





Abstract
Silvicultural practices following clearcutting in boreal forest may encourage the creation of monospecific, single-aged stands having less vegetation heterogeneity and diversity than original stands. We conducted point counts in central Saskatchewan, Canada, 1993–1995, in pure and mixedwood stands dominated by black spruce (Picea mariana), jackpine (Pinus banksiana), trembling aspen (Populus tremuloides), or white spruce (Picea glauca). Mixedwood stands supported more individuals and more species than pure stands. Higher abundance in mixedwood stands relative to pure stands was consistent among nesting guilds and migration strategies. Rarefaction revealed similar patterns, although pure trembling aspen stands were predicted to support more species than aspen-dominated mixedwood stands. Increased avian diversity in mixedwood stands was not solely the result of the mixing of bird species associated with coniferous or deciduous forest types. Chipping Sparrow (Spizella passerina), Pine Siskin (Carduelis pinus), White-winged Crossbill (Loxia leucoptera), Red-breasted Nuthatch (Sitta canadensis), Swainson's Thrush (Catharus ustulatus), and Tennessee Warbler (Vermivora peregrina) were more abundant in mixedwood stands than pure stands. Black-throated Green Warbler (Dendroica virens), Magnolia Warbler (D. magnolia), and Blackburnian Warbler (D. fusca) were abundant in stands dominated by white spruce but were absent from jackpine or black spruce. Other species such as American Redstart (Setophaga ruticilla) and Chestnut-sided Warbler (D. pensylvanica) relied exclusively on pure trembling aspen, particularly stands with dense shrub cover. Several bird species in the boreal forest will be adversely affected by forestry practices that target mature to old aspen and white spruce mixedwoods and promote reduction in mixedwood compositions of regenerating stands.
Introduction
Breeding bird communities associated with southern regions of the North American boreal forest are among the richest and most diverse on the continent (Kirk et al. 1996, Niemi et al. 1998). The majority of species occurring here are Neotropical and short-distance migrant songbirds (Smith 1993). Extensive tracts of contiguous boreal forest undoubtedly represent source habitat for many species and may produce a large portion of the continental breeding population each year. For other species, such as the Tennessee Warbler (Vermivora peregrina), Magnolia Warbler (Dendroica magnolia), Cape May Warbler (D. tigrina), Palm Warbler (D. palmaram), Bay-breasted Warbler (D. castanea), Connecticut Warbler (Oporornis agilis), and Philadelphia Vireo (Vireo philadelphicus), the Canadian boreal forest represents most, if not all, of their breeding range in North America.
An important characteristic of the boreal forest of central and western North America is the mosaic nature of forest stands at a landscape scale. The heterogeneity in stand age and structure results from a fire-dominated disturbance regime, which contributes to the diversity of forest bird communities (Niemi et al. 1998). Previous studies in the boreal forest have concentrated on describing how avian communities change with recognized successional stages. In western Canada, most of this work has emphasized the influence of stand age within deciduous systems (Westworth and Telfer 1993, Schieck et al. 1995). Other studies have described avian communities in conifer-dominated boreal forests, primarily in eastern North America (Erskine 1977, Thompson et al. 1999). Although there appears to be a trend toward higher avian diversity in deciduous stands, this may be less characteristic of more northern forests and may depend on the dominant species of conifer (Kirk et al. 1996, Willson and Comet 1996b). How mixed conifer-deciduous forests influence avian communities is poorly understood (Enoksson 1995). As mixedwood stands often have attributes of deciduous and coniferous stands, avian diversity in such stands might be expected to be higher than in more homogenous forests. Threshold levels of deciduous trees in conifer stands or conifer trees in deciduous stands required by deciduous-associated or conifer-associated birds are largely unknown (Willson and Comet 1996b, Robichaud and Villard 1999). Moreover, mixedwood stands may have unique vegetative and structural attributes that do not resemble those of pure deciduous or pure conifer stands.
Understanding how boreal-forest bird communities respond to forest composition is crucial to predicting how anthropogenic change in the boreal forest will impact a significant component of North America's breeding birds. In western and central Canada, the mixedwood composition of stands that regenerate following harvesting are expected to differ from natural-origin forests due to age truncation and silvicultural practices (Spencer 1993, Graham and Jain 1998). Although details of future mixedwood management are uncertain, the conversion of mixedwoods in favor of single-species management represents a conservation issue, particularly because the maintenance of pure stands may be more commercially viable (Smith and Crook 1996, Grover and Greenway 1999). To determine how breeding bird communities might be affected by single-species forest management, we examined how avian species richness, abundance, and species composition differed in naturally occurring stands of pure and mixed black spruce (Picea mariana), jackpine (Pinus banksiana), trembling aspen (Populus tremuloides), and white spruce (Picea glauca).
Methods
Study Area and Stand Selection
The study was conducted within the southern boreal ecoregion of Saskatchewan, Canada (Kabzems et al. 1986). Successional patterns are complex and related to soil type, moisture, and nutrient regimes (Johnson 1992). Three coniferous species, black spruce, jackpine, and white spruce, and a single deciduous species, trembling aspen, dominate forest stands in this region. The Saskatchewan Forest Inventory system defines pure stands as any stand type where more than 75% of the canopy is comprised of a single tree species (Kabzems et al. 1986). In mixedwood stands, more than 25% of the canopy consists of two or more tree species. All stands examined in this study were grouped into one of eight stand types according to the classification given on Saskatchewan Forest Inventory maps.
Stand selection was based on provincial forest inventory maps and cover maps for Prince Albert National Park. Stands were located in contiguous forest and were bordered by an access road or trail. Four point-count stations were located within each stand. Stations were at least 250 m apart and 100 m from an edge, including edges of other stand types. Stands ranged in size from 12 to several thousand hectares. Surveys were conducted in 11 stands of black spruce dominated forests (6 in pure, 5 in mixedwood), 30 in jackpine (21 in pure, 9 in mixedwood), 54 in aspen (27 in pure, 27 in mixedwood), and 30 in white spruce (13 in pure, 17 in mixedwood). All stands had regenerated following fire, 60 to 110 years ago. Stand age was derived from Saskatchewan Forest Inventory maps.
The majority of stands (n = 64) were surveyed in and around the Prince Albert Model Forest, 70 km north of Prince Albert (53°50′N, 105°50′W) during the summers of 1993 and 1994. An additional 16 stands were surveyed near Big River (53°50′N, 107°01′W) in 1993, 16 near La Ronge in 1995, and near Meadow Lake (54°08′N, 108°26′W) in 1993. All stands were separated by at least 7 km, and each stand was surveyed in only one year.
Bird Survey Techniques
Early morning point counts (04:00–09:00) followed the Indice Ponctual D'Abondance technique of Blondel et al. (1970). At each point-count station, all birds heard or seen during a 10-min count were recorded. Counts were of unlimited distance, subject to the constraint that only birds estimated to be within the stand were recorded. Observers recorded the approximate location of singing birds on maps and excluded birds detected previously. Surveys were performed by five experienced observers who were tested in the lab and field to ensure similar levels of expertise. Point counts were conducted twice at each station, in early and late June. To reduce the influence of observer bias, observers surveyed each station only once, with the order of visits chosen randomly.
All raptors and species that nest in wetland habitats were excluded from analysis. Rare species detected in less than five stands were not analyzed for stand preferences, although all species were used when calculating species richness and total number of individuals. Based on the literature (Freemark and Merriam 1986, Schieck et al. 1995) and our own observations, species were categorized by migratory status (resident, short-distance migrants, Neotropical migrants, and irruptive species) and preferred nesting location (ground, shrub, canopy, and cavity).
Vegetation Sampling
To validate the forest inventory classification for each stand, we visually estimated vegetation attributes while at each station. Canopy, subcanopy, and shrub closure within a 25-m radius of each point-count station were classified as either open (25% closure), moderately open (25 to 50%), moderately closed (50 to 75%), or closed (>75%; Kabzems et al. 1986). The percentage of the canopy, subcanopy, and shrub layer comprised of different species was estimated to the nearest 10%. The average height of the canopy was visually estimated to the nearest meter and the shrub layer to the nearest 50 cm. In four 1-m2 quadrats placed 10 m apart, and in the four cardinal directions centered at the point-count station, we visually estimated the percentage cover to the nearest 10% of low shrubs, lichen, moss, grass, and herbs. Shrubs were defined as any woody plant 1–5 m tall. Subcanopy trees were greater than 5 m tall and were at least 20% shorter than trees comprising the canopy. Using the Shannon index, we estimated canopy, subcanopy, shrub, and ground cover diversity for each station. Each station within a stand was used to calculate an average vegetation score for that stand, which was then used in subsequent analyses.
Statistical Analyses
To determine whether vegetation structure differed among stand types, we used a series of randomization tests based on a two-factor analysis of variance (Manly 1991). In these analyses, the dominant tree species (black spruce, jackpine, trembling aspen, and white spruce) and the degree of stand heterogeneity (pure versus mixedwood) were the independent factors. The significance of the main and interactive effects was determined by randomly shuffling the original data 5,000 times. The number of times the randomly shuffled data had a residual sums of squares (SS) greater than the observed data was used to determine the approximate probability value (Manly 1991). When randomization tests indicated significant main or interactive effects, a randomization test based on the least significant difference (LSD) procedure was used to determine which groups within factors were significantly different. The strength of randomization testing is that the underlying distribution does not need to follow any particular statistical distribution, nor do the data have to fit the assumption of homoscedasticity (Manly 1991).
Species richness for each station was estimated using the program SPECRICH2 (Nichols et al. 1998). SPECRICH2 is an inference method that uses a capture–recapture methodology to estimate the number of species present, correcting for sampling biases. The benefit of this methodology is that it accounts for the problem of differential detectability of species that can influence estimates of species richness when comparing bird communities among different vegetation types (Schieck 1997). The two visits to a station were used as replicate samples of the avian community in SPECRICH2 calculations. Station estimates of species richness were averaged to determine species richness per stand, a value used in all further analyses.
To determine whether species richness, total abundance, abundance of individuals in each nesting guild, and abundance of each migratory group were different among stand types, we used two-factor randomization tests. Abundance was based on the maximum number of individuals of each species detected per station. Abundance per station was pooled for all stations within a stand to determine the abundance of individuals at the stand level.
As sampling effort was unequal among stand types, rarefaction was used to estimate how species richness and total abundance were influenced by sample size. Rarefaction estimates the number of species and number of individuals expected from a given sample of counts based on multiple random sampling (James and Rathbun 1981). Rarefaction estimates were calculated using the computer program EstimateS 5 (Colwell 1997). In this analysis, the maximum number of individuals detected per station was used as our independent measure of the bird community. Rarefaction estimates of species richness were compared among tree species and stand heterogeneity by taking the mean species richness and standard deviation per 25 point-count stations to calculate a series of single-factor ANOVA tests (Gjerde and Sœtersdal 1997). Similar analyses were done with total abundance.
Two-factor randomization tests also were used to determine whether the average abundance of individual species was higher than expected if birds were randomly distributed among stand types. Again, the dominant tree species and the degree of stand heterogeneity were the independent factors. Those species that showed significant (P < 0.10) main or interactive effects among stand types were subjected to two-way indicator species classification (TWINSPAN) to determine which species had the most similar habitat requirements (Gauch 1982). TWINSPAN is a divisive polythetic classification that successively divides stands and species into smaller clusters. Stand clusters are based on the similarity of the avian community; stands clustered together are more likely to contain the same species. Species are clustered based on the similarity of their habitat preferences, and so species clustered together are more likely to be detected in the same stand type. TWINSPAN is based on a reciprocal averaging algorithm (Gauch 1982), so the order of species and sites along the table axis is important. Species next to one another have the greatest similarity in habitat requirements, and sites next to one another have the greatest avian community similarity. The abundance of each species was divided by the total abundance of birds in each stand (i.e., converted to a percentage) before analysis with TWINSPAN, so that default cut levels could be employed.
We used a multiple response permutation procedure (MRPP) to determine whether the divisions of the stands made at each step of the TWINSPAN analysis were significant. MRPP is a nonparametric procedure for testing the hypothesis of no difference between two or more groups of multivariate data (Zimmerman et al. 1985). When the significant TWINSPAN stand clusters had been determined, a single-factor randomization procedure was used to determine which vegetation characteristics and which avian species were significantly different between stand clusters. Stand clusters were created using only the bird data set and were not constrained by the vegetation data set. TWINSPAN and MRPP tests were done using the computer program PC-Ord (McCune and Mefford 1997). All data are reported as x̄ ± SD unless otherwise stated.
Results
Vegetation Structure
Jackpine and trembling aspen stands had mean ages of 87 to 90 years and were younger than the black spruce and white spruce (100 ± 21 years) stands that averaged 111 and 100 years, respectively. The canopy was taller in white spruce (19 ± 3 m) and trembling aspen (20 ± 4 m) than in jackpine (13 ± 3 m) or black spruce (12 ± 2 m) stands (P < 0.001). Canopy closure differed among stands dominated by different tree species (P < 0.05), although no LSD tests were significant. Mixed and pure stands were of similar age, canopy height, and canopy closure. However, mixedwood stands had significantly higher canopy diversity (0.76 ± 0.22) than pure stands (0.37 ± 0.26, P < 0.001).
Subcanopy trees were taller in white spruce (10 ± 4 m) and trembling aspen stands (12 ± 4 m) than in jackpine (7 ± 1 m) and black spruce (7 ± 1 m) stands (P < 0.001). Subcanopy diversity was lower in black spruce (0.07 ± 0.11) stands than jackpine (0.31 ± 0.33), trembling aspen (0.49 ± 0.37), or white spruce (0.41 ± 0.34, P = 0.04). Mixedwood stands had greater subcanopy closure (47 ± 27%, P = 0.05) and subcanopy diversity (0.32 ± 0.31, P < 0.05) than pure stands (32 ± 25% and 0.07 ± 0.11, respectively). However, for subcanopy closure, the interaction between tree species and stand heterogeneity was significant (P < 0.05), as pure black spruce stands had a denser subcanopy than black spruce mixedwoods.
Shrub height and shrub closure were higher in stands dominated by trembling aspen than in the other stand types (P < 0.001). Shrub diversity was higher in trembling aspen (0.72 ± 0.59) and white spruce (0.73 ± 0.64) than jackpine (0.07 ± 0.14) and black spruce (0.03 ± 0.07, P < 0.001). Ground cover diversity was higher in trembling aspen (1.34 ± 0.19) and white spruce (1.39 ± 0.15) than jackpine (1.16 ± 0.20) or black spruce (1.0 ± 0.32, P < 0.001). There was no difference in shrub height (P > 0.20), shrub closure (P > 0.50), shrub diversity (P > 0.50), or ground cover diversity (P > 0.50) between pure and mixedwood stands.
Avian Community
We detected 13,060 individuals of 87 bird species. Of these, 30 species were rare (detected at < 5 sites) and not analyzed statistically. Of the 57 bird species analyzed to establish a preference for a particular stand type, 32 showed significant differences between stands dominated by different tree species (detailed information available from authors on request). Controlling for tree species, Chipping Sparrow (Spizella passerina), Pine Siskin (Carduelis pinus), White-winged Crossbill (Loxia leucoptera), Red-breasted Nuthatch (Sitta canadensis), Swainson's Thrush (Catharus ustulatus), and Tennessee Warbler were more abundant in mixedwood stands than pure stands. The interaction between stand heterogeneity and dominant tree species was significant for Red-eyed Vireo (Vireo olivaceus) and Ovenbird (Seiurus aurocapillus). Both of these species were most abundant in pure aspen stands, but also were common in mixedwood stands dominated by coniferous tree species.
Species Richness
At the stand level, species richness was higher in stands dominated by white spruce (21.6 ± 1.1) and trembling aspen (20.8 ± 0.8) than in jackpine (14.7 ± 1.2) or black spruce stands (13.8 ± 2.2, P < 0.001). Controlling for the dominant tree species, mixedwood stands supported significantly more species than pure stands (20.0 ± 1.1 vs. 15.4 ± 0.9, P = 0.01). This pattern was consistent among the different tree species, as the interaction with stand heterogeneity was not significant (Table 1).
Mean (± SD) species richness and total abundance of the different migratory and nesting guilds at the stand level. Also provided are the results of randomization tests comparing stands dominated by different tree species (tree), level of forest heterogeneity (hetero.), and the interaction between heterogeneity and tree species (T*H)
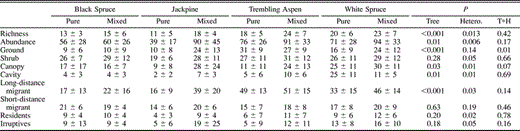
Mean (± SD) species richness and total abundance of the different migratory and nesting guilds at the stand level. Also provided are the results of randomization tests comparing stands dominated by different tree species (tree), level of forest heterogeneity (hetero.), and the interaction between heterogeneity and tree species (T*H)
Species richness, as estimated by rarefaction (hereafter landscape richness), differed among the dominant tree species when pure and mixedwood stands were pooled (F3,96 = 133.9, P < 0.001). Species richness at the landscape scale also was different among tree species, and was highest in trembling aspen (53.0 ± 3.1), followed by white spruce (46.7 ± 3.2), jackpine (40.0 ± 3.2), and black spruce (37.7 ± 2.5). We could not detect a difference in landscape richness between pure and mixedwood stands (51.8 ± 4.0 vs. 49.9 ± 3.2, F1,48 = 3.4, P > 0.50). However, the interaction between dominant tree species and stand heterogeneity was significant (F7,192 = 138.7, P < 0.001). Pure and mixedwood stands of trembling aspen had similar species richness at the landscape scale (P > 0.50). LSD tests indicated mixedwood stands of black spruce, jackpine, and white spruce had more species than pure stands (Fig. 1a, P < 0.01 in all cases).
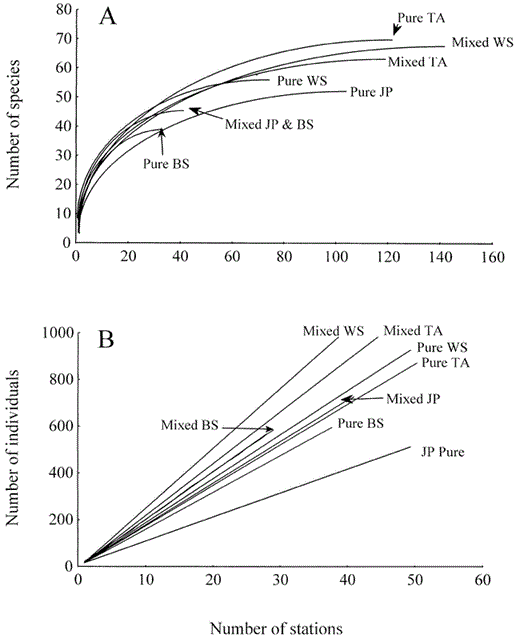
Rarefaction estimates showing: (A) mean species richness and (B) mean abundance of all birds detected at various numbers of point count-stations in each stand type
The number of individuals detected per stand for all species combined (abundance) was higher in white spruce (84.1 ± 32.7) and trembling aspen (83.5 ± 30.5), than in black spruce (57.1 ± 26.0) and jackpine (53.8 ± 35.6, P = 0.01). Mixedwood stands had higher total abundance than pure stands (89.8 ± 34.6 vs. 61.6 ± 28.9, P < 0.01). This pattern was consistent among tree species, as the interaction was not significant (Table 1).
Based on rarefaction estimates, landscape-level abundance was highest in white spruce (568 ± 47), followed by trembling aspen (505 ± 40), black spruce (451 ± 37), and jackpine (353 ± 51, F3,96 = 106.6, P < 0.001). Mixedwood stands had higher abundance than pure stands (571 ± 51 vs. 393 ± 37, F1,48 = 2,083, P < 0.001). However, the interaction between dominant tree species and stand heterogeneity was significant (F7,192 = 872.2, P < 0.001). Higher abundance in mixedwood stands was consistent for all tree species except trembling aspen which had similar total abundance in mixedwood and pure stands (P > 0.20, Fig. 1b).
Nesting guilds
The abundance of cavity-nesting birds was higher in white spruce and trembling aspen than jackpine or black spruce stands (P = 0.01). Canopy-nesting birds were more abundant in white spruce than in trembling aspen, black spruce, or jackpine (P < 0.05). The abundance of shrub-nesting birds did not differ among dominant tree species (P > 0.20). Ground-nesting birds were more abundant in trembling aspen stands than in white spruce, black spruce, or jackpine (P < 0.001). Mixedwood stands had more shrub-, canopy-, and cavity-nesting birds than pure stands, and this pattern was consistent among tree species as none of the interactions were significant (Table 1). The interaction between stand heterogeneity and tree species was significant for ground-nesting birds (P = 0.01). Jackpine-dominated mixedwoods had more ground-nesting birds than pure jackpine stands (P < 0.01). In contrast, all other tree species had similar abundance of ground-nesting species in pure and mixedwood stands (Table 1).
Migratory status
Long-distance migrants, residents, and irruptive species had higher abundance in mixedwood stands than pure stands (Table 1, P < 0.05, in all cases). These patterns were consistent among tree species, as none of the interactions were significant (Table 1). Short-distance migrants were equally abundant in pure and mixedwood stands (P > 0.5). The abundance of residents (P = 0.2), irruptive species (P > 0.1), and short-distance migrants (P > 0.5) was similar among tree species. Neotropical migrants were significantly more abundant in trembling aspen and white spruce than in black spruce or jackpine (P < 0.001).
Twinspan Clusters
Of the 57 bird species analyzed to establish a preference for a particular stand type, 41 showed a significant main or interactive effect at P = 0.1. TWINSPAN grouped these 41 species into seven distinct species clusters (Fig. 2). These species clusters were: (1) species found mainly in black spruce and jackpine: Dark-eyed Junco (Junco hyemalis), Nashville Warbler (Vermivora ruficapilla), Palm Warbler, Ruby-crowned Kinglet (Regulis calendula), Hermit Thrush (Catharus guttatus), and Gray Jay (Perisoreus canadensis), (2) ubiquitous species found primarily in coniferous forest: Pine Siskin, Chipping Sparrow, Northern Flicker (Colaptes auratus), and Yellow-rumped Warbler (Dendroica coronata), (3) species found mainly in white spruce and mixedwood stands containing white spruce: Bay-breasted Warbler (D. castanea), Blackburnian Warbler (D. fusca), Black-backed Woodpecker (Picoides arcticus), Boreal Chickadee (Poecile hudsonicus), Brown Creeper (Certhia americana), Solitary Vireo (Vireo solitarius), Winter Wren (Troglodytes troglodytes), White-winged Crossbill, and Western Tanager (Piranga ludoviciana), (4) common species that had higher abundance in mixedwood stands: American Crow (Corvus brachyrhynchos), Black-throated Green Warbler (D. virens), Red-breasted Nuthatch, Swainson's Thrush, and Tennessee Warbler, (5) common species most abundant in aspen forests with a coniferous or sparse understory: Black-capped Chickadee (Poecile atricapillus), Hairy Woodpecker (Picoides villosus), Magnolia Warbler, and Ovenbird, (6) species commonly found in pure aspen and aspen-dominated mixedwoods: Rose-breasted Grosbeak (Pheucticus ludovicianus), Red-eyed Vireo, White-throated Sparrow (Zonotrichia albicolis), Alder Flycatcher (Empidonax alnorum), Connecticut Warbler, and Philadelphia Vireo, and (7) species found almost exclusively in pure aspen: Brown-headed Cowbird (Molothrus ater), Least Flycatcher (Empidonax minimus), Mourning Warbler (Oporornis philadelphia), Yellow Warbler (D. petechia), American Redstart (Setophaga ruticilla), Canada Warbler (Wilsonia canadensis), and Chestnut-sided Warbler (D. pensylvanica).

Dendrogram showing results of TWINSPAN analysis. When species are closer together on the axis, they are more likely to be found in the same forest stands
At the fourth division in the TWINSPAN classification, 16 stand clusters are typically created. However, pairwise comparisons using MRPP tests indicated only six of the TWINSPAN splits resulted in clusters with significantly different bird communities. Vegetation variables and avian species that were significantly different among stand clusters created by TWINSPAN are shown in Figure 3.

Vegetation variables and bird species that were significantly different among TWINSPAN stand clusters. For each variable, cells in black are not different (based on LSD randomization test) from the highest value, white are not different from the lowest value, and gray are significantly different from the low and high values
Discussion
Our comparison of pure and mixedwood stands in forests dominated by four different tree species demonstrated that avian communities were richer and hosted a greater number of breeding individuals in mixedwood stands compared to similar-aged pure stands. This trend was especially notable for mixedwood stands of black spruce, jackpine, and white spruce for which the results of rarefaction analyses confirmed those of stand-level analyses. For aspen-dominated mixedwoods, stand data showed the same trends as for the other stand types, but rarefaction analyses ultimately predicted greater cumulative number of species in pure vs. mixed aspen stands after 40 point-counts. The higher densities of ground-nesting Neotropical migrants using pure aspen stands compared to the other stand types may have caused this difference. Higher species richness and abundance in mixedwood stands were likely caused by greater floristic and structural diversity in the canopy and subcanopy of mixedwood stands relative to pure stands (Bersier and Meyer 1994, Kirk et al. 1996).
For birds in temperate regions, there appears to be a close relationship between habitat diversity and species diversity (Rosenzweig 1995). Even small patches of different habitat within uniform stands are often enough to account for the occurrence of a particular species. For example, in their analysis of habitat associations of breeding birds of boreal forest in Alaska, Willson and Comet (1996b) noted that one or two spruce trees in a deciduous stand was enough for Ruby-crowned Kinglet or Hermit Thrush to hold a territory, and that these species would forage in the surrounding deciduous vegetation. Similarly, Robichaud and Villard (1999) found that Black-throated Green Warblers breeding in Alberta, Canada, apparently selected territories in mixedwood forest based on the distributions of coniferous species. However, the increase in species richness we observed in mixedwood stands was not just because species that use deciduous forest were found in mixedwood-conifer stands at lower abundance and vice versa. The Chipping Sparrow, Pine Siskin, White-winged Crossbill, Red-breasted Nuthatch, Swainson's Thrush, and Tennessee Warbler were significantly more abundant in mixedwood stands relative to pure stands.
Mixedwood stands may support more species and more individuals for a variety of reasons. Increased structural diversity in mixedwood stands likely provides a greater array of nesting locations than pure stands. Consequently, a greater diversity of bird species may be able to coexist per unit area in mixedwoods. Structural heterogeneity in mixedwoods may increase nesting success by reducing the ability of predators to find nests due to greater visual clutter around nests (Martin 1993). In our study area, artificial nests in white spruce-dominated mixedwoods were less likely to be destroyed by red squirrels (Tamiasciurus hudsonicus) than nests in pure white-spruce stands, even though the abundance of squirrels was similar between these stand types (Bayne et al. 1997). Food abundance also could be different in mixedwoods than in pure stands. Litter and shrub invertebrates can reach higher densities in deciduous forests than in coniferous forests (Werner 1983, Willson and Comet 1996a). Conversely, the canopies of coniferous forests often have higher densities of larval lepidoptera than deciduous forests (Qiong et al. 1996). With coniferous and deciduous trees together, mixedwoods may provide a more diverse and abundant food supply for birds.
Although stand heterogeneity resulted in increased species richness and abundance, absolute differences among stands were highly dependent on the dominant tree species. Overall, white spruce and trembling aspen stands had higher species richness and abundance than jackpine or black spruce stands. In their review of bird communities in forests across North America, James and Wamer (1982) suggested that coniferous forests with only one or two tree species typically had fewer bird species than deciduous forests of the same age. Willson and Comet (1996b) argued that differences in species richness between deciduous and coniferous forests are less consistent in northern and western regions of North America, but that the contrast between coniferous and deciduous forests decreases with increasing latitude. Our data suggest such generalizations about the level of avian diversity in deciduous and coniferous forests may be too simplistic. In the boreal forest of Saskatchewan, bird communities in trembling aspen forests differed considerably from those in coniferous forests. However, white spruce stands supported a much more diverse bird community than black spruce or jackpine. White spruce stands had greater shrub diversity and larger trees than black spruce or jackpine stands, which likely resulted in a greater volume of foliage. Foliage volume is an important factor influencing insect abundance (Newton and Moss 1977) and nest site availability (Martin 1993).
Differences in total bird abundance among stand types were not consistent among nesting guilds. Cavity-nesting birds were most common in mixedwoods, particularly white spruce and trembling aspen, possibly due to the larger size of the trees in these stands. Canopy-nesting birds were more abundant in white spruce stands, reflecting the higher volume of foliage and greater vertical structure of these trees relative to jackpine and black spruce. Shrub-nesting birds were more abundant in mixedwoods, likely due to greater subcanopy diversity in these stands. Ground-nesting birds were most common in pure and mixed trembling-aspen stands, and this corresponded with greater ground cover diversity in these stands.
Almost all bird species in this study were found in at least two stand types and 43% of species were found in all stand types. This suggests that many boreal forest birds have relatively broad habitat requirements, a pattern consistent with previous studies of northern bird communities (Erskine 1977, Welsh and Lougheed 1996, Willson and Comet 1996b). Many species in Saskatchewan seem to be able to use any conifer-dominated habitat (Yellow-rumped Warbler, Ruby-crowned Kinglet, Chipping Sparrow, Cape May Warbler). However, the majority of species found in coniferous forests were most abundant in pure and mixed white-spruce stands (e.g., Blackburnian Warbler, Bay-breasted Warbler, and Winter Wren). Old white-spruce stands are becoming particularly rare in the landscape due to their economic value as timber (Cumming and Armstrong 1999). Conversely, no species occurred exclusively in jackpine stands and many appeared to avoid this stand type. This may be significant from a conservation perspective because silvicultural programs in many areas of western Canada encourage planting of jackpine in cutover spruce or aspen areas due to the rapid growth of jackpine on most soil types (Kabzems et al. 1986). Although many species in the boreal forest seem to be habitat generalists, species like American Redstart, Canada Warbler, and Chestnut-sided Warbler rely on trembling aspen stands exclusively (Hobson and Bayne 2000).
Previous studies of avian habitat associations in the boreal forest of North America have revealed gradients associated with dry to wet stands or with relative deciduous and coniferous composition, usually related to stand age (Schieck et al. 1995, Kirk et al. 1996, Welsh and Lougheed 1996). Our study is the first to examine community changes associated with mixedwood vs. pure stands in western Canada and reveals important potential consequences of silvicultural manipulations in the boreal forest. Current logging practices in the boreal forest of western and central Canada are targeting mature to old white spruce and aspen forests (Cumming et al. 1994). This trend is alarming in that these forests have higher avian diversity than jackpine or black spruce forests common throughout much of the boreal. In addition, stand treatment and future rotational cutting regimes are expected to result in the replacement of mixedwood stands by uniform stands dominated by one tree species and a simplified understory structure (Welsh 1993, Kirk et al. 1996). White spruce stands often occur as the climax phase of succession in the boreal forest and require more time to develop than currently allowed under standard rotation cycles (Kirk et al. 1996). Our study suggests that many avian species may be adversely affected by these changes. Sustainable management of the boreal forest to maintain bird communities requires that appropriate mixtures of different tree species be provided at both the stand and landscape level. Further studies are required that evaluate the most optimal distribution of stand types and ages, because little information exists on the effects on avian communities of landscape-level changes to natural patterns of forest cover in the boreal forest (Drolet and Desrochers 1999).
Acknowledgments
Funding was provided by the Canadian Wildlife Service, Prince Albert National Park, Prince Albert Model Forest (Project #211), and a federal-Saskatchewan Partnership Agreement in Forestry (Project #CS5006). A Natural Sciences and Engineering Research Council of Canada post-graduate scholarship and a graduate fellowship from the University of Saskatchewan provided personal support to EMB. We thank Prince Albert National Park for allowing us to conduct surveys in the Park and for assisting with logistical support. K. Konze, B. Korol, R. Wapple, and E. Cumming conducted the bird surveys. I. Corns kindly provided reference material on mixedwood management. We thank W. Koenig and two anonymous reviewers for helpful comments on earlier drafts of this manuscript.
Literature Cited

{kind=link}
{kind=link}
{kind=link}