | - | - | - | - | - | - | - | - | - | - | - | - | Y | - | - | - | A | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 1 | 34 | 0.1 | - | F | - | - | P | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 1 | 35 | 0.1 | - | - | G | - | P | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | R | - | - | - | - | - | - |
| 1 | 36 | 0.1 | S | F | - | - | P | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | R | - | - | - | - | - | - |
| 1 | 37 | 0.1 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | L | A | - | - | G | - | - | - | R | - | - | - | - | - | - |
| 1 | 38 | 0.1 | - | F | G | - | P | - | - | M | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 1 | 39 | 0.1 | - | F | - | - | P | - | - | - | V | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | K | - | - | - | - | - | - |
| 1 | 40 | 0.1 | - | F | - | - | P | - | - | - | - | T | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | R | - | - | - | - | - | - |
| 1 | 41 | 0.1 | - | F | - | - | P | - | - | - | - | - | - | - | R | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | R | - | - | - | - | - | - |
| 1 | 42 | 0.1 | - | F | - | - | P | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | R | - | - | I | - | - | - |
| 1 | 43 | 0.1 | - | F | - | - | P | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | K | - | - | - | S | - | - |
| 1 | 44 | 0.1 | - | F | G | - | P | - | - | - | - | - | - | - | - | - | - | - | - | - | - | V | - | - | - | - | - | - | - | - | R | - | - | - | - | - | - |
| 1 | 45 | 0.1 | - | F | G | - | P | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | D | - | - | - | - | - | R | - | - | - | - | - | - |
| 1 | 46 | 0.1 | - | F | G | - | P | - | K | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | K | R | - | - | - | - | - | - |
| Isolates, No. | Subtype | Percentage | Amino Acid Position, Reference Sequence | ||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 5, N | 6, L | 9, A | 10, R | 21, S | 27, A | 30, E | 33, L | 53, A | 68, S | 69, A | 88, P | 106, H | 112, D | 118, K | 119, D | 123, T | 143, K | 146, S | 154, A | 172, V | 181, V | 182, G | 184, P | 188, D | 199, D | 209, D | 220, E | 225, S | 229, P | 236, P | 237, V | 256, N | 275, A | 285, S | |||
| 431 | 1 | 47.8 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 141 | 2 | 15.6 | - | F | G | - | P | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | R | - | - | - | - | - | - |
| 62 | 3 | 6.9 | - | F | - | - | P | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | K | - | - | - | - | - | - |
| 59 | 4 | 6.5 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | R | - | - | - | - | - | - |
| 50 | 5 | 5.5 | - | F | G | - | P | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | K | - | - | - | - | - | - |
| 27 | 6 | 3.0 | - | - | - | - | - | - | - | - | - | - | - | - | Y | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 19 | 7 | 2.1 | - | F | G | - | P | - | - | M | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | R | - | - | - | - | - | - |
| 15 | 8 | 1.7 | - | F | - | - | P | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | R | - | - | - | - | - | - |
| 12 | 9 | 1.3 | - | F | G | - | P | - | K | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | R | - | - | - | - | - | - |
| 10 | 10 | 1.1 | - | - | G | - | P | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 9 | 11 | 1.0 | - | F | G | - | P | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 9 | 12 | 1.0 | - | F | G | - | P | - | K | - | - | - | - | - | R | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | R | - | - | - | - | - | - |
| 6 | 13 | 0.7 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | G | - | - | - | - | - | - |
| 6 | 14 | 0.7 | - | - | - | - | P | - | - | - | - | - | - | - | - | - | E | A | - | - | G | - | L | - | - | - | G | G | N | - | R | - | - | - | - | - | - |
| 5 | 15 | 0.6 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | I |
| 3 | 16 | 0.3 | - | F | S | - | P | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | R | - | - | - | - | - | - |
| 2 | 17 | 0.2 | - | - | - | - | - | G | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 2 | 18 | 0.2 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | V | I |
| 2 | 19 | 0.2 | - | - | - | - | P | - | - | - | - | - | - | - | - | N | - | - | - | - | - | - | - | - | - | - | - | - | - | - | K | - | - | - | - | - | - |
| 2 | 20 | 0.2 | - | F | S | - | Q | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | R | - | - | - | - | - | - |
| 2 | 21 | 0.2 | - | F | G | - | P | - | K | - | - | - | - | - | Y | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 2 | 22 | 0.2 | - | F | G | - | P | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | S | - | - | - | - | R | - | - | - | - | - | - |
| 2 | 23 | 0.2 | - | - | - | - | P | - | - | - | - | - | - | - | - | - | E | A | - | - | G | - | L | - | - | - | G | - | - | - | R | - | - | - | - | - | - |
| 2 | 24 | 0.2 | - | - | - | - | P | - | - | - | - | - | - | - | - | - | E | A | - | - | G | - | L | - | - | - | G | G | - | - | R | - | - | - | - | - | - |
| 1 | 25 | 0.1 | - | - | V | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 1 | 26 | 0.1 | - | - | - | S | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 1 | 27 | 0.1 | - | - | - | - | - | - | - | - | - | - | P | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 1 | 28 | 0.1 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | R | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 1 | 29 | 0.1 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | S | - | - | - | - | - |
| 1 | 30 | 0.1 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | S | - | - | - | - |
| 1 | 31 | 0.1 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | D | - | - | - | - | - | - | - | - | - | - | - | - |
| 1 | 32 | 0.1 | - | - | - | - | - | - | - | - | - | - | - | L | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 1 | 33 | 0.1 | - | - | - | - | - | - | - | - | - | - | - | - | Y | - | - | - | A | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 1 | 34 | 0.1 | - | F | - | - | P | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 1 | 35 | 0.1 | - | - | G | - | P | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | R | - | - | - | - | - | - |
| 1 | 36 | 0.1 | S | F | - | - | P | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | R | - | - | - | - | - | - |
| 1 | 37 | 0.1 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | L | A | - | - | G | - | - | - | R | - | - | - | - | - | - |
| 1 | 38 | 0.1 | - | F | G | - | P | - | - | M | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 1 | 39 | 0.1 | - | F | - | - | P | - | - | - | V | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | K | - | - | - | - | - | - |
| 1 | 40 | 0.1 | - | F | - | - | P | - | - | - | - | T | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | R | - | - | - | - | - | - |
| 1 | 41 | 0.1 | - | F | - | - | P | - | - | - | - | - | - | - | R | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | R | - | - | - | - | - | - |
| 1 | 42 | 0.1 | - | F | - | - | P | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | R | - | - | I | - | - | - |
| 1 | 43 | 0.1 | - | F | - | - | P | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | K | - | - | - | S | - | - |
| 1 | 44 | 0.1 | - | F | G | - | P | - | - | - | - | - | - | - | - | - | - | - | - | - | - | V | - | - | - | - | - | - | - | - | R | - | - | - | - | - | - |
| 1 | 45 | 0.1 | - | F | G | - | P | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | D | - | - | - | - | - | R | - | - | - | - | - | - |
| 1 | 46 | 0.1 | - | F | G | - | P | - | K | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | K | R | - | - | - | - | - | - |
The sequence of pcrV was determined by whole-genome sequencing followed by translation of amino acid protein sequences. PcrV amino acid sequences of clinical isolates were compared to data from reference strain PAO1 published by Stover et al [3].
Binding of PcrV Subtypes by Anti-PcrV MEDI3902 Binding Domain
To confirm that different PcrV subtypes were recognized by the anti-PcrV domain contained within MEDI3902, we evaluated whether the anti-PcrV mAb V2L2MD (CDRs contained within MEDI3902) could bind PcrV expressed from whole-cell lysates of subtype-specific strains cultured under T3S-inducing conditions. Reactivity of lysates to purified polyclonal IgG derived from rabbits immunized with recombinant PcrV was performed to assess whether potential differences in protein reactivity were due to anti-PcrV mAb binding or to in vitro PcrV expression levels. Immunoblot analyses indicated that anti-PcrV mAb binding was comparable to the anti-PcrV polyclonal IgG binding tested in parallel for all 46 PcrV variants (Supplementary Figure 1).
Cocrystal Structure of PcrV Fragment Bound to Anti-PcrV Fab
To determine whether any of the amino acid changes were within the antibody-antigen contact regions between MEDI3902 and PcrV, we solved the crystal structure of a PcrV fragment (PcrVfrag) bound to a Fab of the MEDI3902 anti-PcrV binding domain (V2L2MD-Fab). This structure provided us with precise epitope information and the identification of PcrV-antibody contact residues. The crystals of V2L2MD-Fab and the PcrVFrag complex diffracted to a resolution of 2.8 Å and is depicted in ribbon and space-filling models in Figure 2. PcrVfrag interaction is seen with the V2L2-Fab heavy chain and light chain regions (Figure 2A). Interestingly, calculation of the electrostatic surface potential of V2L2MD-Fab allowed the identification of a highly negatively charged pocket located between the heavy and light chains, which is the primary interaction region with PcrVfrag (Figure 2B). From our model, V2L2-Fab contacts with PcrVfrag residues S157, R159, E227, E231, K232, and D233 (Figure 2C; additional interaction details are available in Supplementary Table 1). Consistent with our anti-PcrV binding data, our crystal structure model confirms that none of variant amino acids were within the V2L2MD-Fab-PcrVfrag binding residues. Variant amino acid sequences near the epitope are depicted in red within the V2L2MD-Fab and PcrVfrag space-filling model offering a 180° view of the interaction, confirming that they are located outside of the MEDI3902 anti-PcrV epitope (Figure 2D). These data strongly suggest that the PcrV epitope targeted by MEDI3902 is likely an essential and therefore highly conserved domain of PcrV.
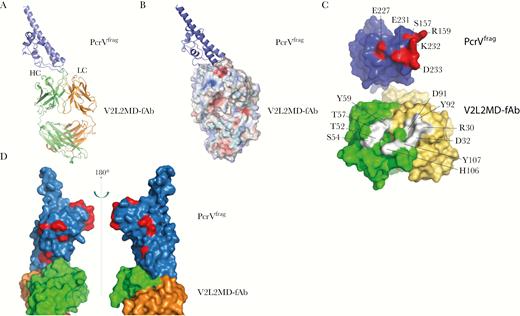
Identification of the anti-PcrV binding epitope of MEDI3902. Cocrystals of V2L2MD-Fab and the PcrVFrag diffracted to a resolution of 2.8 Å. A, Ribbon model of V2L2MD-Fab complexed to PcrVFrag. The light chain (LC) is gold, the heavy chain (HC) is green, and the PcrVFrag is blue. B, Space-filling model V2L2MD-Fab complexed to PcrVFrag. A highly negatively charged pocket interacting with PcrVFrag between CDR1 and 3 of the LC and CDR3 of the HC is indicated in red. C, Interacting surfaces of V2L2-Fab and PcrVFrag are shown in open-book form. Amino acids participating in hydrogen bonding and their numbers are shown. PcrVFrag amino acids are shown in red. V2L2-Fab amino acids are shown in white. D, 180° rotation around the vertical axes of the V2L2-Fab and PcrVFrag complex. Of 35 unique amino acid substitutions, 15 were present in PcrVFrag used for structural analysis and are in red. None of the amino acid changes were within V2L2-Fab and PcrVFrag interaction points.
Identification of T3S-Injectisome and Exoenzyme-Negative Isolates
Of the 913 P. aeruginosa geographically diverse clinical isolates, only 11 strains were pcrV negative. While these strains are rare, they were collected in geographically diverse locations from 2008 to 2014 (Supplementary Table 2). Further investigation into the genotype of these isolates revealed that they lack the entire T3S-injectisome operon and the associated T3S exoenzymes but contain the recently described type-2-secretion system-Txc operon (T2SS-Txc), consistent with these isolates being members of a PA7-like clade [35–37]. These strains are referred to as clonal outliers, based on their sequence and genome divergence from P. aeruginosa clinical isolates [28, 37–39]. Whole-genome–based phylogenetic analysis indicates that these isolates are clustered into a single identifiable outgroup (Figure 3A). When evaluated for cytotoxic activity against cultured epithelial cells, 4 of 11 isolates (strains 808, 566, 319, and 15042) exhibited the ability to lyse epithelial cells, similar to the previously reported PA7 isolates, albeit at a much higher multiplicity of infection when compared to a T3S-injectisome ExoU+ (phospholipase) P. aeruginosa strain (Figure 3B). PAO1, which expresses ExoS (ADP ribosyltransferase plus Rho GTPase activity) rather than ExoU, exhibits little to no cytotoxicity in this assay and was used as a negative control (Figure 3B). In contrast to its potent anticytotoxic activity against T3S-injectisome ExoU+ strains, MEDI3902 was unable to prevent lysis of an epithelial cell line in cytotoxicity assays with PA7-like strains (Figure 3C). However, 10 of 11 PA7-like isolates contained all of the genes required for Psl expression in vitro (Figure 1). Given the high prevalence of Psl expression in PA7-like strains, we postulated that these isolates would be susceptible to anti-Psl–mediated OPK activity. Indeed, both the anti-Psl mAb alone and MEDI3902 mediated potent OPK activity against a representative PA7-like isolate (Figure 4A).
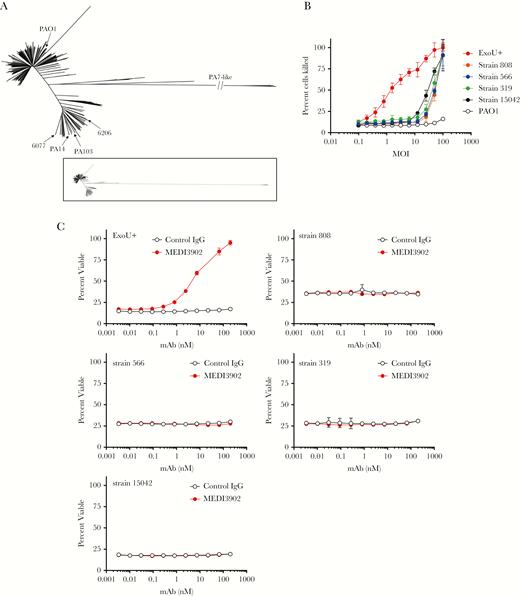
Identification of T3S-injectisome and exoenzyme negative (PA7-like) isolates. A, Core genome–based tree of 913 Pseudomonas aeruginosa genomes with reference strains PAO1, PA14, PA103, 6206, and 6077. The inset illustrates the distance between PA7-like and T3S-injectisome–positive isolates. B, Cytotoxicity assay comparing PA7-like isolates to T3S-injectisome and ExoU+ strain 6077. C, Cytotoxicity assays evaluating MEDI3902 activity against PA7-like isolates and strain 6077. IgG, immunoglobulin G; mAb, monoclonal antibody; MOI, multiplicity of infection.
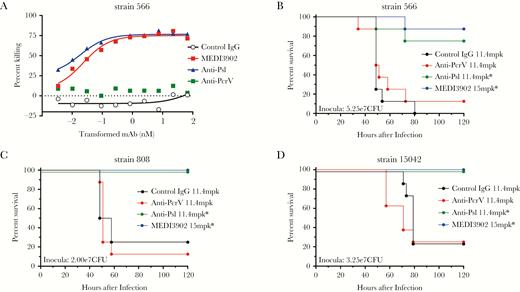
Anti-Psl activity mediates potent protective activity against T3S-negative PA7-like isolates. A, Opsonophagocytic killing activity assay. B–D, Kaplan-Meier curves of acute pneumonia models depicting the protective activity of anti-Psl and MEDI3902 against T3S-negative PA7-like isolates. n = 8 for each monoclonal antibody (mAb) treatment group. CFU, colony-forming units; IgG, immunoglobulin G. *P < .05, by the log-rank test.
Although PA7-like strains are deficient for the T3S injectisome, they contain a novel T2SS-Txc operon and are thought to be capable of inducing lethal infection in mice via secretion of a pore-forming toxin, exolysin [36, 39]. We evaluated representative cytotoxic PA7-like strains in the acute pneumonia model and confirmed their ability to induce lethal pneumonia (Supplementary Figure 2). As depicted in Figure 4B–D, both anti-Psl mAb Psl0096 (CDRs contained within MEDI3902) and MEDI3902 conferred potent protective activity against PA7-like strains. As expected, no protection was observed in mice treated with control IgG or the anti-PcrV mAb (Figure 4B–D). We also identified 12 strains that were T3S-injectisome positive and contained portions of the T2SS-Txc operon. MEDI3902 prevented lethal infection against all T3S-injectisome– and T2SS-Txc–positive isolates tested in the acute pneumonia model (Supplementary Figure 3). A summary of our Psl and PcrV surveillance results are depicted in Figure 5, wherein a whole-genome–based tree is decorated with year and geographical region of collection, along with pcrV gene and psl operon presence/absence. Altogether, our results indicate that MEDI3902 can provide broad coverage against P. aeruginosa strains, even when one of its targets is not expressed by the clinical isolate.
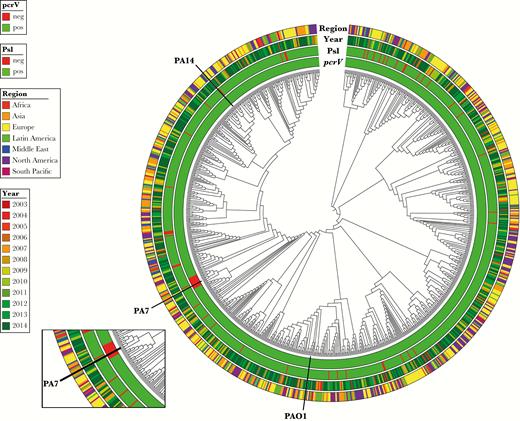
Summary of psl operon and pcrV surveillance data from 913 Pseudomonas aeruginosa clinical isolates. Core genome–based tree with ignored branch lengths is decorated with year and geographical region of collection, along with psl operon and pcrV gene presence (green) or absence (red). Isolates are colored according to their region and year of collections. Reference genomes PAO1, PA7, and PA14 are marked.
DISCUSSION
This study surveyed 913 hospital P. aeruginosa respiratory isolates collected from a diverse patient population in 45 countries from 2003 to 2014 to assess the presence and conservation of PcrV and Psl, the molecular targets of bispecific monoclonal antibody MEDI3902. To our knowledge, this is the first large international surveillance study that characterized the prevalence and variability of these therapeutic targets. Whole genome sequencing analysis revealed that 99% of isolates (902) from the surveillance collection possessed a full-length pcrV gene. An intact psl operon, which is composed of 15 genes (pslA through pslO), was present in 94% of isolates (859). These results emphasize the importance of both PcrV component of T3S system and Psl exopolysaccharide for P. aeruginosa virulence and persistence in the host environment and, at the same time, demonstrate that molecular targets of MEDI3902 are highly prevalent and conserved.
We discovered that 3% of isolates (28) lacked an entire Psl operon and that another 3% (26) lacked ≥1 Psl operon gene. Previous work by Byrd et al characterized the in-frame deletions in each of the 15 psl genes and demonstrated that 4, pslB, pslM, pslN, and pslO, are not essential for Psl synthesis [16]. Consistent with these results, the pslM-null and pslNO-null isolates in our collection expressed Psl, whereas the isolates that lacked one of the pslA, pslC, pslHI, or pslE-O genes did not express detectable Psl. A single Psl-negative isolate contained a deletion in the pslA-pslB intergenic region. The naturally occurring Psl-negative isolates from our collection may point to the existence of selective pressures driving evolution of the psl operon toward adaptation to certain specific environmental factors. We selected a set of Psl-positive isolates, including strains with the complete psl operon and strains lacking nonessential psl genes, and confirmed MEDI3902 binding for the entire set. All but one of the Psl-negative isolates from our collection expressed PcrV and, therefore, retained the ability to interact with the anti-PcrV arm of MEDI3902.
Based on the analysis of PcrV sequences from the study isolates, we identified 46 distinct PcrV subtypes containing amino acid substitutions at 35 unique positions as compared to the reference PcrV sequence from PAO1. These variants shared at least 96.3% identity to the reference sequence, and representative strains from each PcrV variant sequence were bound by the anti-PcrV specificity contained in MEDI3902, including the most divergent sequence substitutions. The crystal structure analysis of an anti-PcrV Fab-PcrV fragment complex indicated that none of the mutations in PcrV sequences were within the antibody epitope (Figure 2).
There was a small number of pcrV gene–negative isolates (n = 11) identified in this study lacking the entire T3S-injectisome operon and T3S-related exotoxins. Interestingly, although our genotypic analysis suggested that these isolates belong to a single lineage, they vary significantly by temporal and geographic locations (Figure 3A). These observations are consistent with the identification of PA7-like isolates, which were reported as negative for both T3S-injectisome operon and associated exotoxin genes [35, 37]. In agreement with the work presented by Cadoret et al, we detected the presence of the novel T2SS-Txc operon in these isolates, thus further confirming the identification of a set of PA7-like isolates in our surveillance collection [36]. Interestingly, all but 2 of the PA7-like isolates lacked the pslM gene that is dispensable for Psl synthesis (Byrd et al [16] and the present study). In vitro cytotoxicity testing revealed cytotoxic activity at a high multiplicity of infection for some of the representative PA7-like isolates in comparison to the ExoU+ strain 6077 (Figure 3B and 3C). Further studies are required to determine whether the cytotoxicity of these isolates is mediated via the T2SS-Txc. As expected, MEDI3902 did not block the cytotoxic effect of these T3S-injectisome null isolates, but both anti-Psl mAb and MEDI3902 were capable of mediating potent opsonophagocytic killing of a representative PA7-like isolate. We also demonstrated the lethal activity of T2SS-Txc–positive cytotoxic strains in a mouse pneumonia model, albeit at a greater inoculum as compared to ExoU+ strains. In this infection model, both MEDI3902 and the anti-Psl mAb alone but not an anti-PcrV mAb were protective, in accordance with these isolates lacking the PcrV target protein yet retaining the ability to express Psl. Consistent with our previous results [24, 32], these data confirm the protective activity of MEDI3902, even in the absence of one of its targets.
Our work does have some limitations. For example, patient outcomes were not available; therefore, we were unable to correlate clinical outcomes with any of the above-mentioned P. aeruginosa strain types on the basis of the presence or absence of pcrV and psl genes and their variations. In addition, another practical limitation is that Psl and PcrV expression was assessed via in vitro assays, and therefore the detectable values may not represent the levels expressed during in vivo infection. Despite these limitations, our study showed that 99.9% of global P. aeruginosa respiratory isolates contained at least one of either the psl operon or pcrV genetic elements and demonstrated potent MEDI3902 activity against diverse strain types, including strains that expressed only a single target and strains with target sequence variations. Multidrug-resistant isolates comprised a large proportion of our collection, but nevertheless MEDI3902 would provide coverage against these multidrug-resistant isolates owing to the high prevalence and conservation of PcrV and Psl. Thus, our results indicate that MEDI3902 could provide broad strain coverage and activity against P. aeruginosa respiratory strains from diverse locations around the world, including strains that are resistant to multiple antibiotics.
Notes
Acknowledgments. We thank JoAnn Suzich, for her critical review of the manuscript; the laboratory personnel at International Health Management Associates, for maintenance of the P. aeruginosa repository; and all members of the global surveillance program who contributed isolates and information for this study.
D. E. T., V. O., A. E. K., R. E. M., E. S., P. W., K. R., and A. D. contributed experimental data; Y. L. and Y. Q. performed statistical analyses; and D. E. T., A. R., C. K. S., and A. D. wrote the manuscript.
Financial support. This work was supported by MedImmune/AstraZeneca.
Potential conflicts of interest. All authors are employees of MedImmune/AstraZeneca and may hold stock in the company. All authors have submitted the ICMJE Form for Disclosure of Potential Conflicts of Interest. Conflicts that the editors consider relevant to the content of the manuscript have been disclosed.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}