





Chlamydia psittaci is a zoonotic pathogen associated primarily with avian chlamydiosis also referred as psittacosis. Human psittacosis can lead to severe cases of respiratory disease. The mule duck is one of the main bird hybrids associated with human cases of psittacosis in France. In order to better understand the epidemiology of avian chlamydiosis, monitoring studies were performed in both breeder flocks and mule duck flocks. Surveys conducted in one professional duck bredding organization revealed little shedding in breeder flocks, whereas heavy but asymptomatic C. psittaci shedding was observed in most of the mule duck flocks, mostly when birds were reared in open range conditions on farms. Human cases of psittacosis linked to duck breeder flocks and their progeny led to detection of heavy shedders in all the suspected flocks despite no birds showing clinical signs. Offspring of one of the infected female flocks was analyzed and also proved to be infected by C. psittaci. Field studies suggest that C. psittaci infections in duck farms involve horizontal and probably vertical transmission but that the environment also plays an important role in maintaining infection on farms. In the light of the widespread occurrence of C. psittaci on duck farms, it has become urgent to clearly identify sources of contamination in order to take appropriate field management measures to minimize worker exposure.
INTRODUCTION
Chlamydia psittaci is a zoonotic pathogen associated primarily with avian chlamydiosis also referred as psittacosis. Human psittacosis can lead to severe cases of respiratory disease. It has been reported that more than 467 bird species may be affected by chlamydial infection (Kaleta and Taday 2003). In birds, the infection is usually systemic and can be fatal. The severity of clinical signs varies, depending on the bird species, age and C. psittaci strain involved (Andersen 1997). Genotype-specific sites of C. psittaci are located in variable domains of outer membrane protein A (ompA). Avian strains of C. psittaci are currently divided into 13 ompA genotypes designated A to F, E/B, 1V, 6N, MatI16, R54, YP84 and CPX0308 (Sachse et al., 2008). A degree of host specificity can be noted, with genotype A being mostly detected in psittacines, genotypes B and E in pigeons or genotypes C and E/B in ducks (Geens et al., 2005).
Typically, transmission to humans is by the inhalation of infectious aerosols when handling infected birds, carcasses or tissues. The symptoms of human psittacosis are variable, ranging from flu-like symptoms to severe febrile illness associated with atypical pneumonia. Professionals such as veterinarians, bird breeders, handlers or traders have been reported to run higher risks (Huminer et al., 1992; Hinton et al., 1993; Saito et al., 2005). Chlamydia psittaci infection in ducks has been previously reported in Europe, Australia and USA (Chalmers, Farmer and Woolcock 1985; Arzey, Arzey and Reece 1990; Hinton et al., 1993; Léon et al., 2004; Guérin et al., 2006; Laroucau et al., 2009a). In France, about 20 severe human cases are reported each year by the National Reference Centre for Chlamydiae (NRC, Bordeaux, France). These cases are mostly linked to ducks, especially those related to the production of foie gras, a culinary specialty made from fresh livers (Laroucau et al., 2009a and unpublished data). The ducks used for foie gras production are known as mule ducks, being sterile hybrids between male Muscovy ducks (Cairina moschata) and female domestic Pekin ducks (Anas platyrhynchos). Female Pekin ducks are artificially inseminated twice a week. Embryonated eggs are transferred to hatcheries for a 4-week incubation period and only male ducklings are used for foie gras production. After hatching, these males are kept in a closed enclosure on straw for about 3–4 weeks. Afterwards they are kept outdoors, on grass, up to the age of 12 weeks and then transferred to another farm for a 12-day period of force-feeding before slaughter (Fig. 1).
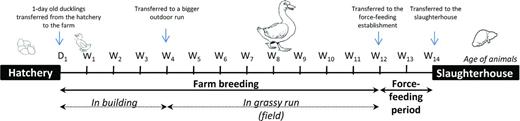
Mule duck production process. One-day old ducklings are transferred to the farm from the hatchery. Up to the age of 4 weeks, animals are kept in a building and generally have access to a small outdoor run. They are then transferred to a larger outdoor run and live outside. At the age of 12 weeks, they are transferred to a force-feeding establishment for 12 days, and finally transferred to the slaughterhouse.
Very little data are available concerning natural C. psittaci infection during the mule duck breeding process. However, it is important to evaluate the chlamydia circulation and identify the critical steps during poultry worker activities in order to reduce human exposure by efficient risk mitigation measures.
The objective of this investigation was to study the occurrence and level of C. psittaci in cloacal swabs—reflecting the shedding of C. psittaci—both in breeder flocks dedicated to mule duck production and in mule duck flocks. Investigations were also conducted on breeder duck farms where human psittacosis cases unexpectedly occurred. A genotype determination method was used to identify circulating C. psittaci strains.
MATERIALS AND METHODS
Avian and human population and sampling
Avian samples
Randomly selected breeder flocks.
In four female Pekin flocks dedicated to the production of mule ducklings, 20 birds were subjected to cloacal swabbing every 6 weeks for a year after the onset of laying Three male Muscovy flocks used to artificially inseminate these females during the survey were also subjected to cloacal swabbing, at the same frequency (Table 1).
Characteristics of in kinetic investigated laying duck flocks (n = 4) dedicated to the mule duck production by artificial insemination. Male flocks used for the semen production were also investigated (n = 3).
| 23S RT-PCR | ||||||||
|---|---|---|---|---|---|---|---|---|
| Flock | Id sampling | Age | Sampling | Sample | C. psittaci | |||
| time | (week) | date | Nb pos/Nb tot | Mean Cta | Estimated concentrationb | genotype | ||
| A (female) | 09-649/1 | 24 | 27/04/09 | cloacal swab | 0/20 | na | na | |
| 09-783/2 | 29 | 03/06/09 | 0/20 | na | na | |||
| 09-1001/1 | 34 | 09/07/09 | 1/20 | 38.8 | 1.9E + 01 | nd | ||
| 09-1226/2 | 43 | 07/09/09 | 0/20 | na | na | |||
| 09-1408/4 | 49 | 14/10/09 | 0/20 | na | na | |||
| 09-1576/6 | 55 | 24/11/09 | 1/20 | 38.0 | 3.3E + 01 | nd | ||
| 10-1911/3 | 64 | 04/02/10 | 3/20 | 36.0 | 1.3E + 02 | nd | ||
| B (female) | 09-649/2 | 24 | 27/04/09 | cloacal swab | 0/20 | na | na | |
| 09-783/3 | 29 | 03/06/09 | 1/20 | 37.6 | 4.3E + 01 | nd | ||
| 09-1001/4 | 34 | 09/07/09 | 1/20 | 38.0 | 3.3E + 01 | nd | ||
| 09-1226/3 | 43 | 07/09/09 | 0/20 | na | na | |||
| 09-1408/5 | 49 | 14/10/09 | 0/20 | na | na | |||
| 09-1576/5 | 55 | 24/11/09 | 1/20 | 39.3 | 1.4E + 01 | nd | ||
| 10-1911/4 | 64 | 04/02/10 | 0/20 | – | – | |||
| C (female) | 09-649/3 | 25 | 27/04/09 | cloacal swab | 1/20 | 37.7 | 4.1E + 01 | nd |
| 09-783/5 | 30 | 03/06/09 | 0/20 | na | na | |||
| 09-1001/3 | 35 | 09/07/09 | 1/20 | 36.7 | 7.8E + 01 | nd | ||
| 09-1226/4 | 44 | 07/09/09 | 0/20 | na | na | |||
| 09-1408/3 | 49 | 14/10/09 | 0/20 | na | na | |||
| 09-1576/4 | 55 | 24/11/09 | 1/20 | 39.7 | 3.3E + 00 | nd | ||
| 10-1911/1 | 64 | 04/02/10 | 0/20 | – | – | |||
| D (female) | 09-649/4 | 25 | 27/04/09 | cloacal swab | 3/20 | 32.2 | 1.6E + 03 | E/B_new |
| 09-783/4 | 30 | 03/06/09 | 0/20 | na | na | |||
| 09-1001/2 | 35 | 09/07/09 | 0/20 | na | na | |||
| 09-1226/1 | 44 | 07/09/09 | 0/20 | na | na | |||
| 09-1408/2 | 49 | 14/10/09 | 0/20 | na | na | |||
| 09-1576/3 | 55 | 24/11/09 | 0/20 | na | na | |||
| 10-1911/2 | 64 | 04/02/10 | 0/20 | na | na | |||
| E (male 1) | 09-649/5 | – | 27/04/09 | cloacal swab | 0/20 | na | na | |
| 09-783/1 | – | 03/06/09 | 0/20 | na | na | |||
| 09-1001/5 | – | 09/07/09 | 0/20 | na | na | |||
| F (male 2) | 09-1226/5 | – | 07/09/09 | cloacal swab | 0/20 | na | na | |
| 09-1408/1 | – | 14/10/09 | 0/20 | na | na | |||
| 09-1576/1 | – | 24/11/09 | 2/10 | 39.1 | 1.6E + 01 | nd | ||
| G (male 3) | 10-1911/5 | – | 04/02/10 | cloacal swab | 0/20 | na | na | |
| 23S RT-PCR | ||||||||
|---|---|---|---|---|---|---|---|---|
| Flock | Id sampling | Age | Sampling | Sample | C. psittaci | |||
| time | (week) | date | Nb pos/Nb tot | Mean Cta | Estimated concentrationb | genotype | ||
| A (female) | 09-649/1 | 24 | 27/04/09 | cloacal swab | 0/20 | na | na | |
| 09-783/2 | 29 | 03/06/09 | 0/20 | na | na | |||
| 09-1001/1 | 34 | 09/07/09 | 1/20 | 38.8 | 1.9E + 01 | nd | ||
| 09-1226/2 | 43 | 07/09/09 | 0/20 | na | na | |||
| 09-1408/4 | 49 | 14/10/09 | 0/20 | na | na | |||
| 09-1576/6 | 55 | 24/11/09 | 1/20 | 38.0 | 3.3E + 01 | nd | ||
| 10-1911/3 | 64 | 04/02/10 | 3/20 | 36.0 | 1.3E + 02 | nd | ||
| B (female) | 09-649/2 | 24 | 27/04/09 | cloacal swab | 0/20 | na | na | |
| 09-783/3 | 29 | 03/06/09 | 1/20 | 37.6 | 4.3E + 01 | nd | ||
| 09-1001/4 | 34 | 09/07/09 | 1/20 | 38.0 | 3.3E + 01 | nd | ||
| 09-1226/3 | 43 | 07/09/09 | 0/20 | na | na | |||
| 09-1408/5 | 49 | 14/10/09 | 0/20 | na | na | |||
| 09-1576/5 | 55 | 24/11/09 | 1/20 | 39.3 | 1.4E + 01 | nd | ||
| 10-1911/4 | 64 | 04/02/10 | 0/20 | – | – | |||
| C (female) | 09-649/3 | 25 | 27/04/09 | cloacal swab | 1/20 | 37.7 | 4.1E + 01 | nd |
| 09-783/5 | 30 | 03/06/09 | 0/20 | na | na | |||
| 09-1001/3 | 35 | 09/07/09 | 1/20 | 36.7 | 7.8E + 01 | nd | ||
| 09-1226/4 | 44 | 07/09/09 | 0/20 | na | na | |||
| 09-1408/3 | 49 | 14/10/09 | 0/20 | na | na | |||
| 09-1576/4 | 55 | 24/11/09 | 1/20 | 39.7 | 3.3E + 00 | nd | ||
| 10-1911/1 | 64 | 04/02/10 | 0/20 | – | – | |||
| D (female) | 09-649/4 | 25 | 27/04/09 | cloacal swab | 3/20 | 32.2 | 1.6E + 03 | E/B_new |
| 09-783/4 | 30 | 03/06/09 | 0/20 | na | na | |||
| 09-1001/2 | 35 | 09/07/09 | 0/20 | na | na | |||
| 09-1226/1 | 44 | 07/09/09 | 0/20 | na | na | |||
| 09-1408/2 | 49 | 14/10/09 | 0/20 | na | na | |||
| 09-1576/3 | 55 | 24/11/09 | 0/20 | na | na | |||
| 10-1911/2 | 64 | 04/02/10 | 0/20 | na | na | |||
| E (male 1) | 09-649/5 | – | 27/04/09 | cloacal swab | 0/20 | na | na | |
| 09-783/1 | – | 03/06/09 | 0/20 | na | na | |||
| 09-1001/5 | – | 09/07/09 | 0/20 | na | na | |||
| F (male 2) | 09-1226/5 | – | 07/09/09 | cloacal swab | 0/20 | na | na | |
| 09-1408/1 | – | 14/10/09 | 0/20 | na | na | |||
| 09-1576/1 | – | 24/11/09 | 2/10 | 39.1 | 1.6E + 01 | nd | ||
| G (male 3) | 10-1911/5 | – | 04/02/10 | cloacal swab | 0/20 | na | na | |
a for positive samples (2μl of ADN tested).
b for positive samples (mean number of genome/swab).
na: not applicable.
nd: not done.
Characteristics of in kinetic investigated laying duck flocks (n = 4) dedicated to the mule duck production by artificial insemination. Male flocks used for the semen production were also investigated (n = 3).
| 23S RT-PCR | ||||||||
|---|---|---|---|---|---|---|---|---|
| Flock | Id sampling | Age | Sampling | Sample | C. psittaci | |||
| time | (week) | date | Nb pos/Nb tot | Mean Cta | Estimated concentrationb | genotype | ||
| A (female) | 09-649/1 | 24 | 27/04/09 | cloacal swab | 0/20 | na | na | |
| 09-783/2 | 29 | 03/06/09 | 0/20 | na | na | |||
| 09-1001/1 | 34 | 09/07/09 | 1/20 | 38.8 | 1.9E + 01 | nd | ||
| 09-1226/2 | 43 | 07/09/09 | 0/20 | na | na | |||
| 09-1408/4 | 49 | 14/10/09 | 0/20 | na | na | |||
| 09-1576/6 | 55 | 24/11/09 | 1/20 | 38.0 | 3.3E + 01 | nd | ||
| 10-1911/3 | 64 | 04/02/10 | 3/20 | 36.0 | 1.3E + 02 | nd | ||
| B (female) | 09-649/2 | 24 | 27/04/09 | cloacal swab | 0/20 | na | na | |
| 09-783/3 | 29 | 03/06/09 | 1/20 | 37.6 | 4.3E + 01 | nd | ||
| 09-1001/4 | 34 | 09/07/09 | 1/20 | 38.0 | 3.3E + 01 | nd | ||
| 09-1226/3 | 43 | 07/09/09 | 0/20 | na | na | |||
| 09-1408/5 | 49 | 14/10/09 | 0/20 | na | na | |||
| 09-1576/5 | 55 | 24/11/09 | 1/20 | 39.3 | 1.4E + 01 | nd | ||
| 10-1911/4 | 64 | 04/02/10 | 0/20 | – | – | |||
| C (female) | 09-649/3 | 25 | 27/04/09 | cloacal swab | 1/20 | 37.7 | 4.1E + 01 | nd |
| 09-783/5 | 30 | 03/06/09 | 0/20 | na | na | |||
| 09-1001/3 | 35 | 09/07/09 | 1/20 | 36.7 | 7.8E + 01 | nd | ||
| 09-1226/4 | 44 | 07/09/09 | 0/20 | na | na | |||
| 09-1408/3 | 49 | 14/10/09 | 0/20 | na | na | |||
| 09-1576/4 | 55 | 24/11/09 | 1/20 | 39.7 | 3.3E + 00 | nd | ||
| 10-1911/1 | 64 | 04/02/10 | 0/20 | – | – | |||
| D (female) | 09-649/4 | 25 | 27/04/09 | cloacal swab | 3/20 | 32.2 | 1.6E + 03 | E/B_new |
| 09-783/4 | 30 | 03/06/09 | 0/20 | na | na | |||
| 09-1001/2 | 35 | 09/07/09 | 0/20 | na | na | |||
| 09-1226/1 | 44 | 07/09/09 | 0/20 | na | na | |||
| 09-1408/2 | 49 | 14/10/09 | 0/20 | na | na | |||
| 09-1576/3 | 55 | 24/11/09 | 0/20 | na | na | |||
| 10-1911/2 | 64 | 04/02/10 | 0/20 | na | na | |||
| E (male 1) | 09-649/5 | – | 27/04/09 | cloacal swab | 0/20 | na | na | |
| 09-783/1 | – | 03/06/09 | 0/20 | na | na | |||
| 09-1001/5 | – | 09/07/09 | 0/20 | na | na | |||
| F (male 2) | 09-1226/5 | – | 07/09/09 | cloacal swab | 0/20 | na | na | |
| 09-1408/1 | – | 14/10/09 | 0/20 | na | na | |||
| 09-1576/1 | – | 24/11/09 | 2/10 | 39.1 | 1.6E + 01 | nd | ||
| G (male 3) | 10-1911/5 | – | 04/02/10 | cloacal swab | 0/20 | na | na | |
| 23S RT-PCR | ||||||||
|---|---|---|---|---|---|---|---|---|
| Flock | Id sampling | Age | Sampling | Sample | C. psittaci | |||
| time | (week) | date | Nb pos/Nb tot | Mean Cta | Estimated concentrationb | genotype | ||
| A (female) | 09-649/1 | 24 | 27/04/09 | cloacal swab | 0/20 | na | na | |
| 09-783/2 | 29 | 03/06/09 | 0/20 | na | na | |||
| 09-1001/1 | 34 | 09/07/09 | 1/20 | 38.8 | 1.9E + 01 | nd | ||
| 09-1226/2 | 43 | 07/09/09 | 0/20 | na | na | |||
| 09-1408/4 | 49 | 14/10/09 | 0/20 | na | na | |||
| 09-1576/6 | 55 | 24/11/09 | 1/20 | 38.0 | 3.3E + 01 | nd | ||
| 10-1911/3 | 64 | 04/02/10 | 3/20 | 36.0 | 1.3E + 02 | nd | ||
| B (female) | 09-649/2 | 24 | 27/04/09 | cloacal swab | 0/20 | na | na | |
| 09-783/3 | 29 | 03/06/09 | 1/20 | 37.6 | 4.3E + 01 | nd | ||
| 09-1001/4 | 34 | 09/07/09 | 1/20 | 38.0 | 3.3E + 01 | nd | ||
| 09-1226/3 | 43 | 07/09/09 | 0/20 | na | na | |||
| 09-1408/5 | 49 | 14/10/09 | 0/20 | na | na | |||
| 09-1576/5 | 55 | 24/11/09 | 1/20 | 39.3 | 1.4E + 01 | nd | ||
| 10-1911/4 | 64 | 04/02/10 | 0/20 | – | – | |||
| C (female) | 09-649/3 | 25 | 27/04/09 | cloacal swab | 1/20 | 37.7 | 4.1E + 01 | nd |
| 09-783/5 | 30 | 03/06/09 | 0/20 | na | na | |||
| 09-1001/3 | 35 | 09/07/09 | 1/20 | 36.7 | 7.8E + 01 | nd | ||
| 09-1226/4 | 44 | 07/09/09 | 0/20 | na | na | |||
| 09-1408/3 | 49 | 14/10/09 | 0/20 | na | na | |||
| 09-1576/4 | 55 | 24/11/09 | 1/20 | 39.7 | 3.3E + 00 | nd | ||
| 10-1911/1 | 64 | 04/02/10 | 0/20 | – | – | |||
| D (female) | 09-649/4 | 25 | 27/04/09 | cloacal swab | 3/20 | 32.2 | 1.6E + 03 | E/B_new |
| 09-783/4 | 30 | 03/06/09 | 0/20 | na | na | |||
| 09-1001/2 | 35 | 09/07/09 | 0/20 | na | na | |||
| 09-1226/1 | 44 | 07/09/09 | 0/20 | na | na | |||
| 09-1408/2 | 49 | 14/10/09 | 0/20 | na | na | |||
| 09-1576/3 | 55 | 24/11/09 | 0/20 | na | na | |||
| 10-1911/2 | 64 | 04/02/10 | 0/20 | na | na | |||
| E (male 1) | 09-649/5 | – | 27/04/09 | cloacal swab | 0/20 | na | na | |
| 09-783/1 | – | 03/06/09 | 0/20 | na | na | |||
| 09-1001/5 | – | 09/07/09 | 0/20 | na | na | |||
| F (male 2) | 09-1226/5 | – | 07/09/09 | cloacal swab | 0/20 | na | na | |
| 09-1408/1 | – | 14/10/09 | 0/20 | na | na | |||
| 09-1576/1 | – | 24/11/09 | 2/10 | 39.1 | 1.6E + 01 | nd | ||
| G (male 3) | 10-1911/5 | – | 04/02/10 | cloacal swab | 0/20 | na | na | |
a for positive samples (2μl of ADN tested).
b for positive samples (mean number of genome/swab).
na: not applicable.
nd: not done.
Randomly selected mule duck flocks.
In seven randomly selected mule duck flocks housed on distinct farms, 15 randomly selected ducks per flock were subjected to swabbing when aged 2, 4, 8, 16 and 17 weeks, according to the different breeding stages (Table 2). These mule duck flocks were not linked to the breeder duck flocks above described, but belonged to the same professional organization.
Characteristics of in kinetic investigated mule duck flocks (n = 7).
| 23S RT-PCR | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Parental | |||||||||||
| Id sampling | Age | Implementation | flock | Nb pos/ | Mean | Estimated | C. psittaci | ||||
| Flock | time | Place | (days) | Effective | date | number | Sample | Nb tot | Cta | concentrationb | genotype |
| A | 09-383/4 | indoors | 16 | 3152 | 19/02/09 | 1 | cloacal swab | 0/15 | na | na | |
| 09-523/15 | indoors | 30 | 3152 | 0/15 | na | na | |||||
| 09-674/4 | outdoors | 56 | 3152 | 15/15 | 26.6 | 7.0E + 04 | E/B_06-859 | ||||
| 09–745/3 | outdoors | 85 | 3152 | 13/15 | 35.5 | 1.8E + 02 | E/B_06-859 | ||||
| 09-745/12 | force-feeding place_1 | 97 | 1610 | 15/05/09 | tracheal swab | 2/15 | 34.6 | 6.4E + 02 | E/B_06-859 | ||
| 09-874/1 | force-feeding place_2 | 97 | 1330 | 4/15 | 36.5 | 8.9E + 01 | E/B_06-859 | ||||
| B | 09-523/14 | indoors | 15 | 6763 | 19/03/09 | 2 | cloacal swab | 0/15 | na | na | |
| 09-674/1 | outdoors | 28 | 6763 | 2/15 | 36.0 | 1.3E + 02 | nd | ||||
| 09-745/4 | outdoors | 57 | 6763 | 13/15 | 33.8 | 5.5E + 02 | C_GR9 | ||||
| 09-874/5 | outdoors | 82 | 6763 | 11/15 | 34.8 | 2.8E + 02 | C_GR9 | ||||
| 09-908/2 | force-feeding place_1 | 93 | 1300 | 11/06/09 | tracheal swab | 2/15 | 39.1 | 1.6E + 01 | nd | ||
| 09-908/1 | force-feeding place_2 | 97 | 1048 | 2/15 | 38.8 | 1.9E + 01 | nd | ||||
| C | 09-383/3 | indoors | 15 | 3254 | 13/02/09 | 3 | cloacal swab | 0/15 | na | na | |
| 09-523/13 | outdoors | 32 | 3254 | 0/15 | na | na | |||||
| 09-674/6 | outdoors | 62 | 3254 | 0/15 | na | na | |||||
| 09-674/7 | outdoors | 77 | 3254 | 1/15 | 36.5 | 1.0E + 02 | nd | ||||
| 09-745/5 | force-feeding place_1 | 91 | 1328 | 07/05/09 | tracheal swab | 1/15 | 39.3 | 1.4E + 01 | nd | ||
| 09-745/6 | force-feeding place_2 | 91 | 1750 | 04/05/09 | 2/15 | 38.7 | 2.0E + 01 | nd | |||
| D | 09-383/5 | indoors | 15 | 2213 | 20/02/09 | 1 | cloacal swab | 0/15 | na | na | |
| 09-523/5 | outdoors | 29 | 2213 | 0/15 | na | na | |||||
| 09-674/8 | outdoors | 55 | 2213 | 12/15 | 30.8 | 4.1E + 03 | C_GR9_new | ||||
| 09-745/1 | outdoors | 77 | 2213 | 10/15 | 34.8 | 2.8E + 02 | C_GR9_new | ||||
| 09-745/8 | force-feeding place_1 | 90 | 1260 | 11/05/09 | tracheal swab | 1/15 | 36.5 | 8.9E + 01 | nd | ||
| 09-745/9 | force-feeding place_2 | 90 | 840 | 0/15 | na | na | |||||
| E | 09-523/1 | indoors | 4 | 2530 | 12/03/09 | 4 | cloacal swab | 0/15 | na | na | |
| 09-523/3 | outdoors | 27 | 2530 | 0/15 | na | na | |||||
| 09-674/2 | outdoors | 54 | 2530 | 0/15 | na | na | |||||
| 09-874/2 | outdoors | 78 | 2530 | 12/15 | 32.4 | 1.4E + 03 | E/B_06-859 | ||||
| 09-874/6 | force-feeding place_1 | 95 | 1330 | 03/06/09 | tracheal swab | 2/15 | 38.5 | 2.3E + 01 | nd | ||
| 09-874/7 | force-feeding place_2 | 95 | 1048 | 1/15 | 35.6 | 1.6E + 02 | C_GR9_new | ||||
| F | 09-239 | indoors | 37 | 3745 | 31/12/08 | 5 | cloacal swab | 0/15 | na | na | |
| 09-383/1 | outdoors | 50 | 3745 | 0/15 | na | na | |||||
| 09-383/7 | outdoors | 66 | 3745 | 15/15 | 24.8 | 2.3E + 05 | C_GR9 | ||||
| 09-523/4 | outdoors | 80 | 3745 | 11/15 | 29.2 | 1.2E + 04 | C_GR9 | ||||
| 09-523/11 | force-feeding place_1 | 97 | 1300 | 26/03/09 | tracheal swab | 0/15 | na | na | |||
| 09-523/10 | force-feeding place_2 | 97 | 2240 | 1/15 | 38.7 | 2.0E + 01 | E/B_new | ||||
| G | 09-523/9 | indoors | 18 | 3570 | 27/02/09 | 6 | cloacal swab | 0/15 | na | na | |
| 09-523/8 | outdoors | 32 | 3570 | 0/15 | na | na | |||||
| 09-674/3 | outdoors | 60 | 3570 | 0/15 | na | na | |||||
| 09-745/7 | outdoors | 81 | 3570 | 0/15 | na | na | |||||
| 09-874/3 | force-feeding place_1 | 95 | 1330 | 19/05/09 | tracheal swab | 0/15 | na | na | |||
| 09-874/4 | force-feeding place_2 | 95 | 1330 | 0/15 | na | na | |||||
| 23S RT-PCR | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Parental | |||||||||||
| Id sampling | Age | Implementation | flock | Nb pos/ | Mean | Estimated | C. psittaci | ||||
| Flock | time | Place | (days) | Effective | date | number | Sample | Nb tot | Cta | concentrationb | genotype |
| A | 09-383/4 | indoors | 16 | 3152 | 19/02/09 | 1 | cloacal swab | 0/15 | na | na | |
| 09-523/15 | indoors | 30 | 3152 | 0/15 | na | na | |||||
| 09-674/4 | outdoors | 56 | 3152 | 15/15 | 26.6 | 7.0E + 04 | E/B_06-859 | ||||
| 09–745/3 | outdoors | 85 | 3152 | 13/15 | 35.5 | 1.8E + 02 | E/B_06-859 | ||||
| 09-745/12 | force-feeding place_1 | 97 | 1610 | 15/05/09 | tracheal swab | 2/15 | 34.6 | 6.4E + 02 | E/B_06-859 | ||
| 09-874/1 | force-feeding place_2 | 97 | 1330 | 4/15 | 36.5 | 8.9E + 01 | E/B_06-859 | ||||
| B | 09-523/14 | indoors | 15 | 6763 | 19/03/09 | 2 | cloacal swab | 0/15 | na | na | |
| 09-674/1 | outdoors | 28 | 6763 | 2/15 | 36.0 | 1.3E + 02 | nd | ||||
| 09-745/4 | outdoors | 57 | 6763 | 13/15 | 33.8 | 5.5E + 02 | C_GR9 | ||||
| 09-874/5 | outdoors | 82 | 6763 | 11/15 | 34.8 | 2.8E + 02 | C_GR9 | ||||
| 09-908/2 | force-feeding place_1 | 93 | 1300 | 11/06/09 | tracheal swab | 2/15 | 39.1 | 1.6E + 01 | nd | ||
| 09-908/1 | force-feeding place_2 | 97 | 1048 | 2/15 | 38.8 | 1.9E + 01 | nd | ||||
| C | 09-383/3 | indoors | 15 | 3254 | 13/02/09 | 3 | cloacal swab | 0/15 | na | na | |
| 09-523/13 | outdoors | 32 | 3254 | 0/15 | na | na | |||||
| 09-674/6 | outdoors | 62 | 3254 | 0/15 | na | na | |||||
| 09-674/7 | outdoors | 77 | 3254 | 1/15 | 36.5 | 1.0E + 02 | nd | ||||
| 09-745/5 | force-feeding place_1 | 91 | 1328 | 07/05/09 | tracheal swab | 1/15 | 39.3 | 1.4E + 01 | nd | ||
| 09-745/6 | force-feeding place_2 | 91 | 1750 | 04/05/09 | 2/15 | 38.7 | 2.0E + 01 | nd | |||
| D | 09-383/5 | indoors | 15 | 2213 | 20/02/09 | 1 | cloacal swab | 0/15 | na | na | |
| 09-523/5 | outdoors | 29 | 2213 | 0/15 | na | na | |||||
| 09-674/8 | outdoors | 55 | 2213 | 12/15 | 30.8 | 4.1E + 03 | C_GR9_new | ||||
| 09-745/1 | outdoors | 77 | 2213 | 10/15 | 34.8 | 2.8E + 02 | C_GR9_new | ||||
| 09-745/8 | force-feeding place_1 | 90 | 1260 | 11/05/09 | tracheal swab | 1/15 | 36.5 | 8.9E + 01 | nd | ||
| 09-745/9 | force-feeding place_2 | 90 | 840 | 0/15 | na | na | |||||
| E | 09-523/1 | indoors | 4 | 2530 | 12/03/09 | 4 | cloacal swab | 0/15 | na | na | |
| 09-523/3 | outdoors | 27 | 2530 | 0/15 | na | na | |||||
| 09-674/2 | outdoors | 54 | 2530 | 0/15 | na | na | |||||
| 09-874/2 | outdoors | 78 | 2530 | 12/15 | 32.4 | 1.4E + 03 | E/B_06-859 | ||||
| 09-874/6 | force-feeding place_1 | 95 | 1330 | 03/06/09 | tracheal swab | 2/15 | 38.5 | 2.3E + 01 | nd | ||
| 09-874/7 | force-feeding place_2 | 95 | 1048 | 1/15 | 35.6 | 1.6E + 02 | C_GR9_new | ||||
| F | 09-239 | indoors | 37 | 3745 | 31/12/08 | 5 | cloacal swab | 0/15 | na | na | |
| 09-383/1 | outdoors | 50 | 3745 | 0/15 | na | na | |||||
| 09-383/7 | outdoors | 66 | 3745 | 15/15 | 24.8 | 2.3E + 05 | C_GR9 | ||||
| 09-523/4 | outdoors | 80 | 3745 | 11/15 | 29.2 | 1.2E + 04 | C_GR9 | ||||
| 09-523/11 | force-feeding place_1 | 97 | 1300 | 26/03/09 | tracheal swab | 0/15 | na | na | |||
| 09-523/10 | force-feeding place_2 | 97 | 2240 | 1/15 | 38.7 | 2.0E + 01 | E/B_new | ||||
| G | 09-523/9 | indoors | 18 | 3570 | 27/02/09 | 6 | cloacal swab | 0/15 | na | na | |
| 09-523/8 | outdoors | 32 | 3570 | 0/15 | na | na | |||||
| 09-674/3 | outdoors | 60 | 3570 | 0/15 | na | na | |||||
| 09-745/7 | outdoors | 81 | 3570 | 0/15 | na | na | |||||
| 09-874/3 | force-feeding place_1 | 95 | 1330 | 19/05/09 | tracheal swab | 0/15 | na | na | |||
| 09-874/4 | force-feeding place_2 | 95 | 1330 | 0/15 | na | na | |||||
a for positive samples (2μl of ADN).
b for positive samples (mean number of genome/swab).
na: not applicable.
nd: not done.
Characteristics of in kinetic investigated mule duck flocks (n = 7).
| 23S RT-PCR | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Parental | |||||||||||
| Id sampling | Age | Implementation | flock | Nb pos/ | Mean | Estimated | C. psittaci | ||||
| Flock | time | Place | (days) | Effective | date | number | Sample | Nb tot | Cta | concentrationb | genotype |
| A | 09-383/4 | indoors | 16 | 3152 | 19/02/09 | 1 | cloacal swab | 0/15 | na | na | |
| 09-523/15 | indoors | 30 | 3152 | 0/15 | na | na | |||||
| 09-674/4 | outdoors | 56 | 3152 | 15/15 | 26.6 | 7.0E + 04 | E/B_06-859 | ||||
| 09–745/3 | outdoors | 85 | 3152 | 13/15 | 35.5 | 1.8E + 02 | E/B_06-859 | ||||
| 09-745/12 | force-feeding place_1 | 97 | 1610 | 15/05/09 | tracheal swab | 2/15 | 34.6 | 6.4E + 02 | E/B_06-859 | ||
| 09-874/1 | force-feeding place_2 | 97 | 1330 | 4/15 | 36.5 | 8.9E + 01 | E/B_06-859 | ||||
| B | 09-523/14 | indoors | 15 | 6763 | 19/03/09 | 2 | cloacal swab | 0/15 | na | na | |
| 09-674/1 | outdoors | 28 | 6763 | 2/15 | 36.0 | 1.3E + 02 | nd | ||||
| 09-745/4 | outdoors | 57 | 6763 | 13/15 | 33.8 | 5.5E + 02 | C_GR9 | ||||
| 09-874/5 | outdoors | 82 | 6763 | 11/15 | 34.8 | 2.8E + 02 | C_GR9 | ||||
| 09-908/2 | force-feeding place_1 | 93 | 1300 | 11/06/09 | tracheal swab | 2/15 | 39.1 | 1.6E + 01 | nd | ||
| 09-908/1 | force-feeding place_2 | 97 | 1048 | 2/15 | 38.8 | 1.9E + 01 | nd | ||||
| C | 09-383/3 | indoors | 15 | 3254 | 13/02/09 | 3 | cloacal swab | 0/15 | na | na | |
| 09-523/13 | outdoors | 32 | 3254 | 0/15 | na | na | |||||
| 09-674/6 | outdoors | 62 | 3254 | 0/15 | na | na | |||||
| 09-674/7 | outdoors | 77 | 3254 | 1/15 | 36.5 | 1.0E + 02 | nd | ||||
| 09-745/5 | force-feeding place_1 | 91 | 1328 | 07/05/09 | tracheal swab | 1/15 | 39.3 | 1.4E + 01 | nd | ||
| 09-745/6 | force-feeding place_2 | 91 | 1750 | 04/05/09 | 2/15 | 38.7 | 2.0E + 01 | nd | |||
| D | 09-383/5 | indoors | 15 | 2213 | 20/02/09 | 1 | cloacal swab | 0/15 | na | na | |
| 09-523/5 | outdoors | 29 | 2213 | 0/15 | na | na | |||||
| 09-674/8 | outdoors | 55 | 2213 | 12/15 | 30.8 | 4.1E + 03 | C_GR9_new | ||||
| 09-745/1 | outdoors | 77 | 2213 | 10/15 | 34.8 | 2.8E + 02 | C_GR9_new | ||||
| 09-745/8 | force-feeding place_1 | 90 | 1260 | 11/05/09 | tracheal swab | 1/15 | 36.5 | 8.9E + 01 | nd | ||
| 09-745/9 | force-feeding place_2 | 90 | 840 | 0/15 | na | na | |||||
| E | 09-523/1 | indoors | 4 | 2530 | 12/03/09 | 4 | cloacal swab | 0/15 | na | na | |
| 09-523/3 | outdoors | 27 | 2530 | 0/15 | na | na | |||||
| 09-674/2 | outdoors | 54 | 2530 | 0/15 | na | na | |||||
| 09-874/2 | outdoors | 78 | 2530 | 12/15 | 32.4 | 1.4E + 03 | E/B_06-859 | ||||
| 09-874/6 | force-feeding place_1 | 95 | 1330 | 03/06/09 | tracheal swab | 2/15 | 38.5 | 2.3E + 01 | nd | ||
| 09-874/7 | force-feeding place_2 | 95 | 1048 | 1/15 | 35.6 | 1.6E + 02 | C_GR9_new | ||||
| F | 09-239 | indoors | 37 | 3745 | 31/12/08 | 5 | cloacal swab | 0/15 | na | na | |
| 09-383/1 | outdoors | 50 | 3745 | 0/15 | na | na | |||||
| 09-383/7 | outdoors | 66 | 3745 | 15/15 | 24.8 | 2.3E + 05 | C_GR9 | ||||
| 09-523/4 | outdoors | 80 | 3745 | 11/15 | 29.2 | 1.2E + 04 | C_GR9 | ||||
| 09-523/11 | force-feeding place_1 | 97 | 1300 | 26/03/09 | tracheal swab | 0/15 | na | na | |||
| 09-523/10 | force-feeding place_2 | 97 | 2240 | 1/15 | 38.7 | 2.0E + 01 | E/B_new | ||||
| G | 09-523/9 | indoors | 18 | 3570 | 27/02/09 | 6 | cloacal swab | 0/15 | na | na | |
| 09-523/8 | outdoors | 32 | 3570 | 0/15 | na | na | |||||
| 09-674/3 | outdoors | 60 | 3570 | 0/15 | na | na | |||||
| 09-745/7 | outdoors | 81 | 3570 | 0/15 | na | na | |||||
| 09-874/3 | force-feeding place_1 | 95 | 1330 | 19/05/09 | tracheal swab | 0/15 | na | na | |||
| 09-874/4 | force-feeding place_2 | 95 | 1330 | 0/15 | na | na | |||||
| 23S RT-PCR | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Parental | |||||||||||
| Id sampling | Age | Implementation | flock | Nb pos/ | Mean | Estimated | C. psittaci | ||||
| Flock | time | Place | (days) | Effective | date | number | Sample | Nb tot | Cta | concentrationb | genotype |
| A | 09-383/4 | indoors | 16 | 3152 | 19/02/09 | 1 | cloacal swab | 0/15 | na | na | |
| 09-523/15 | indoors | 30 | 3152 | 0/15 | na | na | |||||
| 09-674/4 | outdoors | 56 | 3152 | 15/15 | 26.6 | 7.0E + 04 | E/B_06-859 | ||||
| 09–745/3 | outdoors | 85 | 3152 | 13/15 | 35.5 | 1.8E + 02 | E/B_06-859 | ||||
| 09-745/12 | force-feeding place_1 | 97 | 1610 | 15/05/09 | tracheal swab | 2/15 | 34.6 | 6.4E + 02 | E/B_06-859 | ||
| 09-874/1 | force-feeding place_2 | 97 | 1330 | 4/15 | 36.5 | 8.9E + 01 | E/B_06-859 | ||||
| B | 09-523/14 | indoors | 15 | 6763 | 19/03/09 | 2 | cloacal swab | 0/15 | na | na | |
| 09-674/1 | outdoors | 28 | 6763 | 2/15 | 36.0 | 1.3E + 02 | nd | ||||
| 09-745/4 | outdoors | 57 | 6763 | 13/15 | 33.8 | 5.5E + 02 | C_GR9 | ||||
| 09-874/5 | outdoors | 82 | 6763 | 11/15 | 34.8 | 2.8E + 02 | C_GR9 | ||||
| 09-908/2 | force-feeding place_1 | 93 | 1300 | 11/06/09 | tracheal swab | 2/15 | 39.1 | 1.6E + 01 | nd | ||
| 09-908/1 | force-feeding place_2 | 97 | 1048 | 2/15 | 38.8 | 1.9E + 01 | nd | ||||
| C | 09-383/3 | indoors | 15 | 3254 | 13/02/09 | 3 | cloacal swab | 0/15 | na | na | |
| 09-523/13 | outdoors | 32 | 3254 | 0/15 | na | na | |||||
| 09-674/6 | outdoors | 62 | 3254 | 0/15 | na | na | |||||
| 09-674/7 | outdoors | 77 | 3254 | 1/15 | 36.5 | 1.0E + 02 | nd | ||||
| 09-745/5 | force-feeding place_1 | 91 | 1328 | 07/05/09 | tracheal swab | 1/15 | 39.3 | 1.4E + 01 | nd | ||
| 09-745/6 | force-feeding place_2 | 91 | 1750 | 04/05/09 | 2/15 | 38.7 | 2.0E + 01 | nd | |||
| D | 09-383/5 | indoors | 15 | 2213 | 20/02/09 | 1 | cloacal swab | 0/15 | na | na | |
| 09-523/5 | outdoors | 29 | 2213 | 0/15 | na | na | |||||
| 09-674/8 | outdoors | 55 | 2213 | 12/15 | 30.8 | 4.1E + 03 | C_GR9_new | ||||
| 09-745/1 | outdoors | 77 | 2213 | 10/15 | 34.8 | 2.8E + 02 | C_GR9_new | ||||
| 09-745/8 | force-feeding place_1 | 90 | 1260 | 11/05/09 | tracheal swab | 1/15 | 36.5 | 8.9E + 01 | nd | ||
| 09-745/9 | force-feeding place_2 | 90 | 840 | 0/15 | na | na | |||||
| E | 09-523/1 | indoors | 4 | 2530 | 12/03/09 | 4 | cloacal swab | 0/15 | na | na | |
| 09-523/3 | outdoors | 27 | 2530 | 0/15 | na | na | |||||
| 09-674/2 | outdoors | 54 | 2530 | 0/15 | na | na | |||||
| 09-874/2 | outdoors | 78 | 2530 | 12/15 | 32.4 | 1.4E + 03 | E/B_06-859 | ||||
| 09-874/6 | force-feeding place_1 | 95 | 1330 | 03/06/09 | tracheal swab | 2/15 | 38.5 | 2.3E + 01 | nd | ||
| 09-874/7 | force-feeding place_2 | 95 | 1048 | 1/15 | 35.6 | 1.6E + 02 | C_GR9_new | ||||
| F | 09-239 | indoors | 37 | 3745 | 31/12/08 | 5 | cloacal swab | 0/15 | na | na | |
| 09-383/1 | outdoors | 50 | 3745 | 0/15 | na | na | |||||
| 09-383/7 | outdoors | 66 | 3745 | 15/15 | 24.8 | 2.3E + 05 | C_GR9 | ||||
| 09-523/4 | outdoors | 80 | 3745 | 11/15 | 29.2 | 1.2E + 04 | C_GR9 | ||||
| 09-523/11 | force-feeding place_1 | 97 | 1300 | 26/03/09 | tracheal swab | 0/15 | na | na | |||
| 09-523/10 | force-feeding place_2 | 97 | 2240 | 1/15 | 38.7 | 2.0E + 01 | E/B_new | ||||
| G | 09-523/9 | indoors | 18 | 3570 | 27/02/09 | 6 | cloacal swab | 0/15 | na | na | |
| 09-523/8 | outdoors | 32 | 3570 | 0/15 | na | na | |||||
| 09-674/3 | outdoors | 60 | 3570 | 0/15 | na | na | |||||
| 09-745/7 | outdoors | 81 | 3570 | 0/15 | na | na | |||||
| 09-874/3 | force-feeding place_1 | 95 | 1330 | 19/05/09 | tracheal swab | 0/15 | na | na | |||
| 09-874/4 | force-feeding place_2 | 95 | 1330 | 0/15 | na | na | |||||
a for positive samples (2μl of ADN).
b for positive samples (mean number of genome/swab).
na: not applicable.
nd: not done.
Samples from birds linked to human cases of psittacosis.
In five flocks linked to human psittacosis cases, 20 birds per suspected flock were subjected to cloacal swabbing, including the male flock whose semen was used for insemination. Tissue samples (lungs, liver and intestine) were also collected from the progeny of one female flock in the hatchery (dead embryos and 1-day old ducklings) and cloacal swabs were taken from 4-week old ducks bred on the farm.
Human samples
Six people involved in the mule duck production process (breeder duck farms or hatcheries) showed signs of respiratory disease in late 2009. Four of them had to be hospitalized. Aliquots of early and late serum from each hospitalized patient were collected, as were throat swabs after medication (Table 3A).
Summaruy of data from human and animal investigations around human cases of psittacosis.
 |
|
Summaruy of data from human and animal investigations around human cases of psittacosis.
|
|
Direct detection of Chlamydiae from samples
DNA was extracted from bird samples (swabs or tissues) using a QIAamp DNA Mini Kit (Qiagen, Courtaboeuf, France). DNA from human throat swabs was extracted using the automated MagNA Pure DNA extraction kit (Roche Diagnostics, Meylan, France) (De Martino et al., 2006).
A Chlamydiaceae-specific real-time PCR (RT-PCR) targeting the 23S rDNA gene was used in this study (Ehricht et al., 2006). All samples with a cycle threshold over 40 were considered as negatives. DNA concentration for each positive sample was determined by comparison with DNA purified from cell culture containing defined numbers of genome copies of C. psittaci (kindly provided by Dr K. Sachse, Friedrich-Loeffler-Institut, Jena, Germany). The standard curve and the number of genome copies were automatically determined by the 7500 software (Applied Biosystems, France).
Serology of human samples
A commercial immunofluorescence test was used (Chlamydia MIF, Focus, Eurobio, France) to detect C. psittaci antibodies, as previously described (Laroucau et al., 2009a).
Human psittacosis case definition
In the present study, a patient with fever or at least respiratory symptoms having been in contact with ducks or their products was regarded as a suspected case. A possible case was a suspected case combined with an epidemiological link to a confirmed case. A probable case was a suspected case combined with an IgG titer >32. A confirmed case was a suspected case coupled with the positive detection of C. psittaci by PCR in a respiratory sample, seroconversion, or a 4-fold increase in IgG titer.
Genotyping
C. psittaci-specific RT-PCR
The ompA-based RT-PCR assay specific to C. psittaci was conducted as described (Pantchev et al., 2009).
OmpA analysis
Partial ompA gene fragments from human and bird samples were amplified as described previously using 3GPF (5′-ACGCATGCAAGACACTCCTCAAAGCC-3′) and 5GPB (5′-ACGAATTCCTAGGTTCTGATAGCGGGAC-3′) (Kaltenboeck, Kousoulas and Storz 1993). Nucleotide sequences were deposited under GenBank (LN624457 and LN624458) and compared to the ompA sequences from C. psittaci strains representing all currently recognized types and subtypes (Sachse et al., 2008). Multiple sequence alignments were carried out with hierarchical clustering using the free MultAlin software (http://multalin.toulouse.inra.fr/multalin/multalin.html) (Corpet 1988). The following strains and sequences were used for comparison: VS1 (genotype A-VS1, GenBank acc. no. AF269281.1), 6BC (A-6BC, X56980.1), 84–55 (A-8455, Y16561.1), CP3 (B, AF269265.1), GR9 (C, L25436.1), NJ1 (D-NJ1, AF269266.1), 9N (D-9N, EF375557.1), CPMN (E, X12647.1), WS/RT/E30 (EB-E30, AY762613.1), 06-859/1 (EB-859, EU159263.1), KKCP-1 (EB-KKCP, AB284062.1), VS225 (F, AF269259.1), M56 (M56, AF269268.1), WC (WC, AF269269.1), 1V (1V, EF028916.1), 6N (6N, EF197820.1), Mat116 (Mat116, AB284058.1), R54 (R54, AJ243525.1), Daruma-1981 (YP84, AB284065.1) and CPX0308 (CPX0308, AB284064.1).
Statistical analysis (mule duck survey only)
The prevalence and excretion level in the seven mule duck flocks investigated were analyzed separately. The difference between cloacal and tracheal swabs was assumed to be negligible as no difference was observed between both types of swabs collected from 15 birds sampled during the force-feeding period (unpublished data). Changes of environment occurred when ducks were about 32 days old (from indoors to outdoors) and 85 days old (from outdoors to indoors, at the beginning of the force-feeding period). Investigations covered the effect of age, flock and environment on the occurrence of C. psittaci in flocks.
In order to explore the non-linear shape of the age–prevalence relationship in monitored flocks, prevalence (proportion of PCR-positive birds) and shedding data (log of the DNA titer in positive birds) were fitted using a generalized additive mixed model (GAMM) (Wood 2006). The age (in days) was considered a fixed effect and the flock a random effect (flock and farm effects were confounded because a single flock was followed on each farm). Data analysis was performed with the R.2.3.0.1 (R development Core Team) mgcv package (Wood, release 1.7.24).
RESULTS
Randomly selected breeder flocks
Small quantities of Chlamydiaceae were detected in all the flocks studied. Data are summarized in Table 1. Positive samples were all re-analyzed with a C. psittaci-specific RT-PCR. ompA sequencing was possible for three samples collected at the same time from the same flock (Flock D, 27/04/09). A sequence comparison identified a new E/B subgenotype, with mutations in 3, 4 and 6 bases of 1038 identified in comparison to 06-859/1, KKCP-1 and WS/RT/E30, respectively (Sachse et al., 2008) (Fig. S1A, Supporting Information).
Randomly selected mule duck flocks
Chlamydiaceae were detected in most of the seven mule duck flocks studied. Detailed results are presented in Table 2. Three shedding patterns were observed:
Pattern 1: heavy shedding in most 8- and 12-week old birds (flocks A, B, D, E and F), but no shedding detected before these dates (except for flock B), residual and weak positivity in a few birds during the force-feeding period. For flock E, a large number of shedding birds were detected only at the week 12 sampling session.
Pattern 2: a little shedding in some birds from the end of pre-force-feeding continuing through the force-feeding period (flock C).
Pattern 3: no shedding detected at any time (flock G).
The relationship between prevalence and age was analyzed using a GAMM model incorporating a smooth trend for age and a random effect for flock. Significant effects of age and flock were observed (P < 0.05) for flocks A, B, D, E and F. For each flock, data and predicted confidence intervals were plotted in Fig. 2, showing an increasing level of prevalence when ducks were in an open environment or a particular age period (after 8 weeks of age), with a random effect for the flock. The relationship between excretion level in positive samples and age was also significant (P < 0.005) (data not shown).
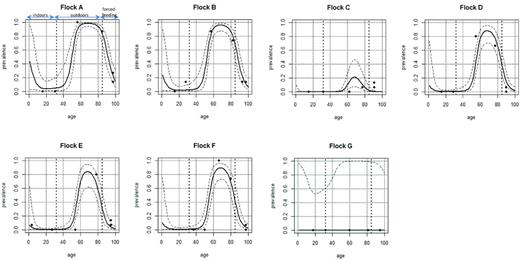
Data and predicted level of shedding in mule duck flocks investigated (A to G) fitted using the GAMM model.
Positive samples were all re-analyzed with a C. psittaci RT-PCR. Whereas for two heavily shedding flocks (flocks A and E), most of the positive samples were confirmed as positive with the C. psittaci RT-PCR, only weak PCR signals were obtained for four flocks (B, C, D and F) from a very limited number of previously confirmed Chlamydiaceae-positive samples. Sequencing the ompA gene for some C. psittaci-negative samples of these flocks confirmed the presence of C. psittaci strains belonging to the GR9 genotype C reference strain.
Whenever possible, at least three samples from each sampling session in each flock were subject to ompA sequencing. Detailed results are given in Fig. S1 (Supporting Information). The sequenced samples from a given flock were always identical. Two main C. psittaci genotypes were identified in these flocks: E/B and C. A sequence comparison identified three different E/B subgenotypes and two different C subgenotypes. Indeed, whereas an E/B subgenotype close to 06-859 was identified, a new sequence was obtained, similar to those identified above in a breeder flock (Fig. S1A, Supporting Information). One C subgenotype was strictly identical to GR9 in flocks B and C, whereas in flocks D and E, a single mutation was detected in the 1105 bp compared to the GR9 ompA sequence (Fig. S1B, Supporting Information).
Investigation of human cases linked to female breeding flocks
In late 2009, four professional workers (cases 2, 4, 5 and 6) in contact with ducks, eggs and/or ducklings were admitted to hospital reporting atypical pneumonia and/or signs of respiratory distress. There was a report of a further two suspect cases (cases n°1 and 3) with flu-like symptoms but who were not hospitalized. Whereas throat samples and the first blood sample collected after hospitalization (1–7 days after the first clinical signs) were all negative for C. psittaci, the second blood sample—taken 16–41 days after the first one—was positive for all hospitalized cases, with at least a 4-fold rise in the antibody titer (Table 3A). As no laboratory confirmation or biological samples (serum or throat swab) were available for cases n°1 and 3, they were considered as possible cases.
Cases 2 and 4 worked in close contact with female Pekin ducks on two different farms (farms 1 and 3) and had cleaning and/or egg collection activities. Cases 5 and 6 worked in a hatchery where eggs from different farms (including farms 1 to 3, Table 3B) were collected daily and incubated. Their activities were linked to ducklings or cleaning.
Once the first cases of human psittacosis were suspected on these three farms (farms 1 to 3), birds were immediately sampled (Table 3B). On farm 1 (linked to human case 2), two different female Pekin flocks were bred separately, whereas on farms 2 and 3 (linked to cases 3 and 4, respectively), only one female Pekin flock was present. All the flocks present on these three farms were sampled and examined. These four different laying flocks had in common a bi-weekly insemination with semen from a single male flock (farm 0). Cloacal shedding was detected by PCR in both the four female flocks and the male flock (Table 3B). Many birds in these flocks were positive for Chlamydiaceae without showing any clinical signs. Biological samples were genotyped and all ompA sequences were found to be identical to those of the C. psittaci 06-859 strain (data not shown).
When Chlamydiaceae were evidenced in these female flocks, investigations were then conducted on randomly selected eggs still being incubated in the hatchery and then on ducklings, all from one of the investigated female flocks (flock 2009141, farm 1). Eggs laid in week 44 (26–30/10/09) (Fig. 2), corresponding to the week when three human cases were reported, were analyzed. Dead embryos (in the last week before hatching) and 1-day old ducklings were necropsied. Among the 20 necropsied embryos, two were Siamese and thus excluded from the analysis. One of these 18 embryos was highly positive (liver, lung and intestines) by RT-PCR, two others having only a minor infection as only one organ was positive and with a low bacterial load (Table 3B). One 1-day old bird in this hatching group was positive with a low bacterial load in the spleen (Table 3B).
After hatching, 1-day old ducklings are transferred to farms. The offspring of flock 2009141 was investigated on a farm. Cloacal swabs were taken from 4-week old ducklings obtained from eggs laid either in week 42 or 44 (parent flock 2009141), kept on two different farms (Table 3B). No birds from the batch laid in week 42 tested positive, whereas all the birds from the batch laid in week 44 shed Chlamydiaceae. Positive samples from breeders and offspring eggs/birds were genotyped and all ompA sequences were identical to those of the C. psittaci 06-859 strain (data not shown).
DISCUSSION
Most human cases of psittacosis reported in France are linked to occupational exposure to ducks. This study aimed to describe natural C. psittaci infection during the mule duck production process, from the breeding stage to the force-feeding period, to refine our knowledge about this disease of public health interest.
Random investigations conducted over one year in four laying flocks dedicated to the production of mule duck eggs showed that Chlamydiaceae are intermittently shed in small quantities by breeder birds. However, heavy C. psittaci shedding in laying flocks was identified after the suspicion of human psittacosis cases on different farms. Such ‘highly infected’ breeder flocks have been reported in the past (Guérin et al., 2006). For unknown reasons, heavy shedding may occur in asymptomatic C. psittaci carrier birds that are only suspected once human infections have occurred. Co-infections with other bacteria or viruses have been shown to exacerbate the pathogenicity of C. psittaci in turkeys (Van Loock et al., 2005, 2006). Unfortunately, no complementary analysis was conducted on the breeder ducks studied which were treated with antibiotics after sampling. Interestingly, all the infected but asymptomatic females from these different breeding farms were inseminated with semen from the same male flock, which was also diagnosed as a heavily shedding flock, suggesting possible venereal transmission as already reported for ruminants (Teankum et al., 2007). The progeny of one of these infected flocks proved to be infected and the same C. psittaci genotype was identified in parents and in infected eggs or ducklings. Even if considered rare, the vertical transmission of C. psittaci has already been described in both chickens and turkeys (Wittenbrink, Mrozek and Bisping 1993; Vanrompay et al., 1997; Lublin et al., 1996). Up to now, few chlamydial isolations have been reported from the eggs or embryos of chickens, ducks, parakeets, sea gulls or snow geese (Vanrompay et al., 1997). Experimental infections conducted on chicken embryos have shown that the inoculum size has an impact on the survival rate of the infected embryos (Braukmann et al., 2012). Only small quantities of bacteria should therefore be transmitted to live ducklings.
As previously suggested (Vanrompay et al., 1997), vertical transmission is probably not the main explanation for the persistence of infected flocks on farms. In our survey on mule duck flocks, heavy shedding was observed with open range birds when tested at 8 weeks old. Recent investigations conducted in chicken flocks, where the first Chlamydia gallinacea strains were isolated, suggested that the environment is probably involved in Chlamydiae transmission as C. gallinacea strains with different ompA sequences were isolated from each investigated chicken flock linked by a single parent flock and bred on separate farms (Laroucau et al., 2009b). In the present study, distinct C. psittaci genotypes (E/B and C genotypes) were also identified in two mule duck flocks originating from the same parent flock. These findings suggest environmental/horizontal transmission rather than vertical transmission, especially as ompA sequencing of positive samples from these two flocks resulted in the identification of a single genotype in each, disproving the hypothesis of the initial presence of various genotypes from the parent flock. Interestingly, the only C. psittaci-free mule duck flock identified in the present study was housed in a new building and had access to an outdoor run never used by other domestic animals before. Even though infection by inhalation has already been reported (reviewed in Harkinezhad, Geens and Vanrompay 2009), transmission by the fecal-oral route is the most probable main pathway in duck flocks because the natural site of infection described in most animals, including birds, is the gastrointestinal tract, in which the bacterium is able to persist indefinitely in a commensal relationship (reviewed in Rank and Yeruva 2014). The experimental oral inoculation of a C. psittaci strain (06-859 genotype) in mule ducklings led to a major proliferation of the bacteria in the large intestinal tract, without any clinical manifestation (Thierry et al., 2013).
Intra-flock contamination implies the persistence of the bacteria in the environment. Cleaning and disinfection procedures as well as C. psittaci's survival capacity in the environment must have an important impact on their persistence. Chlamydiae are environmentally labile but can remain infectious in organic material (litter and feces) for many months (Longbottom and Coulter 2003). Given the attractiveness of wetlands to ducks and the occurrence of free-living amoebae in water sources, it would also be beneficial to study the potential role of amoebae as additional reservoirs for C. psittaci in the environment, as previously suggested (Essig et al., 1997).
On farms, it is probable that infected ducklings from heavily shedding laying flocks prematurely shed Chlamydiae and introduced the bacteria onto the farm by contaminating their environment (both indoors and outdoors). Uninfected ducklings arriving at these contaminated farms could then find surviving Chlamydiae in their environment. This contamination could occur at any time and could explain the different shedding patterns observed, probably depending on the cleaning and disinfection procedures implemented in addition to the birds’ immune status. Waves of infection have been described in C. psittaci-infected turkey flocks (Vanrompay et al., 1997). Once the bacteria have been introduced onto a farm, the infection is probably maintained by a fecal-oral cycle of transmission within flocks. Experimental models are needed in order to test this hypothesis and better understand chlamydial transmission and dissemination in duck flocks. Efficient control strategies can only be established once all possible infection pathways and means of C. psittaci persistence have been identified.
In 2013, the world production of foie gras from ducks and geese was estimated at 26 million tons, 71% of which was produced in France (corresponding to 36 million mule ducks bred each year). Throughout the French territory, the foie gras industry comprises about 20 hatcheries, more than 5000 producers involved in the breeding and force-feeding of the mule ducks and about 20 slaughterhouses (Anonymous 2014). Based on our survey, it appears that mule ducks mostly shed C. psittaci when they are outdoors on grass and when human contact is relatively limited. Duck-linked psittacosis cases are regularly reported in France (http://www.invs.sante.fr/surveillance/psittacose/default.htm) without being linked to a specific stage of the mule duck breeding process. Indeed, human infections occur at various steps of the breeding process: egg incubation/hatching, breeding period, bird transfer, force-feeding period and also at the slaughterhouse. Cases of human infection appear to be linked not only to exposure to a heavy bacterial load but also to the strain's virulence and the human's receptivity. Contaminated dust is known to play an important role in human exposure. Based on observations made in this study, except for the period when birds are allowed to live outdoors, the exposure of professionals to C. psittaci appears low, at least during the force-feeding period. But human cases are nevertheless reported in this workplace and in the slaughterhouses. At least one other shedding pattern, including heavy shedding in young birds from an infected parent flock, was observed in this study, supporting a previous study (Laroucau et al., 2009a). Other shedding schemes, different from those reported in the present study, could exist. Treatment with antibiotics could also stop, delay or render undetectable any shedding. Without any clinical signs observed in infected birds, their infectious status is not predictable, so it is highly recommended that people in contact with duck flocks wear protective clothes, including a mask, when handling birds during vaccinations, bird transfers, etc. A lot of dust is raised during these activities, which could be a source of aerial contamination. Air samples from these specific places should be analyzed in order to quantify the risk of exposure to humans, as previously carried out in Belgian hatcheries and slaughterhouses (Dickx et al., 2010; Dickx and Vanrompay 2011).
Psittacosis was immediately suspected in duck production workers at the onset of clinical signs because the professional organization had been made aware of the possibility of transmission. Therefore, antibiotic treatment was given quickly, explaining the negative PCR results for biological samples collected from hospitalized patients. However, medical staff are not always informed of their patients’ exposure to birds, and the consequences of a C. psittaci infection can be extremely serious (Carlier et al., 2014) or even fatal (Petrovay and Balla 2008; Belchior et al., 2010). Owing to non-specific clinical signs, the early identification of human psittacosis cases remains a challenge and most of human psittacosis cases are probably underdiagnosed.
In conclusion, enteric chlamydial carriage of C. psittaci in French mule ducks is frequent and subclinical under field conditions. While the vertical transmission of C. psittaci has been observed in the past and is supported by this study, it does not appear to be the only transmission pathway between flocks. Horizontal and/or environmental transmission is more probable. Our results highlight the widespread nature and importance of avian chlamydiosis in duck flocks and the urgent need for better knowledge and tools allowing the asymptomatic carriage of C. psittaci in birds to be reduced for the good of public health. Avian chlamydiosis on duck farms is a hidden infection that it is vital for professionals to take seriously.
This work was supported by Afssa (transversalité ChlamCan) and the Vendée Regional Council. We are grateful to Drs Virginie Mick, Danijela Horvatek and Shivaprasad for their valuable advice during the preparation of the manuscript.
Conflict of interest statement. None of the authors has a financial or personal relationship with any other people or organizations that could inappropriately influence or bias this paper.
REFERENCES

{kind=link}
{kind=link}