





SUMOylation is a type of post-translational modification (PTM) that covalently links a small ubiquitin-like modifier (SUMO) to a protein target, a process that involves SUMO E1 activases, SUMO E2 conjugases, and SUMO E3 ligases. The best characterized plant SUMO E3 ligase is SIZ1 (SAP AND MIZ1 DOMAIN CONTAINING LIGASE1) (Castro et al., 2018). SUMOylation affects a variety of target proteins, causing conformational changes and creating or blocking interaction interfaces (Wilkinson and Henley, 2010). The roles of SUMOylation in plant growth, development, and response to abiotic and biotic stresses have been extensively studied. Disruption of SIZ1 leads to dwarf plants and reactive oxygen species (ROS) accumulation under abiotic stress conditions (Miura et al., 2010). In contrast, overexpression of SIZ1 decreases ROS accumulation (Miura et al., 2013; Zhang et al., 2018). Furthermore, siz1 and other SUMO pathway mutants also accumulate salicylic acid (SA), a well-known defense hormone. Although the accumulation of SUMO conjugates in plants in response to ROS also has been observed (Miller et al., 2013), the mechanisms involved in the interaction of SUMOylation and ROS are still unclear.
In the current issue of Plant Physiology, Castro et al. (2022) unveiled critical aspects of SIZ1-mediated SUMO–ROS interplay in Arabidopsis (Arabidopsis thaliana). The authors analyzed the SUMO-conjugate profiles in Arabidopsis Columbia-0 (Col-0) plants and siz1-2 mutants treated with either H2O2 or methyl viologen (MV), an herbicide that causes ROS production in plant cells. They found that the MV-induced ROS initiates SUMO conjugation more efficiently than the H2O2-mediated ROS (Castro et al., 2022). SUMO-conjugates induced by H2O2 or MV were drastically reduced in the siz1 mutants compared with those in Col-0, indicating that H2O2- and MV-induced ROS production promotes SUMO conjugation in a SIZ1-dependent manner. In siz1 mutants, the inhibition of growth of photosynthetic tissues by MV was more profound, while root growth inhibition by MV was less severe. These findings suggest that siz1 mutants are more sensitive to oxidative stress.
To investigate the interplay between ROS and SUMOylation, the authors analyzed the H2O2 and O2− levels in Col-0 and siz1-2 plants. They found that siz1-2 mutants had elevated levels of H2O2 and O2−, indicating that SIZ1-dependent SUMOylation is important for ROS homeostasis (Castro et al., 2022). The high level of ROS in siz1-2 mutants may be caused by over-accumulation of SA because higher levels of SA promote ROS production (Herrera-Vásquez et al., 2015). The authors addressed this question by incorporating siz1 in the NahG (transgenic plant expressing salicylate hydroxylase) background, which has substantially reduced SA levels (Lawton et al., 1995). The siz1 NahG plants had decreased levels of H2O2 and O2− compared with the siz1 single mutant, indicating that SA is involved in SUMO–ROS interplay.
The authors further discussed SUMO–ROS–SA interactions. Overproduction of SA could generate H2O2 and O2− in siz1-2 mutant plants and cause damage to plant cells. In response to oxidative stress, SUMO-conjugate levels increase, primarily in the repression complexes involved in histone deacetylase recruitment (Miller et al., 2013). The authors suggested that SIZ1-dependent SUMOylation of histone deacetylase HDA19 contributes to the repression of SA-biosynthesis genes (Figure 1). Hence, SIZ1 serves as an important regulator in SA-dependent ROS homeostasis.
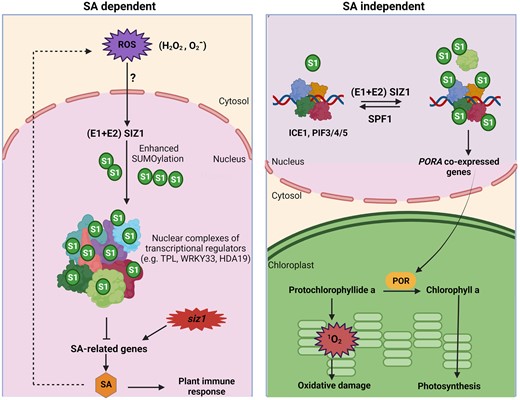
A model for regulating SUMO–ROS interplay by SIZ1 in a SA-dependent and -independent manner. Left panel, reactive oxygen spices (ROS) trigger SUM1/2 (S1) conjugates deposition, in a SUMOylation process under the control of SUMO E1 activases (E1), SUMO E2 conjugases (E2), and SUMO E3 ligase SIZ1. The nuclear components, including several transcriptional regulators, are SUMOylated, causing repression of salicylic acid (SA) associated genes. Mutation in SIZ1 causes an upregulation of SA-related genes, some of which are involved in SA biosynthesis. Accumulation of SA leads to overproduction of H2O2 and . Right panel, SIZ1 regulates singlet oxygen levels in a SA-independent pathway. SUMOylation of the bHLH transcription factors such as ICE1 and PIFs modulates the expression of several co-expressed PORA genes and the de-SUMOylating protease SUMO PROTEASE RELATED TO FERTILITY1 (SPF1) negatively controls PORA genes. The protochlorophyllide oxidoreductase (POR) enzymes are needed to convert PChlide a into chlorophyll a, preventing oxidative damage to the cell. The figure was adapted from Castro et al. (2022) and created with BioRender.com.
In siz1-2 mutants, concentrations of the chlorophyll precursor protochlorophyllide (PChlide), which induces singlet oxygen (1O2) production, were also elevated (Figure 1). The authors proposed that SIZ1 maintains the 1O2 level in the SA-independent pathway by SUMOylating bHLH transcription factors such as PHYTOCHROME INTERACTING FACTORS (PIFs) and INDUCER OF CBF EXPRESSION1 (ICE1) (Figure 1). These SUMOylated transcription factors promote the expression of many PROTOCHLOROPHYLLIDE OXIDOREDUCTASE A (PORA) co-expressed genes and SUMO PROTEASE RELATED TO FERTILITY1 (SPF1, encoding a SUMO protease), which negatively controls PORA genes. Hence, SUMO conjugation of PIFs and ICE1 transcription factors and deconjugation of SPF1 play a critical role in the control of pigment content and 1O2 homeostasis (Figure 1).
Plants respond to multiple environmental stimuli through the ubiquitin-like modifying peptide SUMO. These external stresses lead to the production of ROS. The SUMO-conjugates rapidly accumulate in response to an external oxidative stimulus. ROS and SUMOylation are likely connected at a molecular level. Work by Castro et al. (2022) illustrates a critical overview of ROS regulation by SIZ1-mediated SUMOylation in an SA-dependent/-independent manner. ROS activates SIZ1, which enhances the SUMO-conjugation (Figure 1). Histone deacetylases, such as HAD19, are conserved SUMO targets interacting with TOPLESS (TPL) proteins, and SUMOylation of HAD19 via SIZ1 (Niu et al., 2019) could repress the SA-related genes (Figure 1). The authors also highlighted how SIZ1 negatively regulates PChlide levels and prevents the accumulation of 1O2. Moreover, transcriptomics data demonstrated an association between 1O2 signaling and SIZ1. The present study extensively discusses the crosstalk between ROS and SUMOylation. However, the genetic mechanism directly mediating ROS-induced SUMOylation is still unknown.
Conflict of interest statement. None declared.

{kind=link}