





Abstract
Bananaquits (Coereba flaveola) on the island of Grenada in the West Indies have a plumage color polymorphism in which individuals are either yellow and black or all black. In the southwest of the island there is a cline in plumage morphs in which the frequency of black individuals increases with distance from the island's southwestern tip. We describe the present position (September 2002) and form of this cline in comparison to when it was last mapped, 21 years ago. There has been no net movement of the cline during this period. We suggest that previous movement in the cline could have been related to long-term variation in rainfall.
Estasis en la Clina de Formas de Coereba flaveola en Grenada, Antillas
Resumen. Los individuos de Coereba flaveola de la isla de Grenada en las Antillas presentan un polimorfismo en el color del plumaje, en el cual existen individuos de color amarillo y negro e individuos totalmente negros. Al sudeste de la isla existe una clina de tipos de plumaje en la cual la frecuencia de individuos negros se incrementa con la distancia desde el extremo sudoccidental de la isla. Describimos la forma y posición actual de esta clina en comparación a cuando fue mapeada hace veinte años, y mostramos que no ha ocurrido un desplazamiento neto durante este período. Sugerimos que los movimientos anteriores de esta clina podrían estar relacionados con variaciones a largo plazo de la precipitación.
The two Bananaquit color morphs and the morph-ratio cline on Grenada have been described in detail by Wunderle (1981, 1983): a black and yellow morph (“yellow”) ubiquitous in the Caribbean and much of Latin America, and an all-black (“black”) morph found only on Grenada, St Vincent, and two small islands off the Venezuelan coast (Theron et al. 2001). This system is particularly useful for the study of avian clines because the color difference between the morphs is encoded by two alleles at a single locus, with black dominant to yellow (Theron et al. 2001); the clines are narrow and therefore easy to study; they are replicated; and there are some good historical data on their position.
Clines have attracted substantial theoretical study, and can be categorized as equilibrium or dynamic. Equilibrium clines are maintained by a balance between selection and dispersal, and show no long-term directional movement (Endler 1977). Clines in which a wave of new alleles moves through a population are designated as dynamic (Fisher 1937). The cline described in this note is known to have undergone previous movements, consistent with a dynamic spread of the yellow allele into previously all-black populations (Wunderle 1981, 1983). The aim of this study was to determine whether this spread has continued, or whether some other interpretation of the previous movements, consistent with an equilibrium cline, is possible.
Methods
We carried out fieldwork in the cline during September 2002. In common with Wunderle (1983), we censused different sites by walking slowly and observing Bananaquits along trails and roads in southwestern Grenada (12°01′N, 61°45′W; Fig. 1). We also mist netted and banded Bananaquits at seven sites across the cline. At each of these sites, a total of approximately 40 m of five-shelf net in four or five lengths was set from about 07:30 until about 15:30 for 2–3 days. Two tape recorders placed below the nets were used to play back short sections of Bananaquit song, and were moved regularly between the sections of net. Observational and netting data were collected, where possible, at Wunderle's (1983) sites, but, where there had been changes in access or habitat, we substituted sites at equivalent points in the cline. Data were used only from sites where at least 10 Bananaquits were caught or seen. The straight-line distance from Point Salines lighthouse to each site was measured from a 1:25 000 map (Ordnance Survey International 1988). We used data from Wunderle (1983) to compare the position of the cline in 1981 and 2002. Following Wunderle (1981), our sites can be considered as belonging mainly to two transects following the northern and southern sides of the southwestern peninsula. All data were collected between 30 August and 28 September 2002, and sites were visited in haphazard order with respect to their likely morph ratio.
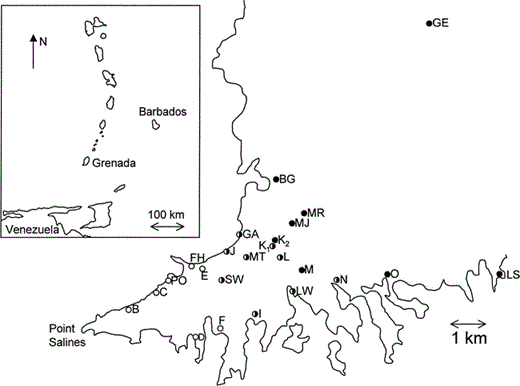
The location of Banaquit sampling sites on the Point Salines peninsula in southwestern Grenada, with (inset) the Lesser Antilles showing the position of Grenada. Circles indicate approximate proportion of black-morph birds at a site in 2002: unfilled: = <20% black morph, half-filled = 20–80%; filled = >80%. Table 1 contains exact values for each site. We split Wunderle's (1983) site K into K1 (Golf Course South) and K2 (Golf Course North) owing to differences in morph ratio in the two areas. Single-letter codes are sites originally sampled by Wunderle (1983); see Table 1 for full names of all sites
The percentage of black-morph Bananaquits at 24 sites in Grenada, September 2002. Single-letter site codes were originally used by Wunderle (1983). Two-letter site codes are other sites surveyed by us. Where we gathered netting and observational data at the same site, we included only the netting data in this table. Sites are listed in order of distance from Point Salines (Fig. 1)
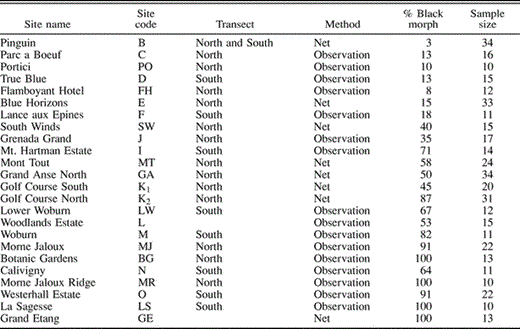
The percentage of black-morph Bananaquits at 24 sites in Grenada, September 2002. Single-letter site codes were originally used by Wunderle (1983). Two-letter site codes are other sites surveyed by us. Where we gathered netting and observational data at the same site, we included only the netting data in this table. Sites are listed in order of distance from Point Salines (Fig. 1)
Statistical Analyses
The data were analyzed with logistic regression models using PROC GENMOD in SAS (SAS Institute Inc. 1999). In these models the response variable had the number of black birds caught or seen at a site as the numerator, and the total number of bananaquits caught or seen as the denominator.
Results
Table 1 shows the sample sizes and percentages of black morphs recorded by us at different sites, excluding observational data that was gathered at the same locations as netting data. We were able to record data at or close to 12 of the 15 sites recorded by Wunderle (1983) in 1981. We also recorded data at another 11 sites. At one of Wunderle's sites (“Golf Course”) we netted in two areas about 250 m apart, in scrub woodland on the north and south sides of the golf course itself. Having done so we found that the morph ratios in the two places were significantly different (χ21 = 10.4, P = 0.001), and so we considered these as two separate sites. We do not know how they compare to the exact location of Wunderle's golf course site, although when we combine our data from the two sites the ratio is 71% black, very similar to that obtained by Wunderle (67% black).
There was no change in the position or slope of the cline between 1981 and 2002 (Fig. 2). In a logistic regression model, distance from Point Salines was highly significant (χ21 = 365.0, P < 0.001), there was no effect of year (χ21 = 0.3, P > 0.5) or the interaction between distance and year (χ21 = 0.3, P > 0.5). In contrast to Wunderle (1981), we found a small but significant difference in ratios between netted samples and observations (for the difference in intercepts for models of netted and observed ratios χ21 = 5.3, P = 0.02). This difference was consistent with a slight over-recording of the rarer morph at the extremes of the cline in observational samples. The cline was significantly less steep along the south coast than along the north coast of the peninsula (χ21 = 4.5, P < 0.05), such that the widths of the cline (between ratios 0.2 and 0.8) were 3.4 km and 3.2 km respectively. Our estimate of the width of the cline through the southern transect was 0.9 km less than Wunderle's (1983) estimate, but the difference was not significant (χ21 = 0.8, P = 0.39). The width of the northern cline is very similar to Wunderle's estimate of 3.0 km.
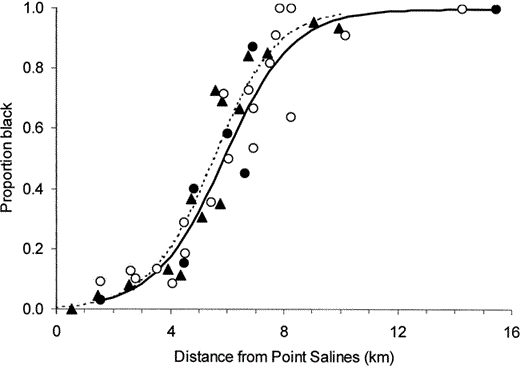
The relationship between the percentage of black-morph Bananaquits at sampling sites and their distance from Point Salines lighthouse, Grenada. Triangles and the dashed line indicate Wunderle's 1981 data (Wunderle 1983). Circles and the continuous line correspond to our 2002 data. Filled circles are data from netting, unfilled circles are observational data
The percentage of black morphs at different sites is strongly related to rainfall (Wunderle 1981), with the black morph more common in wetter areas farther from Point Salines. Therefore, we explored the possibility that previous variation in the position of the cline might have been related to temporal variation in rainfall. Available rainfall data for Grenada do not cover a sufficiently long period, so instead we examined data for neighbouring parts of the Caribbean. The data are from the global precipitation dataset (gu23wld0098.dat, Version 1.0), constructed and supplied by Dr. Mike Hulme at the Climatic Research Unit, University of East Anglia, UK (Hulme et al. 1998, Hulme 2000). Figure 3 shows variation in five-year mean rainfall for Barbados from 1900 to 1996. Barbados is 270 km from Grenada to the northeast, and is the nearest West Indian island with available long-term rainfall data. We also examined data for coastal Venezuela, approximately the same distance from Grenada as Barbados, in the opposite direction. Annual rainfall in the two areas is strongly correlated for the period (r = 0.65, P < 0.001; Sokal and Rohlf 1981), and we are therefore confident that these data are a reasonable indicator of long-term rainfall patterns in Grenada. The rainfall records from both Barbados and Venezuela indicate that this part of the Caribbean was exceptionally dry in the late 1920s, the early 1960s, and the late 1970s (Fig. 3).
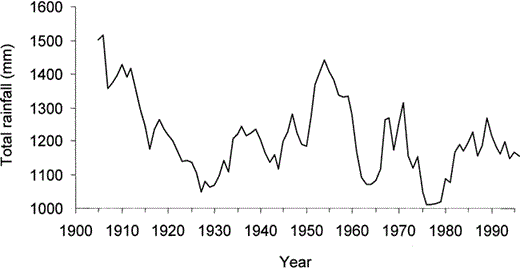
Average annual rainfall in the previous five years against year for Barbados, 1905 to 1996
Discussion
The movement of the cline recorded between 1978 and 1981 (Wunderle 1983) has not continued. There are at least two possible explanations for this. First, the cline may have become trapped in a region of low density or at a habitat barrier (Barton and Hewitt 1985). However Bananaquits have broad habitat preferences (as shown by their ubiquitous distribution, from beaches to mountain rainforest) and there are no large habitat discontinuities or obvious barriers to dispersal in this part of Grenada. Nor do there appear to be any obvious large-scale variations in Bananaquit density, although it is impossible to be certain about this.
Second, the cline may be in equilibrium, maintained by a balance between selection and dispersal (Endler 1977). In the late nineteenth century there were very few yellow birds in Grenada, and these were confined to the extreme southwest (Wunderle 1981). However, the distribution of yellow birds expanded during the early part of the twentieth century, equivalent to a northeastern movement of the cline (Wunderle 1981). This pattern, and Wunderle's (1983) observation of another significant movement of the cline to the northeast between 1978 and 1981, argues against it being in equilibrium. Nevertheless, the cline could still be in equilibrium, with movements linked to changes in a selection regime associated with environmental conditions.
It is clear that the relative proportions of the two morphs at different sites is strongly related to spatial variation in rainfall, with yellow birds occurring in only the driest areas (Wunderle 1981). This suggests the possibility that the observed movements of the cline have been related to temporal variation in rainfall. In this case, we would expect drier conditions to favor the spread of the yellow allele. Observed movements of the cline are consistent with rainfall data in this respect since the large spread in yellow birds that was recorded in 1929 (Wunderle 1981) came immediately after a very dry period, and the similar spread of yellow alleles between 1978 and 1981 followed the very dry period that this part of the Caribbean experienced in the late 1970s. Although the movement of the cline at this time did not exactly match the period of low rainfall, we believe that it is likely that there would be lag in the response of the Bananaquit population to changes in rainfall, which could amount to several breeding seasons.
It has previously been shown that Bananaquits on another Caribbean island, Puerto Rico, suffered a large population decline during a drought there in the mid-1970s, and that there was a lag in the response to rainfall (Faaborg et al. 1984). The size of the Bananaquit population was most closely related to rainfall two years previously. This provides strong support for our suggestion that drought could have perturbed the Bananaquit population in Grenada. If black birds are more strongly affected by drought than yellow birds, as is suggested by their relative rarity in the driest areas of Grenada, then it seems likely that drought could result in movement of the cline. If movement in the cline is caused by variation in rainfall acting with a lag, then the cline should have moved to the southwest during the late 1950s, following a wet period in the mid-1950s. We have been unable to find data to test this prediction.
In summary, the Bananaquit morph-ratio cline has not experienced any net movement in 20 years, which is approximately 20 generations. This suggests that the cline is in equilibrium. Previous movements of the cline may have been the result of long-term variation in rainfall.
Acknowledgments
We thank the Ministry of Agriculture, Forestry, Lands and Fisheries, Grenada, for permission to carry out the work. We also thank C. Macpherson, M. Phillip, D. Ramsier, and M. Sousa for assistance in Grenada; Blue Horizons Hotel and Mr. Ross for permission to mist net on their land, and B. Hatchwell and J. Wunderle for comments on a previous version of the manuscript. The work was funded by the British Ecological Society.
Literature Cited

{kind=link}
{kind=link}
{kind=link}