





Abstract
We investigated whether wing morphology differed between the sedentary House Finches (Carpodacus mexicanus) of western North America and the introduced population of eastern North America, as the latter has developed migratory behavior since its inception. Wing morphology differed between eastern and western House Finches. Eastern House Finches had shorter proximal primaries and a longer outer primary, perhaps reflecting a thinner and more pointed wing, although no disparity in wing length was detected. Since we interpret these differences in wing shape as modifications for flight capability, we believe that initial evidence for morphological divergence relative to migratory habit between eastern and western House Finches has been established here. Confirmatory studies to determine if wing morphology varies according to the gradient in expression of migratory behavior throughout the range of eastern House Finches are now warranted.
La Forma Alar en Carpodacus mexicanus Difiere en Relación a los Hábitos Migratorios entre el Este y Oeste de Norte América
Resumen. Investigamos si la morfología del ala difería entre individuos sedentarios de Carpodacus mexicanus del oeste de Norte América y la población introducida en el este de Norte América, la cual ha desarrollado un comportamiento migratorio desde su inserción. La morfología del ala difirió entre las poblaciones de C. mexicanus del este y del oeste. Los individuos del este tuvieron primarias proximales más cortas y primarias externas más largas, lo que quizás refleja un ala más fina y más puntiaguda, aunque no se detectó una diferencia en el largo del ala. Ya que interpretamos estas diferencias en la forma del ala como modificaciones para la capacidad del vuelo, creemos que se ha establecido una evidencia inicial de divergencia morfológica en relación con el hábito migratorio entre las poblaciones de C. mexicanus del este y del oeste de Norte América. Actualmente, son necesarios estudios que confirmen y determinen si la morfología del ala varía de acuerdo al gradiente de expresión del comportamiento migratorio a través del rango de distribución de la población de C. mexicanus del este.
One of the most notable ornithological events of the twentieth century in North America was the introduction of House Finches (Carpodacus mexicanus) into the eastern United States from native western populations and their subsequent spread (Hill 1993). The eastern population is believed to have originated from the release of a small but unknown number of captive birds on Long Island, New York, around 1940 (Elliot and Arbib 1953), and the source of the birds is presumed to be the Los Angeles area of southern California (Mundinger 1975). After a decade or so of local increases in the vicinity of western Long Island, the introduced population grew exponentially and rapidly expanded its range across the continent (Veit and Lewis 1996). House Finches are now naturalized and common throughout most of the eastern and midwestern United States, and in southern Canada.
As House Finches in the native range are essentially sedentary, one striking outcome of the introduction has been the appearance and increased frequency of migratory behavior in the eastern population of House Finches (Belthoff and Gauthreaux 1991, Able and Belthoff 1998). In fact, the eastern House Finch provides one of the very few cases in which the establishment of a large-scale pattern of migratory behavior has been documented under natural conditions in our lifetime. Over the 60 or so generations since their introduction, a complex system of seasonal migration has evolved in House Finches (Able and Belthoff 1998) in a remarkably brief and perhaps unprecedented period of time, something that studies on captive birds suggest can occur under strong selection (Berthold et al. 1990, Berthold 1996).
Wings of migrant species often are characterized by long distal primaries and short proximal primaries, resulting in relatively long and thin wings (compared with nonmigratory species; Rayner 1988, Winkler and Leisler 1992, Alerstam 1993, Lockwood et al. 1998), and a wing tip close to the leading edge. A pointed tip may reduce drag associated with the wing tip vortex (Lockwood et al. 1998). Even within species, comparisons between sedentary and migratory individuals or populations show that the migratory representatives indeed have wing features more characteristic of migration (Senar et al. 1994, Copete et al. 1999, Perez-Tris and Telleria 2001).
Wilson (1975) pointed out that behavior is the part of the phenotype that is most likely to change in response to long-term environmental change, such that behavior will usually be altered first and physical structure second. Thus, given that selection has apparently favored migratory behavior in eastern House Finches since their introduction, we investigated whether corresponding changes in wing morphology have arisen as well.
Methods
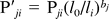
Statistical Analysis
Wing length in eastern and western House Finches was compared using ANOVA. We compared transformed primary lengths (P1′ through P9′) between sexes and populations using two-way multivariate analysis of variance (MANOVA) followed by inspection of univariate tests to determine which transformed primary lengths contributed to overall range differences. Because sex did not affect P1′ through P9′, and sex did not interact with range in either multivariate or univariate analyses, we pooled sexes for all subsequent analyses and used all 152 captured individuals. To help control for inflated Type I errors following the initial MANOVA, follow-up univariate ANOVAs were evaluated using sequential Bonferroni corrections (Rice 1989), in which alpha was 0.05/9 (= 0.0055) for the first comparison and sequentially adjusted. We conducted PCA on the standardized primary lengths and compared PC scores between House Finches in the eastern and western populations using ANOVA with sequential Bonferroni corrections. Analyses were conducted using SAS (Version 8.2, SAS Institute Inc. 2000), and all results are reported as means ± SE.
Results
Despite the prediction relative to migratory habit, there was no difference in mean wing length between the eastern (76.7 ± 0.2 mm) and western (76.9 ± 0.3 mm) House Finches in our sample (F1,150 = 0.2, P = 0.68). However, there was a significant difference in standardized primary lengths between populations (MANOVA: Wilks' lambda = 0.79, F9,142 = 4.1, P < 0.001). Follow-up univariate ANOVAs revealed that mean P2′ through P6′ were greater in western House Finches, whereas average P9′ was greater in eastern House Finches (Fig. 1).
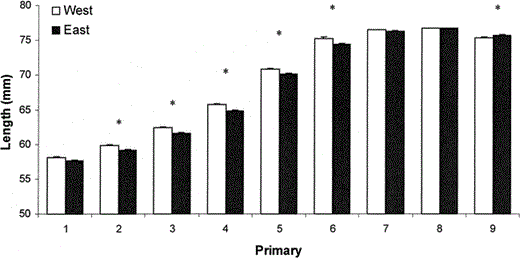
Standardized primary lengths of House Finches in two populations in the eastern and western United States (Ithaca, New York, n = 92 birds, and Boise, Idaho, n = 60 birds). Asterisks above bars indicate primaries for which the two samples differed significantly using one-way ANOVA and sequential Bonferroni corrections following a significant MANOVA
The first three principal components (PCs) explained 73% of the variation in transformed primary lengths (Table 1). The first axis, PC1, represented increasing proximal primary (P2′–P6′) lengths. PC2 represented increasing distal primary length (P8′ and P9′), and P6′ loaded most highly on PC3 (Table 1). Eastern and western House Finches differed significantly along PC1 (F1,150 = 25.9, P < 0.001). Eastern House Finches (PC1 = −0.64 ± 0.20) had shorter proximal primary lengths than western House Finches (PC1 = 0.98 ± 0.25), and this contributed to wings of eastern House Finches being thinner and somewhat more pointed (Fig. 2, 3). Eastern and western House Finches did not differ along PC2 (0.08 ± 0.12 and −0.12 ± 0.14, respectively; F1,150 = 1.1, P = 0.29) or along PC3 (−0.10 ± 0.10 and 0.16 ± 0.12, respectively; F1,150 = 2.6, P = 0.11).
Results of a principal components (PC) analysis on standardized primary lengths (P1′ through P9′) of House Finches from eastern and western North American populations (n = 152). P1′ represents standardized length of the most proximal primary
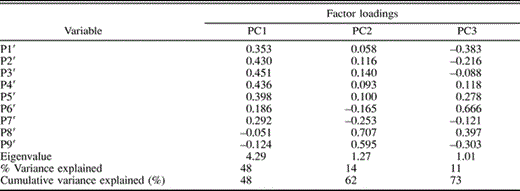
Results of a principal components (PC) analysis on standardized primary lengths (P1′ through P9′) of House Finches from eastern and western North American populations (n = 152). P1′ represents standardized length of the most proximal primary
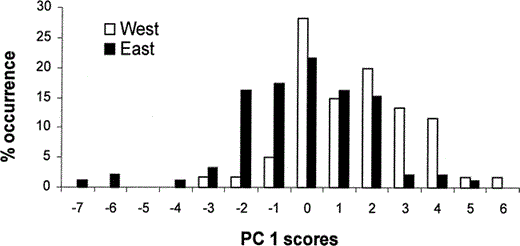
Frequency distribution (% occurrence) of eastern-U.S. (n = 92) and western-U.S. (n = 60) House Finches along the first axis generated by a principal components analysis of standardized primary lengths (Table 1). Increasing PC1 score corresponds to increasing lengths of proximal primaries
Graphical representation of distal wing segments produced using mean standardized primary lengths to visualize differences in wing morphology between eastern and western House Finches (note that these are not wing tracings). Wings are less rounded proximally (thinner) and more pointed distally in eastern House Finches, which appears to correlate with their more migratory habit
Discussion
Results of our study indicate that the wings of the more migratory eastern House Finches tended to be more pointed than those of western House Finches. The shorter proximal primaries of eastern House Finches perhaps reflected a thinner wing proximately. In accordance with another prediction of morphology based on migration, the transformed length of P9 also was significantly longer in eastern House Finches, which may represent the wing point being closer to the leading edge.
While we observed wing-shape differences consistent with each population's migratory habit, the predicted increase in the overall wing length of the more migratory (eastern) population was not observed. It is possible that this is related to the short amount of time that selection has had to operate on wing morphology. Longitudinal studies of wing morphology in House Finches could help document these changes over time. Alternatively, decreased wing lengths in at least one population of eastern House Finches have been documented after the recent mycoplasmal conjunctivitis outbreak (Nolan et al. 1998), so differences in wing lengths may have been more apparent before the outbreak. Finally, we were uncertain if the eastern House Finches we sampled were actually migratory or sedentary, as House Finches in the East are partial migrants (Belthoff and Gauthreaux 1991, Able and Belthoff 1998). A future study in which the migration distance of individuals is known might uncover a significant difference in wing length between the two populations. Nevertheless, the patterns of wing-shape differences between regions (east and west) that we observed despite the lack of differences in overall wing length indicate potential changes in wing morphology consistent with a more migratory habit in eastern House Finches.
An alternative hypothesis for the pattern of wing-shape differences we observed between eastern and western House Finches is that the measured morphological features of eastern House Finches also may be characteristic of the southern California source population from which the eastern population presumably arose. That is, southern California House Finches could have shorter proximal primaries and longer distal primaries than the sample of western House Finches we measured from Idaho for the current study, and these differences carried over via their introduction to the East.
In conclusion, we interpret the differences that were detected in wing shape between eastern and western House Finches as initial evidence for morphological divergence as a result of migratory habit. However, our study was limited to single populations in the East and the West. Confirmatory tests should include measurements of wing morphology in populations throughout both the sedentary and migratory ranges of House Finches. For instance, Able and Belthoff (1998) showed that populations colonized most recently, and farther from the origin of the eastern population, had the greatest propensity to migrate. Considering this gradient in the expression of migratory behavior, we hypothesize that wing morphology will vary accordingly within the eastern population of House Finches.
Acknowledgments
We thank the Cornell Laboratory of Ornithology's House Finch Project personnel, C. Moulton, and J. Soules for assistance in the field. We especially thank A. Dhondt for his assistance, hospitality, and many stimulating and interesting discussions. We also thank J. C. Senar and J. Lleonart for providing the SIZESTD program for use in this study, and J. C. Senar and two anonymous referees for helpful comments on our manuscript. Finally we thank Sigma Xi, The Scientific Research Society, and Boise State University for providing financial or logistical support. This research was completed in partial fulfillment of requirements for undergraduate research credit at Boise State University for JRE.
Literature Cited
Author notes
Corresponding author. [email protected]

{kind=link}
{kind=link}
{kind=link}