





Nutrient sensing in the hypothalamus is tightly related to food intake regulation. However, the mechanisms by which the nutrient-sensing cells of the brain translate this signal of energy need into feeding behavior via regulation of neuropeptide expression are not known. To address this issue, we investigated two neuronal cell lines expressing agouti-related protein (AgRP), ex vivo hypothalamic tissues, and in vivo whole animals. Maintaining cells in a low cellular ATP concentration generated by low glucose, 2-deoxyglucose (2-DG), ATP synthesis inhibitor, and 5-aminoimidazole-4-carboxamide 1-β-d-ribofuranoside increased phosphorylation of AMP-activated protein kinase (AMPK) and increased AgRP expression, whereas maintaining cells in high ATP status by high glucose and pyruvate supplementation in 2-DG-treated cells decreased phosphorylation of AMPK and decreased AgRP expression. Overexpression of a dominant-inhibitory mutant of AMPK significantly decreased low-glucose- or 2-DG-induced AgRP expression. Furthermore, ex vivo hypothalamus culture in high glucose concentrations decreased both expression and phosphorylation of AMPK and expression of both AgRP and neuropeptide Y, whereas pyruvate supplementation suppressed a 2-DG-induced AgRP expression. Finally, our in vivo studies clearly show that central administration of pyruvate dramatically delayed 2-DG-induced food intake. These data indicate that modulation of ATP levels in neuronal cells triggers a cascade of events via AMPK that modulate feeding behavior to restore energy status of cells.
A LARGE BODY of evidence supports the roles of the central nervous system in regulating feeding behavior and body fat content (1, 2). The brain receives signals of whole-body energy status via hormones and metabolites such as leptin, ghrelin, insulin, glucagon-like peptide 1, glucose, amino acids, and fatty acids. It is proposed that these diverse and complex signals are integrated by neurons in the arcuate nucleus, including neuropeptide Y (NPY)/agouti-related protein (AgRP) neurons and proopiomelanocortin (POMC)/cocaine-amphetamine-related transcript neurons.
Growing experimental evidence supports the classical glucostatic (3, 4) and ischymetric hypotheses (5). Specifically, there is increasing evidence for existence of cells in the arcuate nucleus and hindbrain that have the correct gene expression (6–8), electrophysiology (9), and feeding behavioral response to nutrients that supply energy to the cell. For example, central administration of 2-deoxy-d-glucose (2-DG), a nonmetabolizable glucose analog that inhibits glucose use, increases AgRP and NPY gene expression and elicits glucoprivic food intake (10, 11). However, the mechanisms by which signals from metabolites are translated into neurochemical signals still remain unsolved.
To address this issue, we developed in vitro and ex vivo systems for cellular level studies. In the N1E-115 and GT1-7 neuroblastoma cell lines, glucose suppressed expression of AgRP, whereas 2-DG induced AgRP expression. Next, we provided evidence that the modulation of cellular ATP concentration by glucose, 2-DG, pyruvate, ATP synthesis inhibitor, and 5-aminoimidazole-4-carboxamide 1-β-d-ribofuranoside (AICAR) regulates AgRP expression, probably through the AMP-activated protein kinase (AMPK) pathway. Our findings in vitro were further confirmed by ex vivo and in vivo studies where changes in neuronal energy status affect AMPK phosphorylation and neuropeptide expression, leading to changes in food intake. Our present data suggest that energy sensing through ATP status is a main switch to regulate neuropeptide expression and food intake regulation.
Materials and Methods
Cell culture and treatment
N1E-115 neuroblastoma was obtained from ATCC (Manassas, VA), and GT1-7 cell line was a generous gift from Dr. Pamela Mellon (University of California, San Diego, San Diego, CA). The cells were cultured in DMEM containing 25 mm glucose, 10% fetal bovine serum, 50 U of penicillin, and 50 μg/ml streptomycin at 37 C in 5% CO2. For analysis of the effects of metabolites on AgRP expression, the confluent cells were further cultured in DMEM containing a low percentage (1%) of serum because serum contains glucose and factors (for example insulin and leptin) that can affect AMPK phosphorylation and neuropeptide expression. To test glucose or 2-DG levels on AgRP expression, cells were cultured in various levels (0.5, 1, 2.5, 5, 10, and 25 mm) of glucose or 2.5 mm glucose plus various concentrations of 2-DG (0, 1, 2.5, 5, 10, and 25 mm) for 16 h. All of the experiments with cell lines were done using media containing 1% fetal bovine serum (FBS) and 2.5 mm glucose.
Plasmids and transfection
pcDNA3-Myc-tagged wild-type and dominant-inhibitory mouse AMPKα (AMPKα2WT and AMPKα2KD) (12) were subcloned into pTRE-puro plasmid (Invitrogen Corp., Carlsbad, CA). GT1-7 neuroblastoma cells (∼70% confluent) were transfected with pTRE-puro-AMPKα2WT or pTRE-puro-AMPKα2KD plasmids using the LipofectAMINE reagent as described by the manufacturer’s protocol (Invitrogen). Transfected cells were grown 5 d in DMEM containing 1 μg/ml puromycin, and the pool of puromycin-resistant clones was amplified for subsequent treatments. Transfected cells were incubated with medium containing 1% FBS plus 1, 2.5, or 10 mm glucose, or 2.5 mm glucose plus 10 mm 2-DG for 16 h.
Ex vivo experiments
Fresh hypothalamuses with the dimension of 3.2 mm × 3.5 mm × 2 mm and the average weight of approximately 7 mg were excised from mice and immediately cultured in the media containing DMEM, 1% FBS, and various concentrations of glucose (1, 2.5, 5, and 10 mm) for 2 h. The hypothalamuses were also incubated with media containing DMEM in 2.5 mm glucose with either 10 mm 2-DG or 2-DG (10 mm) plus pyruvate (10 mm) for 2 h. The protein was extracted from the hypothalamus and subjected to Western blot for phospho-AMPK, AMPK, and β-actin. The total RNA was also extracted from the hypothalamuses and subjected to quantitative real-time RT-PCR for AgRP, NPY, POMC, and cyclophilin. The results were reported as percentages of values obtained from expression of target genes divided by cyclophilin expression with six samples per group.
Quantitative real-time RT-PCR
Real time RT-PCR was performed in a 25-μl final reaction volume using TaqMan 1000 Rxn PCR core reagent kit (Applied Biosystems, Branchburg, NJ). Cyclophilin mRNA levels from each sample were used as internal controls to normalize the mRNA levels (13). Detailed methods and PCR condition, using ABI PRISM 7700 sequence detector (Applied Biosystems), are described in our previous report (6). The data within the linear region of the amplification curve are analyzed according to ABI’s user bulletin No. 2. The TaqMan probe is dual labeled with 5′-FAM and 3′-BHQ. Primer and probe sequences are available on request.
ATP concentration
Total cellular ATP concentration was measured using the bioluminescent somatic cell assay kit (Sigma Chemical Co., St. Louis, MO) according to the manufacturer’s instructions.
Western blot
The cells and tissues were homogenized in ice-cold lysis buffer as described in a previous report (14). The membranes were incubated with polyclonal primary antibodies, phospho-AMPK antibody (Cell Signaling Technology, Inc., Beverly, MA; 1:300 dilution), and AMPK antibody (Cell Signaling; 1:1000 dilution) in blocking solution for 1 h at room temperature and washed five times for 10 min each with TBS plus 0.05% Tween 20. The membranes were further incubated with horseradish peroxidase-conjugated secondary antibody for 1 h at room temperature.
Experimental animals, cannulation, and intracerebroventricular (ICV) injection
Male Sprague Dawley rats, 230–270 g, were maintained on a 12-h light, 12-h dark cycle with lights on at 0700 h. All rats were singly housed and fed ad libitum. Drinking water was available at all times. Animal experiments were approved by the Pennington Biomedical Research Center Animal Care and Use Committee. For cannulation, rats were anesthetized with Ketamine/acepromazine/xylazine, and 24-gauge guide cannulas (Plastics One, Roanoke, VA) were placed in the lateral cerebroventricle (0.8 mm posterior to bregma, 1.4 mm lateral to midline, and 3.5 mm below the skull). After a 10-d recovery, rats were injected with 10 μl of saline, 2-DG (3 mg), or 2-DG (3 mg) plus pyruvate (4 mg) via the cannula during 0900–1030 h. Food intake was recorded at 30 min, 1 h, and 2 h after injection.
Data presentation
Data from in vitro studies are presented as mean ± sem of at least three independent experiments performed in duplicate for in vitro experiments. Comparisons involving more than two groups were analyzed using ANOVA. Student’s t test was used when determining the significant difference between two groups. The minimum level of significance was set at P < 0.05.
Results
We first selected two independent neuronal cell lines, N1E-115 and GT1-7 neuroblastoma cell lines expressing AgRP (15, 16), to examine whether changes in energy status, such as glucose deprivation or high levels of glucose, can affect expression of AgRP, a potent hunger neuropeptide. As shown in Fig. 1A, increasing glucose levels produced a dose-dependent decrease in AgRP expression in both cell lines. Because glucose is a major energy source, we next tested whether AgRP expression is related to changes in cellular energy availability. Around 2.5–5 mm glucose, cellular ATP concentrations were sharply increased up to 2-fold, compared with 1 mm glucose. Above 5 mm glucose, ATP levels were not further increased (Fig. 1B).
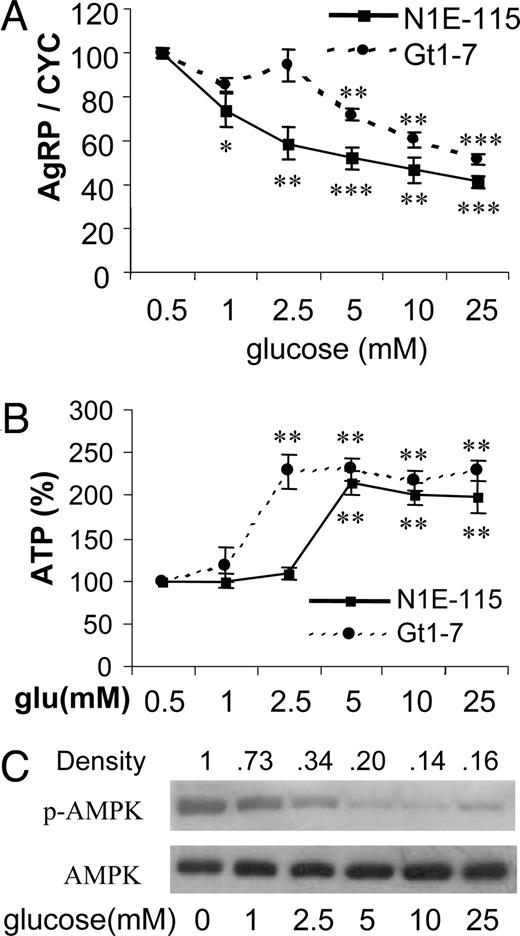
Glucose suppresses expression of AgRP expression in neuronal cell lines. A, Two cell lines were cultured at various concentrations of glucose for 16 h. Total RNAs were subjected to quantitative real-time RT-PCR for AgRP gene expression and normalization with cyclophilin (CYC) gene expression. Data are presented as percentile mean ± sem value of five independent experiments. B, Relative ATP concentrations were determined using the luciferase bioassay as described in Materials and Methods. Data are presented as percentage changes of mean ± sem values of four independent experiments. Statistical differences vs. 0.5 mm (glucose): *, P < 0.05; **, P < 0.01; ***, P < 0.001. C, Cell extracts were prepared and subjected to Western blot using anti-phospho-AMPK (p-AMPK) and anti-AMPK.
AMPK functions as a fuel sensor in many tissues and organs where it inhibits anabolic pathways when cellular ATP levels are depleted and when AMP rises in response to limited energy availability (17). AMPK has been known to be expressed in the brain and to be activated in response to glucose deprivation (18). In our study, it was of interest to investigate whether various energy statuses at different glucose concentrations could modulate the phosphorylation state of AMPK. Our Western blot, using an antibody specific for phosphorylated threonine at the 172nd amino acid of AMPK, showed that glucose lower than that of physiological levels increased AMPK phosphorylation, whereas elevation of glucose concentration over the physiological range dramatically decreased phosphorylation of AMPK (Fig. 1C).
2-DG has been known as an inhibitor of glucose use and a low-glucose mimetic in a wide variety of physiological situations (19). We further investigated whether 2-DG could reverse the down-regulation of AgRP expression as previously shown with high levels of glucose in Fig. 1A. In both cell lines, 2-DG linearly increased expression of AgRP in a dose-dependent manner (Fig. 2A). Because 2-DG inhibits glycolysis, we tested whether 2-DG-induced AgRP expression is related to changes in cellular energy availability. Treatment of neuronal cell cultures with various 2-DG concentrations showed a significant dose-dependent decrease in cellular ATP levels in both cell lines (Fig. 2B). In addition, the reduction of cellular ATP concentration by 2-DG treatment increased phosphorylation of AMPK in a dose-dependent manner (Fig. 2C). Taken together, a decrease in cellular ATP levels by either low glucose or increased 2-DG increased AMPK phosphorylation, which is presumably associated with increased AgRP expression in these low-energy statuses.

2-DG increases expression of AgRP expression in neuronal cell lines. A, Two cell lines were cultured at various concentrations of 2-deoxyglucose for 16 h. Total RNAs were subjected to quantitative real-time RT-PCR for AgRP gene expression. Data are presented as percentile mean ± sem value of four independent experiments. Statistical differences vs. 0 mm (2-DG): * and #, P < 0.05; **, P < 0.01; ***, P < 0.001. B, Relative ATP concentrations were determined using the luciferase bioassay as described in Materials and Methods. C, Cell extracts were prepared and subjected to Western blot using anti-phospho-AMPK (p-AMPK) and anti-AMPK.
Because 2-DG blocks glycolysis and also inhibits mitochondrial respiration caused by depletion of pyruvate, we tested whether pyruvate can bypass the blockage of glycolysis by 2-DG treatment and inhibit 2-DG-induced AgRP expression. As shown in Fig. 3A, supplementation of pyruvate resulted in a reduction of AgRP expression compared with the basal condition. Furthermore, pyruvate attenuated or reversed a 2-DG-induced increase in AgRP expression in both cell lines. We then studied whether pyruvate could increase cellular ATP concentration in 2-DG-treated cells, in which ATP concentration might be a downstream effector molecule to change AgRP expression. Cellular levels of ATP were measured in these cells treated with 2-DG and pyruvate. Incubation of N1E-115 cell lines with 5 mm pyruvate significantly increased cellular ATP levels by 18%, whereas 2-DG decreased ATP levels by 12% (Fig. 3B). In addition, pyruvate prevented the decrease in cellular ATP levels in 2-DG-treated N1E-115 cells. These results indicate that supplementation of pyruvate rescues the low-energy status of 2-DG-treated cells, which is associated with induction of AgRP expression.
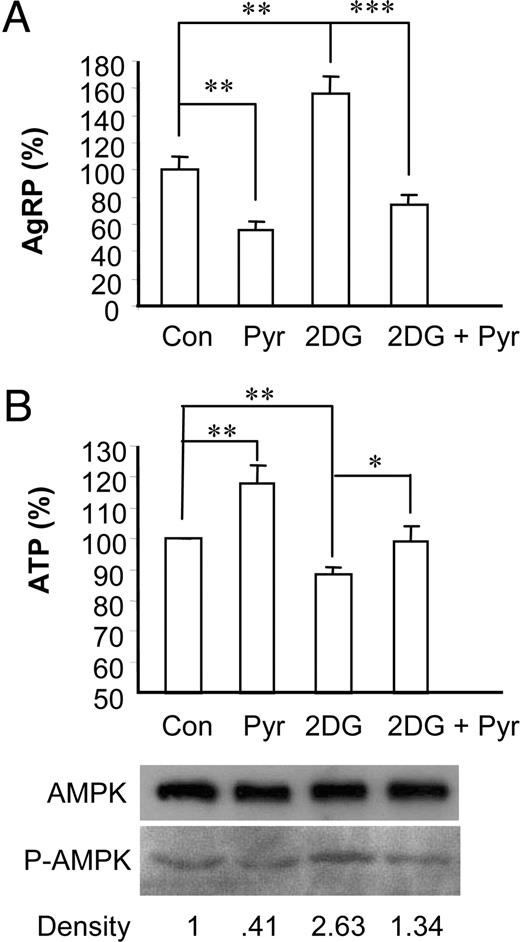
Pyruvate suppresses 2-DG-induced AgRP expression in N1E-115 neuronal cell line. Cells were cultured in media containing 2.5 mm glucose (control) plus pyruvate (5 mm), 2-DG (10 mm), or 2-DG (10 mm) plus pyruvate (5 mm) for 16 h. A, Total RNA was extracted for quantitative real-time RT-PCR to measure AgRP expression. B, Relative ATP concentrations were measured. Data are presented as mean ± sem value of three to five independent experiments. *, P < 0.05; **, P < 0.01. Cell extracts were prepared and subjected to Western blot using anti-phospho-AMPK (P-AMPK) and anti-AMPK.
The decreased ATP content of the neuronal cell lines after 2-DG treatment and its rescue by pyruvate suggests that cellular ATP concentration may be negatively correlated with the expression patterns of AgRP expression as shown in Fig. 3. Therefore, we next asked whether direct modulation of intracellular ATP concentration could affect AgRP expression. To study the effect of ATP depletion on AgRP expression, we used sodium azide (NaN3), which affects mitochondrial oxidative phosphorylation. As shown in Fig. 4B, NaN3 decreased ATP concentration and increased the amount of phosphorylated AMPK in a dose-dependent manner. Furthermore, depletion of ATP by NaN3 treatment increased expression of AgRP in a dose-dependent manner (Fig. 4A). These data clearly show that changes in ATP concentration directly regulate AgRP expression, suggesting a possibility that ATP concentration can regulate activation of AMPK that serves as a downstream effector molecule in the signaling pathway of glucose- or 2-DG-regulated AgRP expression.
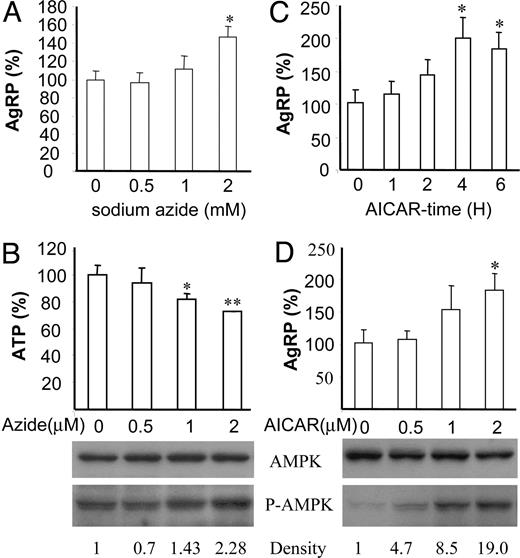
Activators of AMPK, sodium azide, and AICAR induce expression of AgRP in neuronal cell lines. A, N1E-115 neuronal cells were treated with indicated dosage of sodium azide for 4 h, and total RNA extracted from those cells was used for quantitative real-time RT-PCR for AgRP expression. A–D, Data are presented as percentile mean ± sem value of three independent experiments. Statistical differences vs. control (no treatment): *, P < 0.05; **, P < 0.01. B, Relative ATP concentrations in the cells treated with various dosages of sodium azide were measured. Cell extracts were prepared and subjected to Western blot using anti-phospho-AMPK (P-AMPK), and anti-AMPK. C, Time-course study of AICAR (2 mm) on AgRP expression. Three independent experiments were performed in duplicate. D, Dose-dependent increases in AgRP expression in response to AICAR. N1E-115 cells were treated with indicated dosages of AICAR for 4 h, and protein was extracted from those cells and subjected to Western blot for phosphorylated AMPK (P-AMPK) and total AMPK.
Because low cellular ATP levels are associated with increased AgRP expression, the most probable mechanism is that the decreased levels of ATP together with the concomitant increase in AMP levels become a signal that is responsible for the induction of AgRP expression. To test this possibility, N1E-115 cells were treated with AICAR, a cell-permeable AMP analog. As shown in Fig. 4C, 2 mm AICAR could increase AgRP expression linearly up to 2-fold at 4 h in a time-course study. AICAR induced phosphorylation of AMPK and AgRP expression in a dose-dependent manner (Fig. 4D). These results show that direct modulation of either ATP or AMP concentrations could affect AgRP expression, indicating again that cellular energy status can be an important signal for regulation of AgRP expression.
The experiments presented so far suggested that changes in cellular energy status regulate AgRP expression possibly via AMPK. Next, we show direct evidence that AMPK is involved in regulation of AgRP expression. We overexpressed wild-type and dominant-negative AMPKα2 and tested whether expression of the AMPKα2-DN construct could suppress induction of AgRP expression in response to low glucose and 2-DG. Low glucose induced expression of AgRP in cells expressing AMPKα2WT, whereas AMPKα2-DN expression blocked low-glucose-induced AgRP expression (Fig. 5B). In addition, AMPKα2-DN overexpression attenuated 2-DG-induced AgRP expression. These results clearly show the contribution of AMPK in the regulation of AgRP expression.
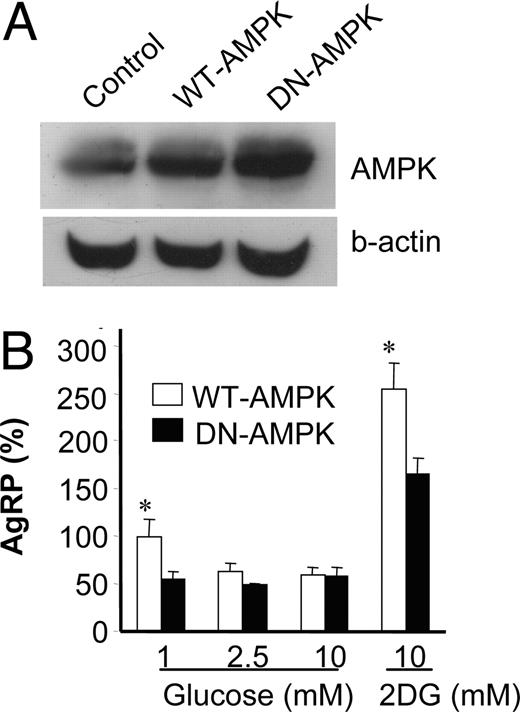
Overexpression of AMPK attenuated low-glucose- or 2-DG-induced AgRP expression. A, The cell lysates from stably transfected GT1-7 cells with vectors containing either AMPKα2-WT or AMPKα2-DN were subjected to Western blot for total AMPK and β-actin (b-actin). B, Stably transfected cells were incubated with medium containing 1, 2.5, or 10 mm glucose or 2-DG (10 mm) overnight. Then, the AgRP expression in these cells was measured by quantitative real-time PCR. Results are expressed as percentile mean ± sem of four experiments. Statistical differences between WT-AMPK vs. DN-AMPK: *, P < 0.05.
Our in vitro model allows more direct testing of the effect of glucose on AgRP expression. However, this system cannot address the indirect effects of interactions between cells within the hypothalamus because astrocytes and glial cells have been known to provide nutrients to the neighboring neuronal cells (20). Therefore, we further investigated whether changes in cellular energy status could affect neuropeptide gene expression ex vivo. The fresh hypothalamuses were cultured ex vivo in 1, 2.5, 5, or 10 mm glucose concentrations for 2 h. AgRP expression was sharply decreased from 1–2.5 mm glucose but not further decreased at 5 and 10 mm glucose concentrations (Fig. 6A), which indicates saturation of the responsiveness above a 2.5-mm glucose concentration. NPY expression was decreased with increased glucose concentration in a dose-dependent manner (Fig. 6B.), which agrees well with the down-regulation of NPY gene expression in the hypothalamus by high-energy statuses, such as feeding or ICV glucose. Expression of POMC was not significantly different but had a tendency to be up-regulated with increased glucose concentrations (Fig. 6C). These studies, using ex vivo hypothalamic tissue culture, are the first reports to show that modulation of nutrients can regulate neuropeptide expression ex vivo. In addition, Western blot for phospho-AMPK and total AMPK showed that, compared with 10 mm glucose, hypothalamus cultured in 1 mm glucose increased AMPK phosphorylation, suggesting that low glucose activated AMPK more highly (Fig. 6D). Furthermore, the AMPK protein levels were elevated in the hypothalamus treated with low glucose.
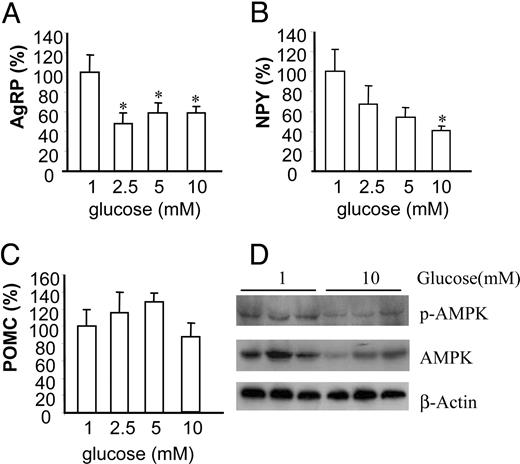
Glucose regulates neuropeptide expression in hypothalamus in ex vivo culture. A–C, Fresh hypothalamuses were cultured in various concentrations of glucose for 2 h. Total RNA was extracted from hypothalamuses for quantitative real-time RT-PCR for AgRP (A), NPY (B), and POMC (C). Data are presented as percentile mean ± sem value of six mouse hypothalamuses. Statistical differences vs. 1 mm (glucose): *, P < 0.05. D, Hypothalamuses were cultured ex vivo at 1 or 10 mm glucose concentrations for 2 h, and total protein was extracted from hypothalamuses and subjected to Western blot for phosphorylated AMPK (p-AMPK) and total AMPK. β-Actin serves as a validation for loading equal amounts of samples.
We next confirmed our in vitro finding that pyruvate supplementation suppressed 2-DG-induced AgRP expression in the ex vivo condition. Ex vivo hypothalamus cultured with 2-DG significantly increased AgRP expression, and supplementation of pyruvate suppressed 2-DG-induced AgRP expression (Fig 7A). To relate to food intake, we then tested whether pyruvate coadministration with 2-DG would attenuate 2-DG-induced food intake. Consistent with our previous reports and others (21, 22), in these studies, ICV 2-DG robustly stimulated food intake with the greatest intake (2.73 ± 0.19 g) occurring within the first 30 min after 2-DG administration, whereas the saline-injected rats ate 0.88 ± 0.23 g within the first hour. Furthermore, pyruvate supplementation dramatically inhibited 2-DG-induced food intake for the first 30 min (Fig. 7B). For the next 30 min, suppression of 2-DG-induced food intake by pyruvate was not observed, and food intake was increased dramatically, suggesting pyruvate supplementation delays 2-DG-stimulated feeding.
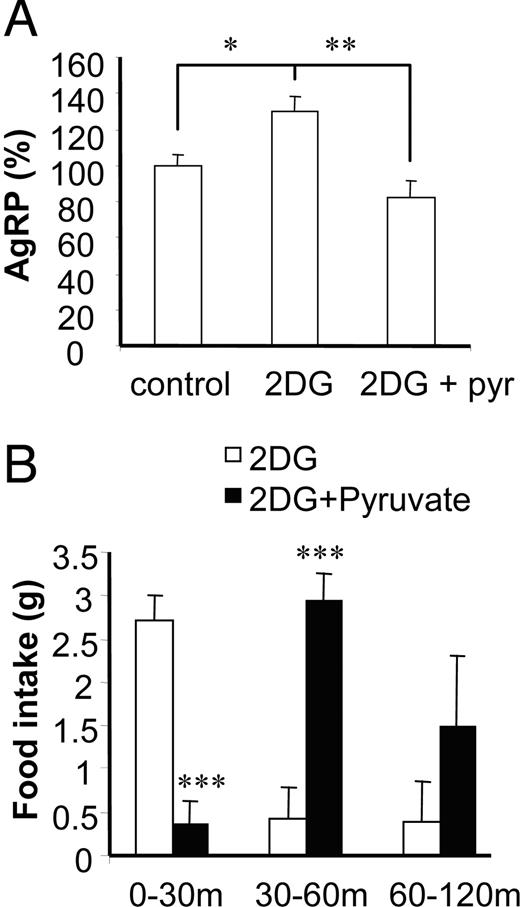
Pyruvate supplementation suppressed 2-DG-induced AgRP expression, and central administration of pyruvate delays 2-DG-induced hyperphagia. A, Hypothalamuses were cultured ex vivo in media containing 2.5 mm glucose, 2.5 mm glucose plus 2-DG (10 mm), or 2.5 mm glucose plus 2-DG plus pyruvate (pyr) (10 mm) for 2 h. Data are presented as percentile mean ± sem value of at least six mouse hypothalamuses. *, P < 0.05; **, P < 0.01. B, 2-DG (3 mg) or 2-DG (3 mg) plus pyruvate (4 mg) in a 10-μl volume were injected into the lateral ventricle of rats. Food intake was measured at indicated time intervals. Data are presented as mean ± sem value of four rats per group. Statistical differences between 2-DG vs. 2-DG plus pyruvate: ***, P < 0.001.

Schematic diagram. Glucose down-regulates AgRP expression through AMPK. 2-DG inhibits glycolysis, decreases cellular ATP levels, increases phosphorylation of AMPK, and increases AgRP expression. Pyruvate bypasses glycolysis and suppresses 2-DG-induced AgRP expression. Direct modulation AMPK by ATP synthesis inhibitor and AICAR (a cell-permeable adenosine analog) regulates AgRP expression. TCA, tricarboxylic acid cycle.
Discussion
It is clear that neuronal cells sense nutrient availability and signal relevant neuronal pathways that lead to feeding behavior. Until now, it was not known how specific neuronal cells convert nutrient status into feeding signals. To understand the cellular and molecular mechanisms by which nutrients regulate the expression of feeding relevant neuropeptides, it is necessary to develop an in vitro system for direct cellular level studies. It has been shown that the N1E-115 mouse neuroblastoma cell line expresses the gene product for neuropeptides, including AGRP, that play a role in feeding behavior (15). N1E-115 neuroblastoma cells have been used as a neuronal model system. These cells also have been used to study polyunsaturated fatty acid modulation of cAMP formation (23) and insulin-stimulated myoinositol uptake (24). Raising the glucose concentration from 5.6 to 25 mmol/liter was associated with decreased myoinositol uptake, with an inhibitory constant (Ki) of 20.4 mmol/liter for N1E-115 cells (25). These characteristics indicate the potential utility of the N1E-115 neuroblastoma cell line as a model for studies of nutrients regulation of expression of feeding relevant neuropeptides. In addition, Gt1-7, a mouse hypothalamic neuronal cell line, has been known to express AgRP mRNA (16).
Using two mouse neuroblastoma cell lines, we demonstrated for the first time that glucose directly regulates the expression of AgRP. In addition, two cell lines sharply increased cellular ATP concentrations in a range of 1–5 mm glucose concentration, and thereafter the cellular ATP concentrations were saturated. It is noteworthy to mention that extracellular glucose concentration of the brain is approximately 2.4 mm in normoglycemia and that glucose concentration is maintained in a range of 0.2–4.5 mm even in hypo- and hyperglycemic states, whereas blood glucose concentrations vary in a much wider range of approximately 2.8–15.2 mm in hypoglycemia and hyperglycemia (26). In addition, degrees of AMPK phosphorylation are linearly decreased with increased glucose concentration at physiological levels. Therefore, minute changes in glucose concentration or energy status can be monitored and sensed by a delicate sensing system in the brain to generate hunger or satiety signals. Lastly, magnitudes of change in AgRP expression in response to glucose concentrations are similar to the degree of changes in AgRP expression in the arcuate during fasting and refeeding conditions (27). Our data on AgRP expression in two independent neuronal cell lines clearly demonstrated that these cells could respond to glucose deprivation or high levels of glucose, providing a reliable in vitro system to study the cellular mechanisms of nutrient regulation of AgRP expression. In addition, in vitro studies allow more direct testing of the effect of glucose on AgRP expression without the disadvantage of indirect effects brought about by interactions of cells with other areas of the brain and changes in blood hormones or substrates.
Inhibition of glucose use by 2-DG has been known to cause glucoprivic food intake and to increase AgRP and NPY expression in the arcuate (10, 11). However, it is not clear how glucoprivic conditions are converted to specific signals to AgRP neurons, which stimulate food intake. Our study showed that modulation of glucose availability in two cell lines could affect AgRP expression, indicating a direct effect of glucose and 2-DG on these neuronal cell lines. Because inhibition of glycolysis by 2-DG increased AgRP expression, it was hypothesized that glucose metabolites in the glycolysis pathway could be an effector molecule that is responsible for regulation of AgRP expression. To test this hypothesis, pyruvate was supplemented and suppressed a 2-DG-induced AgRP expression both in cell line cultures and hypothalamus culture ex vivo. These findings suggested that downstream metabolites of the glycolysis pathway beyond pyruvate or energy status might be responsible for regulation of AgRP expression.
Our studies clearly show that glucoprivic conditions elicit low cellular ATP concentration and that direct modulation of ATP synthesis regulates AgRP expression. In addition, expression of dominant negative AMPK in the neuronal cell line blocked low-glucose-induced AgRP expression and attenuated 2-DG-induced AgRP expression. These findings indicate that increased expression of AgRP in glucoprivic conditions is, at least partially, mediated through the regulation of AMPK.
It is possible that depletion of ATP in the hypothalamus of 2-DG-treated rats can be rescued by pyruvate supplementation, which readily generates enough ATP through the tricarboxylic acid cycle to normalize the energy state, thereby resulting in the suppression of 2-DG-induced food intake for the first 30 min. However, the lack of supplemented pyruvate that may occur during the 30- to 60-min period probably resumed 2-DG-induced food intake. Therefore, it will be interesting to investigate whether a continuous supply of pyruvate in the brain can delay glucoprivic food intake over a long-term period. Taken together, our in vitro, ex vivo, and in vivo studies provide strong evidence that restoring neuronal energy status by pyruvate supplementation decreased AgRP expression and food intake through the phosphorylation states of AMPK.
Because AgRP expression is colocalized with NPY expression in the same neurons of the arcuate, NPY expression can be also regulated in the same manner as AgRP within the same cells. Acute ip administration of 2-DG has been known to increase food intake and both AgRP and NPY expression in the arcuate (10). Most recent studies also show that adenoviral expression of AMPK in the arcuate increased food intake and expression of these orexigenic peptides (28). Therefore, it is likely that in response to glucose fluctuation, the metabolic and nutritional signals in the NPY/AgRP neurons can be generated and converted into the secondary signals that possibly share the same initiation steps. But the common signals, for example AMPK activities, need to be diverged to regulate two independent expressions of genes unless these promoters contain the same response elements. It will be interesting to investigate the downstream signaling pathway of AMPK connecting AMPK to the regulation of AgRP and NPY gene expression.
In agreement with our studies, several reports show that peripheral administration of lactate and pyruvate decreased food intake in rats (29). In fact, lactate is produced by astrocytes and glial cells in the brain and converted to pyruvate by lactate dehydrogenase (20), providing an energy source to neighboring neuronal cells. In addition, blood lactate and pyruvate can be transported across the blood-brain barrier (30) and used as alternative energy sources during periods of decreased glucose availability. Pyruvate supplementation has been known to increase ATP content and enhance energetic status during brain hemorrhagic shock (31). Based upon our findings and previous reports, we hypothesized that modulation of nutrient availability and use would affect food intake through changes in neuronal energy status. Consistent with our hypothesis, ICV infusion of brain fuels, including glucose, glycerol, and β-hydroxybutyrate, reduced food intake (32), and mercaptoacetate, an inhibitor of fatty acid oxidation, caused lipoprivic feeding (10). These studies further support our conceptual hypothesis. However, additional studies revisiting these experiments from the view of energy status will reconcile new concepts and help to better understand mechanisms of energy-privic feeding.
As a conclusion, our studies provide direct evidence of the critical role played by neuronal cell energy status in the activation and expression of AMPK, modulation of NPY and AGRP expression, and subsequent food intake. The data support our overall hypothesis that energy status of the nutrient-sensing cells of the brain is the major switch for the control of neuropeptide expression and feeding behavior. Manipulation of the energy status of the cells included modification of the rate of neuronal glucose, pyruvate oxidation, and direct manipulation of ATP concentration.
Fairly recent reports show that inhibition of β-oxidation by C75 administration generates low energy status, which stimulates food intake through AMPK (33). In addition, most recent studies show that both adenoviral expression of AMPK in the arcuate and ICV injection of AICAR increased food intake, suggesting involvement of AMPK in food intake regulation (28, 34). Our studies provide strong evidence that glucose, a major cellular energy source, regulates food intake through the cellular ATP status-AMPK axis. Therefore, the final common pathway for sensing glucose, fatty acids, and other nutrients can be merged into the energy status of the cell. These key findings should provide novel intervention opportunities for manipulation of feeding behavior.
Acknowledgments
We are grateful to Dr. Morris Birnbaum (University of Pennsylvania) for generously providing vectors containing AMPKα2WT and AMPKα2KD and to Dr. Pamela Mellon (University of California, San Diego) for providing the GT1-7 cell line.
This study was supported in part by Biotechnology for Students and Teachers program of LSU AgCenter and the LSU Pennington Biomedical Research Center.
Abbreviations
- AgRP,
Agouti-related protein;
- AICAR,
5-aminoimidazole-4-carboxamide 1-β-d-ribofuranoside;
- AMPK,
AMP-activated protein kinase;
- 2-DG,
2-deoxy-d-glucose;
- FBS,
fetal bovine serum;
- ICV,
intracerebroventricular;
- NPY,
neuropeptide Y;
- POMC,
proopiomelanocortin.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}