





Abstract
Barred Owls (Strix varia) have expanded their range throughout the ranges of Northern (Strix occidentalis caurina) and California Spotted Owls (S. o. occidentalis). Field observations have suggested that Barred Owls may be behaviorally dominant to Spotted Owls. Therefore, we conducted a test of behavioral dominance by assessing responsiveness of Spotted Owls to conspecific calls when they were in the simulated presence (i.e., imitation of Barred Owl vocalizations) of a Barred Owl. We hypothesized that Spotted Owls would be less likely to respond to conspecific calls in areas where Barred Owls were common. We used a binary 2 × 2 crossover experimental design to examine male Spotted Owl responses at 10 territories randomly selected within two study areas that differed in abundance of Barred Owls. We also conducted a quasi experiment at four study areas using response data from any Spotted Owl (male or female) detected following exposure to Barred Owl calls. We inferred from the crossover experiment that the simulated presence of a Barred Owl might negatively affect Spotted Owl responsiveness. Both subspecies of Spotted Owl responded less to Spotted Owl calls after exposure to Barred Owl calls, Northern Spotted Owls responded less frequently in areas having higher numbers of Barred Owls, and California Spotted Owls responded less frequently than Northern Spotted Owls overall.
¿Es la Emisión de Llamados de Strix Occidentalis Suprimida por la Presencia de S. varia?
La especie Strix varia ha extendido su distribución a través de los rangos de S. occidentalis caurina y S. o. occidentalis. Algunas observaciones de campo han sugerido que los individuos de S. varia podrían ser comportalmente dominantes sobre los individuos de S. occidentalis. Por lo tanto, realizamos una prueba de la dominancia comportamental que consistió en determinar la propensión de S. occidentalis a responder a llamados coespecíficos cuando se simulaba la presencia de S. varia mediante la imitación de sus vocalizaciones. Planteamos la hipótesis de que los individuos de S. occidentalis serían menos propensos a responder a los llamados coespecíficos en áreas donde S. varia era común. Empleamos un diseño experimental binario cruzado de 2 × 2 para examinar las respuestas de los machos de S. occidentalis en 10 territorios elegidos al azar en dos áreas de estudio que diferían en la abundancia de S. varia. También realizamos un semi-experimento en cuatro áreas de estudio usando datos sobre las respuestas de cualquier individuo macho o hembra de S. occidentalis detectado luego de ser expuesto a los llamados de S. varia. A partir del experimento cruzado, inferimos que la presencia simulada de S. varia podría afectar negativamente la propensión de los individuos de S. occidentalis a responder. Ambas subespecies de S. occidentalis respondieron menos a los llamados de su especie luego de la exposición a llamados de S. varia. Los individuos de S. o. caurina respondieron con menor frecuencia en áreas con números mayores de S. varia y, en general, los individuos de S. o. occidentalis respondieron menos frecuentemente que los de S. o. caurina.
Introduction
Barred Owls (Strix varia) have recently expanded their geographic range, and can now be found throughout the entire range of the Northern Spotted Owl (Strix occidentalis caurina; Dark et al. 1998, Kelly et al. 2003, Courtney et al. 2004). Because the Northern Spotted Owl is federally listed as a threatened species (U.S. Department of the Interior 1990), there is concern that Barred Owls may be displacing Northern Spotted Owls (Kelly et al. 2003, Courtney et al. 2004). The Barred Owl has also invaded the range of the California Spotted Owl (S. o. occidentalis) in the Sierra Nevada (Dark et al. 1998). However, the number of Barred Owls in the central Sierra Nevada is low, with their most southerly record consisting of two “Sparred” Owls (Spotted × Barred Owl hybrids) in the central Sierra Nevada (Seamans et al. 2004; MLC, pers. obs.) and one Barred Owl in the southern Sierra (G. Steger, Pacific Southwest Research Station, pers. comm.).
Interspecific territoriality and agonistic behavior among raptors has been well documented (Janes 1985). However, there is little evidence documenting the outcome of Spotted Owl encounters with Barred Owls (Courtney et al. 2004). Anecdotal evidence suggests that Spotted Owls are negatively affected by Barred Owls because Barred Owls probably compete with Spotted Owls for habitat and prey (Hamer 1988, Dunbar et al. 1991, Herter and Hicks 2000, Courtney et al. 2004), hybridize with Spotted Owls (Hamer et al. 1994, Dark et al. 1998), and may opportunistically prey upon Spotted Owls (Leskiw and Gutiérrez 1998). In Oregon and Washington, Spotted Owls declined after Barred Owls were detected within 0.8 km of their territory centers (Kelly et al. 2003). However, Spotted Owls may be declining for other reasons such as habitat loss or poor environmental conditions (e.g., bad weather; Franklin et al. 2000, Seamans et al. 2002, Kelly et al. 2003). In addition, Kelly et al. (2003) assumed that detection probability of Spotted Owls was constant regardless of the presence of Barred Owls, and they noted that this assumption was probably violated. Some biologists have suggested that Spotted Owls have changed their behavior in response to negative interactions with these new potential competitors by responding less to heterospecific and conspecific calls (Courtney et al. 2004). Although Barred Owls respond regularly to Spotted Owl calls (Herter and Hicks 2000), we did not know if the converse was true. Therefore, we experimentally evaluated Spotted Owl response following exposure to Barred Owl vocalizations in areas where Barred Owls are common (i.e., Spotted Owls are “experienced” with Barred Owls) and where they are rare (i.e., Spotted Owls are “naive”). The specific objective of our study was to determine whether imitating Barred Owl calls within known Spotted Owl territories caused a change in Spotted Owl responsiveness to conspecific calls, and, if so, whether such a difference could be attributed to Spotted Owl experience with Barred Owls. Quantifying changes in Spotted Owl responsiveness will help biologists account for changes in detection probabilities in demographic models. In addition, detectable changes in behavior can increase our understanding of interference competition between Barred and Spotted Owls.
Methods
Study Areas
We conducted this experiment at four long-term Spotted Owl demographic study areas in California (Eldorado), Oregon (Klamath and Corvallis), and Washington (Cle Elum; Fig. 1). The geographic range of these study areas was large, and included the following counties: Placer and Eldorado (Eldorado), Douglas (Klamath), Lane, Polk, and Benton (Corvallis), and Kittitas (Cle Elum). Study areas differed in climate, elevation, and vegetation (Forsman et al. 1996, Seamans et al. 2001), but all were in mountainous regions, predominantly on public land, and were dominated by coniferous forest in various seral stages. Study areas also differed by the subspecies of Spotted Owl inhabiting the area and the prevalence of Barred Owl detections (Table 1). We considered the proportion of historic Spotted Owl territories in which Barred Owls were detected during the 2004 field season as a gross index of Barred Owl abundance on each study area. Ten territories occupied by male Spotted Owls were selected for inclusion in the Barred Owl calling experiment from each of these areas except Cle Elum, where only six were used. Territories were located within or adjacent to the demography study areas.
Locations of long-term Spotted Owl demographic study areas in California, Oregon, and Washington.
Spotted Owl territories used in this study were associated with these areas. Ten territories each were surveyed at the Eldorado, Klamath, and Corvallis study areas, while six territories were surveyed at the Cle Elum study area.
Characteristics of the Spotted Owl demographic study areas used in the true (random allocation of treatment with male vocal response) and quasi experiments (nonrandom allocation of treatment with male or female vocal response) to assess the effect of Barred Owl vocalizations on Spotted Owl short-term responsiveness. Barred Owl detection is defined as the proportion of historic Spotted Owl territories in each study area where one or more Barred Owls were detected in the year of our study (2004).

Characteristics of the Spotted Owl demographic study areas used in the true (random allocation of treatment with male vocal response) and quasi experiments (nonrandom allocation of treatment with male or female vocal response) to assess the effect of Barred Owl vocalizations on Spotted Owl short-term responsiveness. Barred Owl detection is defined as the proportion of historic Spotted Owl territories in each study area where one or more Barred Owls were detected in the year of our study (2004).
Field Methods
We examined whether the response frequency of naive and experienced Spotted Owls to simulated Spotted Owl calls differed after exposure to simulated Barred Owl calls. We had initially planned to conduct the experiment in exactly the same manner at all four sites, but logistical constraints (e.g., shortage of personnel) required selection of territories that were geographically closer to the base of operations or fewer in number. Therefore, to evaluate Spotted Owl responses we conducted both a true experiment (territories selected randomly from the study population) and a quasi experiment (territories either not selected randomly or the experiment concluded when either sex of territorial Spotted Owl responded). In both cases, the experimental unit was the individual Spotted Owl, the measurement unit was the detection survey at the Spotted Owl territory, and the treatment was the imitation of Barred Owl calls within the Spotted Owl territory. For the true experiment, we generated a list of territories that were known to be occupied by a Spotted Owl male or pair in the year of our study (2004) and that were located within 1 km of a road or trail. The latter constraint was minor (most territories are within this distance of a trail or road) but important, because otherwise we could not easily execute the individual calling surveys within the specified time period (see below). From this list, we randomly selected 10 territories per study area. These territories were then randomly assigned to one of two groups to receive treatment in one of two experimental survey periods. For the quasi experiment, all features of the study design were the same except that: (1) imitations of Spotted Owl calls were discontinued when either a male or female Spotted Owl responded, and (2) treatment territories were not randomly selected. Although we knew that the Eldorado study area had fewer Barred Owls than the study areas occupied by Northern Spotted Owls, we had no a priori knowledge of Barred Owl detection rates at the three Northern Spotted Owl study areas. Thus, this did not influence selection of the two study areas used in the true experiment. To simulate Barred Owl vocalizations, we imitated the territorial calls of the Barred Owl using a mechanical device at six evenly spaced call points within the Spotted Owl territories. These call points were selected based on long-term knowledge of owl responses at that territory, and hence were placed in such a fashion as to enhance the possibility of an owl hearing a vocalization in that territory. We surveyed each Spotted Owl territory prior to experimentation to demonstrate Spotted Owl occupancy in the territory. Treatment surveys consisted of a complete survey imitating Barred Owl calls at the six call points, followed by a second survey where we returned to the six call points in the same sequence and imitated Spotted Owl calls using the same mechanical device (the devices were used to provide consistency among observers). For the control surveys, we imitated Spotted Owl calls only at the six call points established for each territory. Each territory was surveyed once per experimental survey period. Surveys were conducted from 21 July to 12 August 2004. The two experimental survey periods were separated by ≥2 weeks to minimize carryover effects of treatments applied during the first experimental survey period, although we also tested for carryover effects in our analysis.
Crossover Design
We used a 2 × 2 binary crossover design to conduct these experiments (Senn 1993). Our crossover design consisted of: (1) randomly applying the treatment (i.e., imitating Barred Owls calls) to half the experimental units during the first survey period and using the other half as controls (i.e., imitating Spotted Owl calls only), then (2) switching treatment and control units for the second survey period. We tested if Barred Owl presence (i.e., imitated Barred Owl calls) had an effect on Spotted Owl responsiveness by comparing Spotted Owl response to conspecific calls after they had been exposed to simulated Barred Owl vocalizations (treatment surveys) and had not been exposed to simulated Barred Owl vocalizations (control survey where only Spotted Owl calls were imitated). This design allowed us to use individual owls as their own controls, thus controlling for variation among experimental units (Ratkowski et al. 1993, Senn 1993). Assignment to treatment groups was random, but spatial constraints determined the order of visits to territories within periods (territories within the same general area were surveyed on the same night). The survey order established for the first experimental period was followed in the second experimental period so that the time between surveys at each territory was similar. We used only male Spotted Owl response as the variable of interest in the true experiment because males are more likely to respond to intruding owls than females (Reid et al. 1999). We used the detection of a male or female as the response variable in the quasi experiment.
In a similar study of Great Horned (Bubo virginianus) and Spotted Owls (Crozier et al. 2005) we guarded against the threat of predation and other confounding effects by waiting 24 hr between Great Horned and Spotted Owl surveys. In this study we were less concerned about predation because Barred Owls are rare in the Eldorado study area (Seamans et al. 2004) and because there is only circumstantial evidence of Barred Owl predation on Spotted Owls in the Pacific Northwest (Leskiw and Gutiérrez 1998). To avoid confounding effects of imitating both species' calls from the same point in immediate succession (i.e., we would not know to which species the Spotted Owl was responding), we first imitated Barred Owl calls at all six points in sequence, and then returned to the starting point and resurveyed the call points in the same sequence using Spotted Owl calls.
Imitated Call Survey Design
Within each territory, we established six survey (call) points approximately 0.4–0.6 km apart to obtain consistent survey coverage. We surveyed for owls using methods similar to those of Forsman (1983) for Spotted Owls and McGarigal and Fraser (1984, 1985) for Barred Owls. Barred Owl surveys were extended to 10 min to standardize survey effort between the species. A complete survey consisted of the combined results of all individual call points from one survey period within a territory. Treatments were composed of two parts, Barred Owl calls followed by Spotted Owl calls. In contrast, control surveys consisted of imitated Spotted Owl calls only. If a male Spotted Owl responded to our imitation of Spotted Owl calls, whether during treatment or control surveys, we did not continue to call at remaining call points because we were interested in the outcome of surveys, not individual call points.
We conducted calling surveys from 30 min after sunset to approximately 01:00 PST to reduce within-night variation in responsiveness (Forsman 1983). We did not conduct surveys if either the wind was >12 km hr−1 or it was raining (Forsman 1983). During treatment surveys, we imitated Barred Owl hoots (McGarigal and Fraser 1984, 1985, Laidig and Dobkin 1995) using Primos owl calls (Primos, Inc., Flora, Mississippi). We used these devices to consistently reproduce the same vocalization among study areas and surveys. For the first min at each call point observers listened for unsolicited calls (Johnson 1993). For the next 6 min we imitated Barred Owl hoots. A Barred Owl imitation consisted of approximately six sets of 10 sec ascending (whoo-hoo-hoo-hoo-hoo-hoo-hoo-hoo whoo-whaaaaa) or typical nine-note (“Who cooks for you? Who cooks for y'all,” which is the most common Barred Owl call [McGarigal and Fraser 1985]) calls, each separated by a 10 sec pause (McGarigal and Fraser 1984, 1985, Laidig and Dobkin 1995). We imitated the first 10 sec calling bout perpendicular to the road, and then rotated 180° following each 10 sec imitation. After the final imitation, 3 min were spent at the call point listening and recording owl responses. During Spotted Owl surveys, we used the same Primos owl calls to imitate Spotted Owls for 10 min, producing 3–5 typical four-note location calls every 15 sec. The 10 sec and 15 sec intervals of silence between Barred Owl and Spotted Owl calls, respectively, represented the frequency of unsolicited calls observed under natural conditions for each species (Spotted Owl: Forsman et al. 1984, Johnson 1993; Barred Owls: McGarigal and Fraser 1985, Mazur and James 2000). We considered a positive survey response for a territory to be any survey in which a Spotted Owl was detected at any call point during the Spotted Owl imitation part of the survey (i.e., if a Spotted Owl was detected during Barred Owl imitations it was not considered a positive response). For each owl that was detected, observers noted the time of detection, sex (based on pitch of call; Forsman 1983, Mazur and James 2000), response type (visual or vocal), and the compass direction and distance to the owl. We also recorded the location, date, time, temperature, wind, cloud cover, and precipitation at each call point (Fuller and Mosher 1981). We noted all Barred Owl detections during these surveys.
Statistical Analyses
We developed a suite of a priori hypotheses (models) to explain how imitating Barred Owl calls might affect short-term responsiveness of both naive and experienced Spotted Owls (Table 2). We used the experimental PROC GLIMMIX to analyze our models (SAS Institute 2004a, 2004b), which were mixed models with a binary response (0 or 1). We estimated variation in individual owl responsiveness by considering each Spotted Owl territory a random effect in all models. Treatment, experience (proportion of historic Spotted Owl territories in study areas in which one or more Barred Owls were detected in 2004; Table 1), subspecies, structural components of the study design (carryover and period), and detection of a Barred Owl(s) during call imitations were fixed effects. However, only two Barred Owl detections occurred during 72 treatment and control surveys in territories occupied by a male Spotted Owl, so we were unable to test the effect of a Barred Owl actually being detected within a Spotted Owl territory. We analyzed data within a pseudolikelihood framework using a generalized linear mixed model (GLMM) with a logit link function and binomial error because our response variable was binary (no response = 0, Spotted Owl vocal response = 1). Pseudoestimation techniques allowed us to achieve more accurate parameter estimates than other techniques (SAS Institute 2004b; V. Agboto, University of Minnesota, Department of Statistics, pers. comm.). However, it was inappropriate to use pseudo-AIC values to compare models in a model selection context, so we present means ± SE, and F- and P-values. At least in the case of the true experiment, presentation of probability values is appropriate.
A priori models used to evaluate the effects of treatment (imitating Barred Owl calls) on responsiveness of male Spotted Owls to conspecific calls. All models are generalized linear mixed models (GLMMs) in which territory (TERR) was considered a random effect and all other variables were fixed effects. T, EXP, SUBSP, P, and C indicate Treatment, Experience, Subspecies, Period, and Carryover, respectively. Each model included an intercept. K indicates the number of model parameters. Experience models were used only for comparison of all four study areas for the quasi experiment. All other models were used in analysis of both the true and quasi experiments.
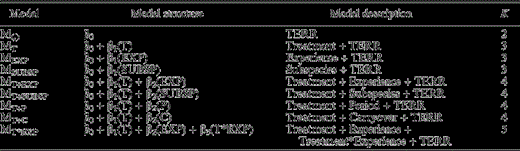
A priori models used to evaluate the effects of treatment (imitating Barred Owl calls) on responsiveness of male Spotted Owls to conspecific calls. All models are generalized linear mixed models (GLMMs) in which territory (TERR) was considered a random effect and all other variables were fixed effects. T, EXP, SUBSP, P, and C indicate Treatment, Experience, Subspecies, Period, and Carryover, respectively. Each model included an intercept. K indicates the number of model parameters. Experience models were used only for comparison of all four study areas for the quasi experiment. All other models were used in analysis of both the true and quasi experiments.
Results
True Experiment
None of the fixed parameters in any of the models were statistically significant (P > 0.05 for all parameters; Table 3). However, male Spotted Owl response to Spotted Owl calls occurred less frequently following imitation of Barred Owl calls at both study areas (Fig. 2). The California Spotted Owl was less responsive than the Northern subspecies during both treatment and control surveys, but again the difference was not statistically significant (Table 3, Fig. 2). We could not discriminate between the effects of subspecies and experience with Barred Owls because these two factors were confounded. Variables for carryover and period effects explained little variation in male Spotted Owl response frequency.
Estimates of fixed parameters with associated standard errors (SE), degrees of freedom (df), F-values (calculated from Type III sums of squares) and P-values for hypothesized models explaining short-term responsiveness of male Spotted Owls to Spotted Owl calls after exposure to Barred Owl calls in the central Sierra Nevada, California (California Spotted Owl, CSO) and Coast Ranges, Oregon (Northern Spotted Owl). Estimates of fixed parameters for the true experiment were not statistically different from zero for any models (α = 0.05). For individual parameters, estimates represent the probability that Treatment (T) = control, Subspecies (SUBSP) = CSO, Period (P) = first, and Carryover (C) = no.
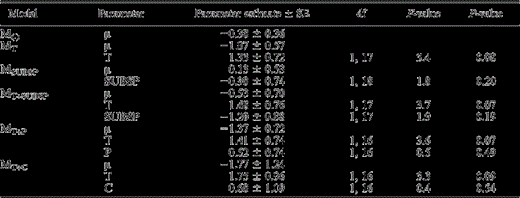
Estimates of fixed parameters with associated standard errors (SE), degrees of freedom (df), F-values (calculated from Type III sums of squares) and P-values for hypothesized models explaining short-term responsiveness of male Spotted Owls to Spotted Owl calls after exposure to Barred Owl calls in the central Sierra Nevada, California (California Spotted Owl, CSO) and Coast Ranges, Oregon (Northern Spotted Owl). Estimates of fixed parameters for the true experiment were not statistically different from zero for any models (α = 0.05). For individual parameters, estimates represent the probability that Treatment (T) = control, Subspecies (SUBSP) = CSO, Period (P) = first, and Carryover (C) = no.
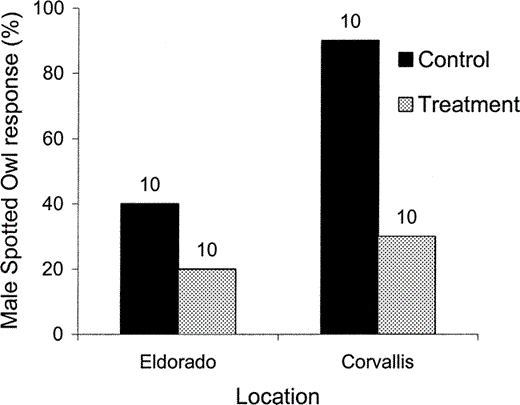
Male Spotted Owls responded less frequently during Treatment (Barred Owl followed by Spotted Owl imitations) than during Control (Spotted Owl imitations only) surveys during a crossover experiment, although this was not statistically significant.
Also, male Spotted Owls responded less frequently at the Eldorado study area in California, inhabited by California Spotted Owls and few Barred Owls, than at the Corvallis study area in Oregon, inhabited by Northern Spotted Owls and where Barred Owls are more common. The y-axis represents the percentage of surveys in which a male Spotted Owl responded. Numbers above bars represent sample size.
Quasi Experiment
Results from the quasi experiment showed similar patterns to those of the true experiment, but estimates of some fixed parameters were statistically significant. Spotted Owl response to Spotted Owl calls following imitation of Barred Owl calls was less frequent than Spotted Owl response following imitation of only Spotted Owl calls for all experimental groups (Fig. 3). In addition, California Spotted Owls were less responsive than Northern Spotted Owls, and Northern Spotted Owls were less responsive in areas with more Barred Owls. Estimates of fixed parameters were significant for treatment and subspecies, but not experience (Table 4). Variables for carryover and period effects explained little variation in Spotted Owl response frequency.
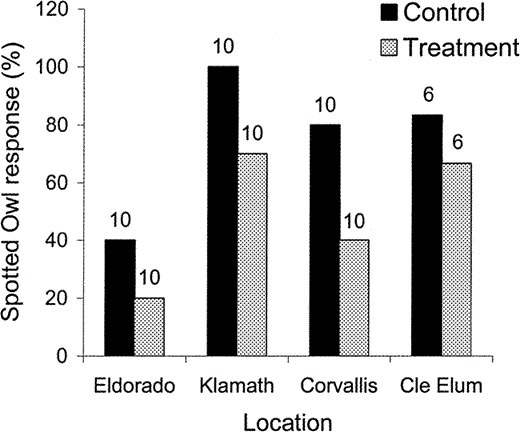
Spotted Owls (male or female) responded less frequently during Treatment (Barred Owl followed by Spotted Owl imitations) than during Control (Spotted Owl imitations only) surveys during a quasi experiment.
Also, Spotted Owls responded less frequently at the Eldorado study area in California, inhabited by California Spotted Owls and few Barred Owls, than at the Klamath and Corvallis study areas in Oregon or the Cle Elum study area in Washington, inhabited by Northern Spotted Owls and where Barred Owls are more common. The y-axis represents the percentage of surveys in which a Spotted Owl of either sex responded. Numbers above bars represent sample size.
Estimates of fixed parameters with associated standard errors (SE), degrees of freedom (df), F-values (calculated from Type III sums of squares) and P-values for hypothesized models explaining short-term responsiveness of male and female Spotted Owls to Spotted Owl calls after exposure to Barred Owl calls in the central Sierra Nevada, California (Eldorado; California Spotted Owl, CSO) or Pacific Northwest (Klamath, Corvallis, and Cle Elum; Northern Spotted Owl). Estimates of fixed parameters for the quasi experiment were statistically different from zero for treatment and subspecies effects (α = 0.05). For individual parameters, estimates represent the probability that Treatment (T) = control, Subspecies (SUBSP) = CSO, Period (P) = first, and Carryover (C) = no.
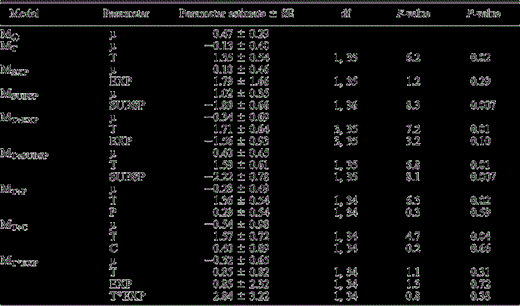
Estimates of fixed parameters with associated standard errors (SE), degrees of freedom (df), F-values (calculated from Type III sums of squares) and P-values for hypothesized models explaining short-term responsiveness of male and female Spotted Owls to Spotted Owl calls after exposure to Barred Owl calls in the central Sierra Nevada, California (Eldorado; California Spotted Owl, CSO) or Pacific Northwest (Klamath, Corvallis, and Cle Elum; Northern Spotted Owl). Estimates of fixed parameters for the quasi experiment were statistically different from zero for treatment and subspecies effects (α = 0.05). For individual parameters, estimates represent the probability that Treatment (T) = control, Subspecies (SUBSP) = CSO, Period (P) = first, and Carryover (C) = no.
Spotted Owl Response to Barred Owl Calls
Male Spotted Owls did respond to Barred Owl calls. In fact, during treatment surveys, Spotted Owls sometimes responded to imitations of Barred Owls during the first part of the survey, but failed to respond to Spotted Owl calls during the second part of treatments. For example, at the Corvallis study area, a male Spotted Owl responded during Barred Owl imitations at six of 10 territories, and then responded to subsequent imitated Spotted Owl calls at only four of these territories. At the Eldorado study area, male Spotted Owls responded during Barred Owl surveys at three of 10 territories, but only one of these males responded during the subsequent Spotted Owl surveys. In the quasi experiment, male and female Spotted Owls responded similarly; some Spotted Owls responded to Barred Owl calls, but failed to respond to subsequent Spotted Owl calls. At one territory in the Klamath study area, a Barred Owl responded to the Barred Owl calls during treatment and also engaged in a calling bout with the resident territorial Spotted Owl during the control survey. At the Corvallis study area, both a Spotted and Barred Owl were detected during a control survey. However, this Barred Owl was not detected during the treatment survey, when Barred Owl calls were imitated.
Discussion
There was evidence suggestive of a treatment effect in the true experiment (i.e., responses fewer but not statistically significant, P = 0.08) and there was a significant treatment effect in the quasi experiment, thus Barred Owl calling appeared to decrease Spotted Owl responsiveness to conspecific calls. This suggests that there is behavioral suppression of Spotted Owls by Barred Owls. These results also suggest that presence of Barred Owls likely affects Spotted Owl detection probabilities and should be considered when drawing inferences about Spotted Owl surveys conducted in areas that are also occupied by Barred Owls. For example, Kelly et al. (2003) assumed that Spotted Owls could be detected even if they occurred in areas where Barred Owls were present, but noted that this assumption was probably violated because Spotted Owls may be less likely to respond to vocal imitations or recordings if Barred Owls are present. If the presence or absence of Barred Owls is to be included as a covariate in Spotted Owl demographic models, directly surveying for Barred Owls could be useful as Barred Owl detections could be used as an uncorrected index of presence or absence. However, using actual Barred Owl surveys to detect Barred Owls in these areas might have the confounding effect of reducing Spotted Owl responsiveness. Thus, we recommend separating species-specific surveys for these two owls in time and, if possible, space (i.e., do not use Barred Owl call points in areas occupied by Spotted Owls). Alternatively, Barred Owl detections recorded during Spotted Owl surveys could be used as an index of presence or absence, but such an index should be examined for its efficacy in each study area.
Discrepancies between the true and quasi experiments cannot be directly explained, but since trends in raw data were similar, variation in statistical significance might be somewhat explained by differences in sample size. We decided to analyze these data sets separately because owls at two areas were randomly selected whereas at the other two study areas they were not. In addition, there was a smaller sample of owls from the Cle Elum study site. We have no a priori reason to believe that the nonrandom selection of sites biased these particular results (i.e., they were not selected for a specific reason other than to compensate for logistics); thus, we attribute differences between the two data sets primarily to sample size variation.
Of interest is that the same trend observed in Northern Spotted Owl study sites was also noted in the Eldorado study area, where there were almost no Barred Owls (one pair of hybrids in the entire study area of 365 km2). In fact, naive California Spotted Owls were less responsive than experienced Northern Spotted Owls. Differential responsiveness to vocal imitations between subspecies has been previously noted for surveys conducted during daylight (Gutiérrez et al. 1995), but was larger than we expected. This suggests either that Spotted Owls may simply be less responsive in the presence of a novel owl species or that California Spotted Owls behave differently. We speculate that negative interactions between the species may serve to increase the reticence of Spotted Owls in vocalizing when Barred Owls are present.
The fact that Spotted Owls responded to Barred Owl calls argues against our speculation. However, the nature of these species interactions may be complex. We expected to detect Barred Owls if they were present because previous studies have noted that they readily respond to both conspecific and Spotted Owl calls (McGarigal and Fraser 1985, Herter and Hicks 2000). Although Barred Owls were detected with some regularity across three of the four long-term Spotted Owl demography study areas in 2004, only two Barred Owls were detected in the occupied Spotted Owl territories used in this study. This may indicate possible avoidance or spatiotemporal partitioning of areas between the two species (Laidig and Dobkin 1995). In contrast, Great Horned Owls were often detected in Spotted Owl territories during a similar experimental study (Crozier et al. 2005). Our results suggest that interspecific competition between Barred and Spotted Owls might occur, and that calling by Spotted Owls was altered after encountering calls by Barred Owls. Because responsiveness of Spotted Owls was different between treatment and control surveys following imitation of Barred Owl calls during treatment surveys, we infer that Barred Owl calls caused Spotted Owls to alter their behavior, but the causal mechanism is unknown (behavior suppression or novel stimulus).
Courtney et al. (2004) summarize the potential negative interactions between Spotted and Barred Owls. Field biologists have hypothesized one such interaction, which is that Spotted Owls might be less responsive when Barred Owls are present. If true, this might negatively affect Spotted Owls because they use vocalizations to establish and maintain breeding territories. Our results support this hypothesis, but it is not entirely clear why Spotted Owls were less responsive in an area where they had little or no exposure to Barred Owls. Future studies of interactions between these species should focus on experiments, refining experimental protocols for behavioral studies such as ours, and examining direct interactions between these two species.
Acknowledgments
We thank the USDA Forest Service, the U.S. Fish and Wildlife Service, and the U.S. Bureau of Land Management for providing support. Vincent Agboto, University of Minnesota, Department of Statistics provided valuable insight into pseudoestimation techniques. We are grateful to the many field technicians who assisted in collecting the data associated with this study: C. Barclay, S. Brown, S. Dallmann, S. Fiala, M. Force, C. Hansen, J. Lowden, V. Pedro, C. Seck, A. Sidie, M. Taylor, and H. Wise. Special thanks to Tom Snetsinger, who conducted surveys, helped with logistics, and reviewed a draft of this paper. Lorelle Berkeley also reviewed a draft of this paper. George Steger, Pacific Southwest Research Station, kindly reported the presence of a Barred Owl in the Southern Sierra. This study was funded by Region 5, USDA Forest Service (Contract #53-91S8-5-EC15 to RJG), the Pacific Northwest Research Station, and the Bureau of Land Management through the Regional Interagency Monitoring Program.
Literature Cited
Author notes
Corresponding author. E-mail: [email protected]

{kind=link}
{kind=link}
{kind=link}