





Abstract
In southeastern United States farmscapes, corn, peanut, and cotton fields are often closely associated with each other. Thus, this 5-yr study was conducted to determine parasitism and predation of sentinel and natural occurring Euschistus servus (Say) and Nezara viridula (L.) egg masses in farmscapes in Georgia. A variety of parasitoids including six scelionids, Trissolcus basalis (Wollaston), T. brochymenae (Ashmead), T. euschisti (Ashmead), T. thyantae Ashmead, Telenomus podisi Ashmead, and Gyron obesum Masner, and one encyrtid, Ooencyrtus spp., was recovered from E. servus and N. viridula eggs, but overall the greatest diversity of parasitoids emerged from stink bug eggs in corn. Only T. basalis and T. podisi parasitized both N. viridula and E. servus eggs in all three crops over all farmscapes. A wide variety of predators, including Orius insidiosus (Say), Geocoris punctipes (Say), G. uliginosus (Say), Coccinella septempunctata (L.), Hippodamia convergens Guérin-Méneville, Harmonia axyridis (Pallas), Coleomegilla maculata (De Geer), Scymnus loewii Mulsant, Scymnus ardelio Horn, and Solenopsis invicta Buren, preyed on stink bug eggs in each crop. When differences were detected, generally percent parasitization per egg mass was higher than predation in corn, and percent predation was higher than parasitization in peanut. Overall, percent predation per egg mass was higher than parasitization in cotton, but on one occasion percent parasitization was higher than predation in cotton associated with corn. Seasonal occurrence and abundance of natural enemies, levels of parasitization and predation of stink bug eggs, and percent egg predation by sucking and chewing predators indicated that natural enemies of stink bug eggs disperses from crop to crop in the farmscapes. Stink bug egg predators and parasitoids are likely responding to host plant switching by their prey or hosts. In conclusion, this study has shown that natural enemies of stink bugs are prevalent and important biological control agents in these farmscapes.
In the southeastern United States, the southern green stink bug, Nezara viridula (L.), and the brown stink bug, Euschistus servus (Say), are predominant species of phytophagous stink bugs in a variety of crops. In Georgia, corn is one of the first cultivated host plants available to stink bugs (Todd and Herzog 1980, Tillman 2010a). Generally, peanut is the next cultivated host plant of stink bugs (Tillman 2008). Soybean and cotton are mid-to-late-season cultivated host plants for stink bugs (Todd and Herzog 1980, Bundy and McPherson 2000, Tillman et al. 2009). In farmscapes in this region, these four crops are often closely associated with each other, and thus stink bugs in the first available crop may disperse into the next available crop(s). For example, in peanut-cotton farmscapes, N. viridula and E. servus develop in peanut and then switch to feeding on cotton especially at the common boundary of the two crops (Tillman et al. 2009). In all likelihood, availability of suitable food in time and space and farmscape structure drive the spatiotemporal distribution and abundance of stink bugs in these farmscapes. Similarly, stink bugs worldwide use sequences of host plants in a variety of cropping systems for food throughout the year (Toscano and Stern 1976, Jones and Sullivan 1982, Velasco and Walter 1992, Ehler 2000).
Assessment of the natural biological control of N. viridula and E. servus eggs at the farmscape level is essential in strategic placement and design of biological control strategies for management of these pests. Even though the sequence of host plants has been determined for stink bugs in various farmscapes world-wide, as mentioned above, and stink bug egg parasitization and predation have been assessed for some individual crops (Yeargan 1979; Buschman and Whitcomb 1980; Ragsdale et al. 1981; Krispyn and Todd 1982; Orr et al. 1986; Stam et al. 1987; Shepard et al. 1994; Corrêa-Ferreira and Moscardi 1995; Jones 1995; van den Berg et al. 1995; Jones et al. 1996; Ehler 2002; Tillman 2008, 2010b; Koppel et al. 2009), the natural biological control of stink bug eggs has not been studied in any farmscapes except for the study conducted by Ehler (2000). Thus, this study was conducted to examine predation and parasitism of N. viridula and E. servus eggs in some existing farmscapes in Georgia.
Materials and Methods
Study Sites.
This study was conducted in corn-cotton, corn-peanut, peanut-cotton, and corn-peanut-cotton farmscapes located in Irwin Co. in southwestern Georgia. Corn, peanut, and cotton fields ranged in size from 5 to 25 ha. All recommended agricultural practices for production of corn (Lee et al. 2010), peanut (UGA 2010), and cotton (Collins et al. 2010) were followed including planting all crops at a row width of 0.91 m. Insecticides were not applied to fields for management of stink bugs during the tests.
Sentinel Stink Bug Egg Masses.
To assess parasitism and predation of eggs of E. servus and N. viridula in these farmscapes, egg masses of the two pests were placed as sentinels in crops at the interface of the farmscapes as described by Tillman (2010b). Egg masses were obtained in the laboratory from field-collected females as described by Tillman (2008). The number of egg masses per pest species per crop per field is listed on tables with mortality data. Egg masses recovered from the field were brought into the laboratory, examined for predation of individual eggs, and then, if appropriate, held for emergence of adult parasitoids. An egg that had been fed on by a sucking predator was recognizable because the inside of the egg was dry even though the egg shell was intact, and the tiny feeding hole(s) made by predator was visible on the egg shell. An egg that had been fed on by a chewing predator was recognized by the remaining bits of egg shell still attached to the cloth or by the absence of the egg. Percent predation was determined for sucking and chewing predators.
Naturally Occurring Stink Bug Egg Masses and Predators.
To determine occurrence of parasitoid species emerging from naturally occurring stink bug eggs and occurrence and abundance of stink bug egg predators, crops in these farmscapes were examined weekly throughout the field for stink bug egg masses, predators, or both, during the growing season. The number of samples per week per crop per field obtained for naturally occurring stink bug egg masses and stink bug predators in these farmscapes is listed in Table 1. For sampling purposes, the aerial parts of all corn and cotton plants within 1.83 m of row were visually checked thoroughly for all insects of interest. In these crops, predator species were identified and recorded in the field. Peanut was sampled by sweeping 7.31-m of row using a sweep net (38 cm diameter). Once a sweep sample was collected, it was put in a 3.8 liter self-sealing plastic bag that was then placed in a cooler and transported to the laboratory where predator species were identified and recorded. All egg masses found during sampling of these crops were held in the laboratory for emergence of adult parasitoids. Voucher specimens of all insects are stored in the USDA-ARS Crop Protection and Management Research Laboratory in Tifton, GA.
Number of samples obtained per week per crop per field for naturally occurring stink bug egg masses and stink bug predators in farmscapes during 5-yr study
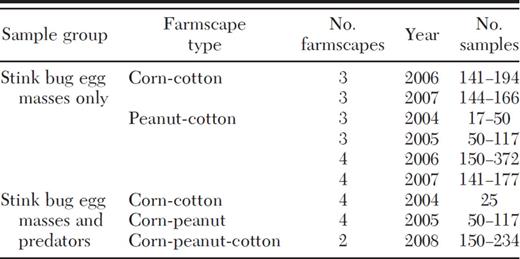
Number of samples obtained per week per crop per field for naturally occurring stink bug egg masses and stink bug predators in farmscapes during 5-yr study
Data Analysis.
Overall occurrence of parasitization by each specific parasitoid species or combination of parasitoid species was calculated for each stink bug species in each crop in each farmscape. Next, least squares means for percent parasitization and predation per egg mass for sentinel egg masses of N. viridula and E. servus were obtained for each date for each crop in a farmscape using the PROC MIXED procedure (SAS Institute 2003). In preliminary analyses, the fixed effects were week, crop, mortality type, week × crop, week × mortality type, crop × mortality type, week × crop × mortality type. Random effects were field and residual error. Where preliminary analyses revealed that there was a significant week, crop, and mortality type interaction, fixed effects were changed to week × crop × mortality type. Least squares means were separated by least significant difference (LSD) (SAS Institute 2003) where appropriate. Mean percent parasitization and predation per egg mass were determined for sentinel egg masses in individual crops in a farmscape using PROC MEANS (SAS Institute 2003).
Seasonal graphs of natural enemies were constructed for crops in all farmscapes except for peanut-cotton farmscapes. Means per crop per sampling date were obtained for predators using PROC MEANS (SAS Institute 2003). For seasonal graphs of Orius insidiosus (Say) in corn-peanut farmscapes in 2005, density in corn was divided by 10 to better visualize relationships between predators. For seasonal graphs of Solenopsis invicta Buren, density in peanut was multiplied by 10.
Results
Stink Bug Egg Parasitoids.
Stink bug egg parasitoids associated with N. viridula and E. servus in corn, peanut, and cotton in corn-cotton, corn-peanut, peanut-cotton, and corn-peanut-cotton farmscapes in Georgia consisted of seven species: six scelionids, Trissolcus basalis (Wollaston), T. brochymenae (Ashmead), T. euschisti (Ashmead), T. thyantae Ashmead, Telenomus podisi Ashmead, and Gyron obesum Masner, and one encyrtid, Ooencyrtus spp. (Table 2). Only T. basalis and T. podisi parasitized both N. viridula and E. servus eggs in all three crops over all farmscapes. Generally, T. basalis was the most prevalent parasitoid of N. viridula while T. podisi was the predominant parasitoid of E. servus. When multiparasitism, or parasitism of an individual egg mass by two parasitoid species, occurred, it always involved one of these predominant species. T. brochymenae parasitized eggs of both E. servus and N. viridula at very low levels, but only in corn farmscapes. A combination of T. brochymenae and T. podisi was recovered from an E. servus egg mass in corn. T. euschisti, either alone or in combination with T. podisi, was recovered only from E. servus eggs in both crops in corn-cotton farmscapes while T. thyantae was recovered from E. servus and N. viridula eggs in corn and peanut in corn-peanut and corn-peanut-cotton farmscapes. Multiparasitism of an E. servus egg by T. thyantae in combination with T. podisi or T. basalis occurred in corn in corn-peanut farmscapes. G. obesum was recovered only from N. viridula eggs in each crop in corn-peanut-cotton farmscapes, but only in combination with T. basalis. This was the only case of multiparasitism of N. viridula eggs observed in this study. An encyrtid, Ooencyrtus spp., was recovered from N. viridula eggs in corn and peanut and E. servus in corn and cotton.
Occurrence of parasitization of egg masses by T. basalis (1), T. brochymenae (2), T. euschisti (3), T. thyantae (4), T. podisi (5), Ooencyrtus spp. (6), and G. obesum (7) for E. servus (ES) and N. viridula (NV) in corn (CO), peanut (PE), and cotton (CT) in corn-cotton, corn-peanut, peanut-cotton, and corn-cotton-peanut farmscapes

Total no. of parasitized egg masses from which adult parasitoids emerged.
Both parasitoid species emerged from each egg mass.
Includes naturally occurring eggs collected in corn-cotton and peanut-cotton farmscapes in 2006 and 2007.
Occurrence of parasitization of egg masses by T. basalis (1), T. brochymenae (2), T. euschisti (3), T. thyantae (4), T. podisi (5), Ooencyrtus spp. (6), and G. obesum (7) for E. servus (ES) and N. viridula (NV) in corn (CO), peanut (PE), and cotton (CT) in corn-cotton, corn-peanut, peanut-cotton, and corn-cotton-peanut farmscapes
Total no. of parasitized egg masses from which adult parasitoids emerged.
Both parasitoid species emerged from each egg mass.
Includes naturally occurring eggs collected in corn-cotton and peanut-cotton farmscapes in 2006 and 2007.
Stink Bug Egg Predators.
A wide variety of predators preyed on N. viridula and E. servus egg masses in each crop in corn-cotton, corn-peanut, peanut-cotton, and corn-peanut-cotton farmscapes. Sucking predators that were observed feeding on stink bug eggs included O. insidiosus, Geocoris punctipes (Say), and G. uliginosus (Say). Chewing predators that were observed feeding on stink bug eggs included fire ants, S. invicta, larvae of lady beetles, including Coccinella septempunctata (L.), Hippodamia convergens Guérin-Méneville, Harmonia axyridis (Pallas), Coleomegilla maculata (De Geer), Scymnus loewii Mulsant, and S. ardelio Horn, and unknown species of grasshoppers.
Corn-Cotton Farmscapes.
For E. servus sentinel egg masses, factorial analyses revealed that there was no significant week, crop, and mortality type effect and no significant week and crop, week and mortality type, crop and mortality type, and week, crop, and mortality type interactions in corn-cotton farmscapes in 2004 (Table 3). Percent parasitization of E. servus egg masses, though, was relatively high in corn in June, and there was a general trend toward higher parasitization per egg mass than predation in corn for both weeks sentinel egg masses were present in each crop in the farmscapes (Table 4).
F, df, and P values for E. servus and N. viridula sentinel egg masses in corn-cotton (CO-CT), corn-peanut (CO-PE), peanut-cotton (PE-CT), and corn-peanut-cotton (CO-PE-CT) farmscapes during 5-yr study
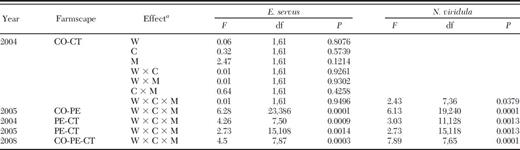
W, week; C, crop; M, mortality type.
F, df, and P values for E. servus and N. viridula sentinel egg masses in corn-cotton (CO-CT), corn-peanut (CO-PE), peanut-cotton (PE-CT), and corn-peanut-cotton (CO-PE-CT) farmscapes during 5-yr study
W, week; C, crop; M, mortality type.
Means (M) and least squares means (LSM) for percent parasitization (Para) and predation (Pred) per egg mass (EM) for sentinel egg masses of E. servus (ES) and N. viridula (NV) in corn (CO) and cotton (CT) in four corn-cotton farmscapes in 2004
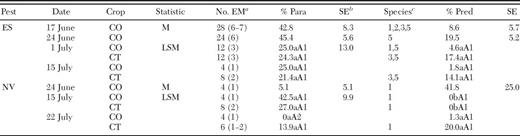
Least squares means within a row followed by the same lowercase letter are not significantly different between mortality factors (LSD, P > 0.05). Least squares means within a column followed by the same uppercase letter are not significantly different between crops for same mortality factor within the same week (LSD, P > 0.05). Least squares means within a column followed by the same no. are not significantly different between dates for the same crop and mortality factor (LSD, P > 0.05).
Total no. of stink bug egg masses (no. of stink bug egg masses per field).
SEM or common SE for least squares means.
1, T. basalts; 2, T. brochymenae; 3, T. euschisti; 5, T. podisi.
Means (M) and least squares means (LSM) for percent parasitization (Para) and predation (Pred) per egg mass (EM) for sentinel egg masses of E. servus (ES) and N. viridula (NV) in corn (CO) and cotton (CT) in four corn-cotton farmscapes in 2004
Least squares means within a row followed by the same lowercase letter are not significantly different between mortality factors (LSD, P > 0.05). Least squares means within a column followed by the same uppercase letter are not significantly different between crops for same mortality factor within the same week (LSD, P > 0.05). Least squares means within a column followed by the same no. are not significantly different between dates for the same crop and mortality factor (LSD, P > 0.05).
Total no. of stink bug egg masses (no. of stink bug egg masses per field).
SEM or common SE for least squares means.
1, T. basalts; 2, T. brochymenae; 3, T. euschisti; 5, T. podisi.
For N. viridula sentinel egg masses, the interaction between week, crop, and mortality type was significant in corn-cotton farmscapes in 2004 (Table 3). On 15 July, percent parasitization of an N. viridula egg mass was moderately high in corn, and percent parasitization per egg mass was significantly higher than percent predation for both crops (Table 4). The next week no N. viridula egg masses were parasitized in corn probably because the crop was senescing, but eggs of this pest were parasitized in cotton, even though at a moderately low level.
In these corn-cotton farmscapes, percent parasitization per egg mass for E. servus decreased numerically in corn from 24 June to 1 July, and ≈24% of the egg masses in cotton were parasitized on the latter date (Table 4). T. podisi and T. euschisti parasitized E. servus eggs in corn on 17 June, and both parasitoids emerged from E. servus eggs in cotton on 1 July. T. basalis emerged from N. viridula eggs in corn on 24 June and in cotton on 15 and 22 July. Altogether, these results suggest that these parasitoids dispersed from corn into cotton in these farmscapes.
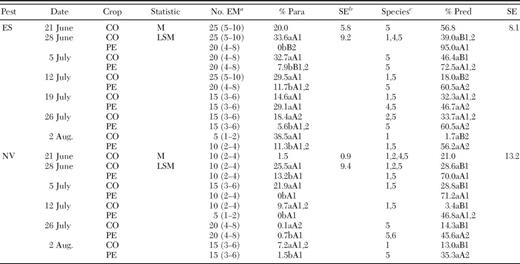
In general, stink bug predation per egg mass ranged from ≈20–40% in corn on 24 June, and afterwards egg predation decreased in corn and increased in cotton (Table 4). On 17 June, O. insidiosus density was very high in corn (Fig. 1). Afterwards, density of this predator decreased in corn and began building up in cotton. Likewise, in corn, percent egg predation by sucking predators was relatively high on 17 June and then dropped the following week. By 1 July, 40% of the stink bug eggs were fed on by sucking predators in cotton. Collectively, these results suggest that O. insidiosus dispersed from corn into cotton in these farmscapes.
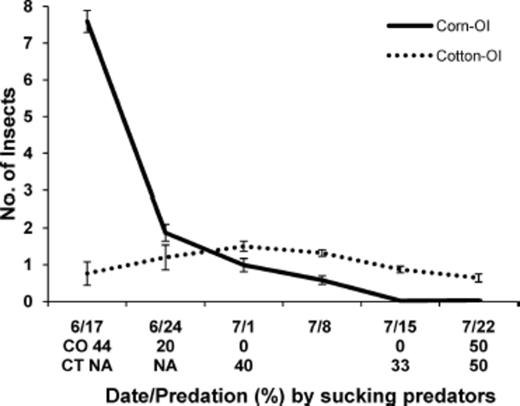
Mean (±SE) number of O. insidiosus (OI) and percent egg predation by sucking predators in corn and cotton in corn-cotton farmscapes for sampling dates in 2004.
Corn-Peanut Farmscapes.
Factorial analyses revealed that there was a significant week, crop, and mortality type interaction for sentinel egg masses of E. servus and N. viridula in corn-peanut farmscapes in 2005 (Table 3). Percent parasitization per egg mass in corn peaked at ≈40% for E. servus and at ≈25% for N. viridula (Table 5). The peak parasitization rate for egg masses of both stink bug species was almost 10% lower in peanut compared with that in corn. In addition, in corn, percent predation of egg masses was not significantly different from parasitization for both stink bugs for every date. Contrariwise, percent predation of E. servus egg masses in peanut was significantly higher than parasitization on 28 June, 5, 12, and 26 July and 2 August. On all but the fourth date, percent predation per egg mass was significantly higher in peanut than in corn. Conversely, percent parasitization of E. servus egg masses was significantly higher in corn than in peanut on the first two dates. Similarly, for N. viridula eggs, percent predation per egg mass was significantly higher than parasitization in peanut on each date, and it was significantly higher in peanut compared with corn on each date.
Means (M) and least squares means (LSM) for percent parasitization (Para) and predation (Pred) per egg mass (EM) for sentinel egg masses of E. servus (ES) and N. viridula (NV) in corn (CO) and peanut (PE) in four corn-peanut farmscapes in 2005
Least squares means within a row followed by the same lowercase letter are not significantly different between mortality factors (LSD, P > 0.05). Least squares means within a column followed by the same uppercase letter are not significantly different between crops for same mortality factor within the same week (LSD, P > 0.05). Least squares means within a column followed by the same no. are not significantly different among dates for same crop and mortality factor (LSD, P > 0.05).
Total no. of stink bug egg masses (no. of stink bug egg masses per field).
SEM or common SE for least squares means.
1, T. basalis; 2, T. brochymenae; 4, T. thyantae; 5, T. podisi; 6, an Ooencyrtus spp.
Means (M) and least squares means (LSM) for percent parasitization (Para) and predation (Pred) per egg mass (EM) for sentinel egg masses of E. servus (ES) and N. viridula (NV) in corn (CO) and peanut (PE) in four corn-peanut farmscapes in 2005
Least squares means within a row followed by the same lowercase letter are not significantly different between mortality factors (LSD, P > 0.05). Least squares means within a column followed by the same uppercase letter are not significantly different between crops for same mortality factor within the same week (LSD, P > 0.05). Least squares means within a column followed by the same no. are not significantly different among dates for same crop and mortality factor (LSD, P > 0.05).
Total no. of stink bug egg masses (no. of stink bug egg masses per field).
SEM or common SE for least squares means.
1, T. basalis; 2, T. brochymenae; 4, T. thyantae; 5, T. podisi; 6, an Ooencyrtus spp.
In these corn-peanut farmscapes, T. podisi parasitized E. servus eggs in corn on 21 June and subsequently were recovered from eggs of this pest in peanut every date egg masses were placed in peanut (Table 5). T. thyantae and T. basalis parasitized E. servus eggs in corn on 28 June and then in peanut on 19 July and 2 August, respectively. In addition, T. basalis and T. podisi parasitized N. viridula eggs in corn on 21 June and in peanut the following week. Apparently, these parasitoids dispersed from corn into peanut in these farmscapes.
Overall, when sentinel egg masses were present in both crops, percent predation per egg mass for stink bugs in peanut could be very high, peaking at ≈95% for E. servus and at ≈70% for N. viridula, but it was around 50% lower for each stink bug species in corn compared with peanut (Table 5). Density of O. insidiosus was relatively very high in corn in mid-to late-June, and percent egg predation by sucking predators was moderately high in corn on 14 June (Fig. 2 A). After this date, percent egg predation by sucking predators occurred in one or both crops, but at very low levels except on 2 August when percent predation by sucking predators (presumably by Geocoris spp.) was moderately high in corn. Presence and abundance of these sucking predators in corn and peanut over time indicate that they dispersed from corn into peanut. For chewing predators, percent egg predation was very high in both crops from 21 June to 26 July when lady beetle larvae were present in each crop (Fig. 2 B). Evidently, stink bug egg predation in corn and peanut can be attributed, at least in part, to these sucking and chewing predators.
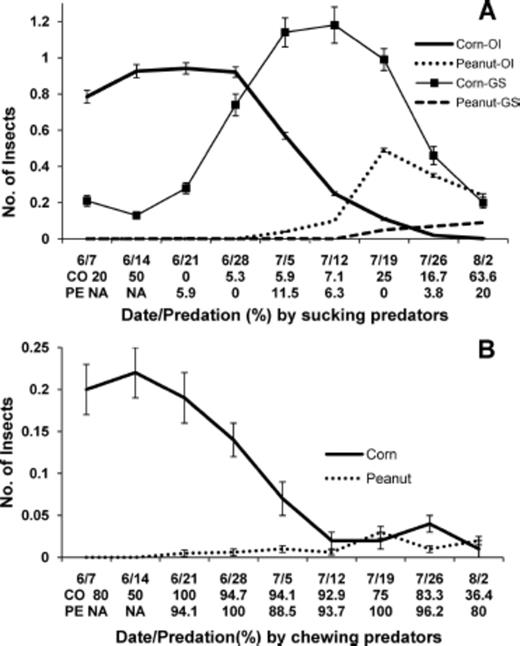
Mean (±SE) number of predators and percent egg predation by sucking or chewing predators in corn and peanut in corn-peanut farmscapes for sampling dates in 2005. (A) O. insidiosus (OI) and Geocoris spp. (GS). Density of O. insidiosus in corn was divided by 10. (B) Lady beetles (H. convergens, C. septempunctata, C. maculata, H. axyridis, S. loewii, and S. ardelio).
Peanut-Cotton Farmscapes.
For 2004 peanut-cotton farmscapes, a significant week, crop, and mortality type interaction was detected for sentinel egg masses of E. servus and N. viridula (Table 3). Percent parasitization per egg mass ranged from 0 to ≈20% for E. servus and from two to ≈13% for N. viridula (Table 6). On 25 August, percent predation per egg mass for E. servus was very high in cotton and was significantly higher than percent parasitization in this crop, and furthermore, percent egg predation was significantly higher for this pest in cotton compared with peanut. Likewise, percent predation per egg mass for N. viridula was high in cotton on this date. Predation per egg mass for N. viridula was significantly higher than parasitization in cotton on this sampling date and the proceeding one. For both pest species, there was a general trend toward higher predation per egg mass than parasitization in peanut, even though the two mortality factors were not significantly different.
Least squares means for percent parasitization (Para) and predation (Pred) per egg mass (EM) for sentinel egg masses of E. servus (ES) and N. viridula (NV) in peanut (PE) and cotton (CT) in three peanut-cotton farmscapes in 2004 and 2005

Least squares means within a row followed by the same lowercase letter are not significantly different between mortality factors (LSD, P > 0.05). Least squares means within a column followed by the same uppercase letter are not significantly different between crops for same mortality factor within the same week (LSD, P > 0.05). Least squares means within a column followed by the same no. are not significantly different among dates for same crop and mortality factor (LSD, P > 0.05).
Total no. of stink bug egg masses (no. of stink bug egg masses per field).
Common SE.
1, T. basalts; 5, T. podisi.
Least squares means for percent parasitization (Para) and predation (Pred) per egg mass (EM) for sentinel egg masses of E. servus (ES) and N. viridula (NV) in peanut (PE) and cotton (CT) in three peanut-cotton farmscapes in 2004 and 2005
Least squares means within a row followed by the same lowercase letter are not significantly different between mortality factors (LSD, P > 0.05). Least squares means within a column followed by the same uppercase letter are not significantly different between crops for same mortality factor within the same week (LSD, P > 0.05). Least squares means within a column followed by the same no. are not significantly different among dates for same crop and mortality factor (LSD, P > 0.05).
Total no. of stink bug egg masses (no. of stink bug egg masses per field).
Common SE.
1, T. basalts; 5, T. podisi.
For 2005 peanut-cotton farmscapes, the interaction between week, crop, and mortality type was significant for sentinel egg masses of E. servus and N. viridula (Table 3). Overall, parasitization per egg mass for these stink bugs was relatively low, ranging from 0 to ≈20%, and was not significantly different between crops for each stink bug species (Table 6). Generally, percent predation per egg mass for both stink bugs gradually decreased from a very high level to a moderately high level in peanut from 20 July to 10 August. On 20 July, percent predation per egg mass peaked in peanut and was significantly higher in peanut than in cotton for both pests. Percent predation per egg mass was significantly higher than percent parasitization for E. servus in peanut on 20 and 27 July and for N. viridula in both crops on 27 July and 10 August and in peanut on 3 August. Clearly, predation was a key mortality factor for eggs of both pests in both crops in these peanut-cotton farmscapes.
Corn-Peanut-Cotton Farmscapes.
Factorial analyses revealed that there was a significant week, crop, and mortality type interaction for sentinel egg masses of E. servus in corn-peanut-cotton farmscapes in 2008 (Table 3). In corn, percent parasitization per egg mass for E. servus from 29 May through 10 July was low, ranging from ≈3–30% (Table 7). By 17 July, though, percent parasitization of these egg masses in corn was very high and significantly higher than predation in this crop and significantly higher than parasitization of eggs in peanut. Generally, percent parasitization per egg mass for E. servus in peanut was moderately low, ranging from ≈0–40%. However, on 11 September, parasitization of eggs of this pest in peanut was moderately high and numerically higher than predation in this crop and significantly higher than parasitization of these eggs in cotton. On 10 July, percent predation per egg mass in peanut was significantly higher than parasitization in this crop and significantly higher in peanut than in cotton. Generally, percent egg parasitization in cotton was low, ranging from ≈0–15%. On the last two sampling dates in September, percent predation per egg mass for E. servus was significantly higher than parasitization in cotton.
Means (M) and least squares means (LSM) for percent parasitization (Para) and predation (Pred) per egg mass (EM) for sentinel egg masses of E. servus (ES) and N. viridula (NV) in corn (CO), peanut (PE), and cotton (CT) in two corn-peanut-cotton farmscapes in 2008
Least squares means within a row followed by the same lowercase letter are not significantly different between mortality factors (LSD, P > 0.05). Least squares means within a column followed by the same uppercase letter are not significantly different between crops for same mortality factor within the same week or combined weeks (LSD, P > 0.05). Least squares means within a column followed by the same no. are not significantly different among dates for same crop and mortality factor (LSD, P > 0.05).
Total no. of stink bug egg masses (no. of stink bug egg masses per field).
SEM or common SE for least squares means.
1, T. basalts; 2, T. brochymenae; 4, T. thyantae; 5, T. podisi; 7, G. obesum.
Dates: 28 Aug., 4 Sept., 11 Sept., and 18 Sept.
Means (M) and least squares means (LSM) for percent parasitization (Para) and predation (Pred) per egg mass (EM) for sentinel egg masses of E. servus (ES) and N. viridula (NV) in corn (CO), peanut (PE), and cotton (CT) in two corn-peanut-cotton farmscapes in 2008
Least squares means within a row followed by the same lowercase letter are not significantly different between mortality factors (LSD, P > 0.05). Least squares means within a column followed by the same uppercase letter are not significantly different between crops for same mortality factor within the same week or combined weeks (LSD, P > 0.05). Least squares means within a column followed by the same no. are not significantly different among dates for same crop and mortality factor (LSD, P > 0.05).
Total no. of stink bug egg masses (no. of stink bug egg masses per field).
SEM or common SE for least squares means.
1, T. basalts; 2, T. brochymenae; 4, T. thyantae; 5, T. podisi; 7, G. obesum.
Dates: 28 Aug., 4 Sept., 11 Sept., and 18 Sept.
E. servus eggs were parasitized by T. podisi and T. basalis in early season corn and subsequently in peanut on 10 July (Table 7). In addition, T. thyantae parasitized eggs of this pest in corn in July and emerged from E. servus eggs in peanut later in September. Eggs of E. servus were parasitized by T. podisi consistently in peanut, and then this wasp parasitized eggs of this pest in cotton. Altogether, these results indicate that T. podisi, T. basalis, and T. thyantae dispersed from corn into peanut, and T. podisi dispersed from peanut into cotton.
A significant week, crop, and mortality type interaction was detected for sentinel egg masses of N. viridula in these farmscapes (Table 3). Percent parasitization per egg mass for this pest was very low, ranging from ≈0–8%, in each crop in these farmscapes over the season (Table 7). Nevertheless, predation of N. viridula eggs was significantly higher than parasitization in peanut on 10 and 17 July and in cotton on the latter date and significantly higher for peanut compared with corn on both dates. Percent predation per egg mass for N. viridula was significantly higher than parasitization in both peanut and cotton the last 4 wk of the study.
N. viridula eggs were parasitized by T. basalis, alone and in combination with G. obesum, in early season corn (Table 7). Then, T. basalis parasitized N. viridula eggs in peanut on 10 July. Later in September, eggs of N. viridula were parasitized by T. basalis–G. obesum in peanut and cotton. Collectively, these results indicate that T. basalis and G. obesum dispersed from corn into peanut, and then from peanut into cotton.
Predation of E. servus and N. viridula egg masses reached relatively high levels in corn, then in peanut, and lastly in cotton in these farmscapes (Table 7). Percent predation per egg mass for E. servus was very high in corn on 29 May. Afterwards, predation of E. servus egg masses gradually declined in corn until 10 July when predation was high in peanut. From 10 and 17 July, predation of N. viridula egg masses increased in peanut from 50 to 98% as corn was drying out. In August, percent predation of E. servus egg masses declined in peanut. Later in September, predation of E. servus egg masses was moderately high in cotton. These results suggest that stink bug egg predators dispersed from corn into peanut and then into cotton.
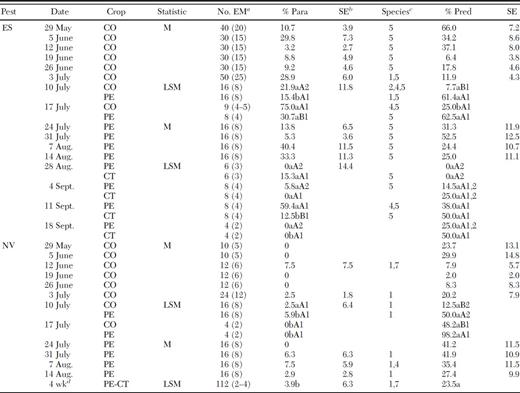
Seasonal occurrence and abundance were determined for the sucking predators, O. insidiosus and Geocoris spp., for each crop in these farmscapes. Density of O. insidiosus peaked on 29 May and was relatively high in corn through 26 June (Fig. 3 A). By 3 July, density of this predator had declined in corn, and the predator was found for the first time in peanut. Density of O. insidiosus began increasing in peanut on 10 and 17 July peaking on 31 July. As density of this predator decreased in peanut in mid-to-late-August, it was observed in both peanut and cotton. The population of this predator then peaked in cotton on 21 August. Density of Geocoris spp. also was relatively high in corn and peaked on 10 July (Fig. 3 B). Geocoris spp. were present in peanut at relatively low levels as early as 3 July but began increasing in this crop after declining in corn on 31 July. Density of Geocoris spp. peaked in peanut on 14 August. Geocoris spp. were present in cotton for the first time on 7 August, and they built up in cotton as their numbers declined in peanut. These results suggest that O. insidiosus and Geocoris spp. dispersed from crop to crop throughout the seasonal succession of crops in these corn-peanut-cotton farmscapes.
Mean (±SE) number of predators and parasitoids and percent egg predation by sucking predators in corn, peanut, and cotton in corn-peanut-cotton farmscapes for sampling dates in 2008. (A) O. insidiosus (OI) and T. basalis (TB) and (B) Geocoris spp. Percent egg predation by sucking predators below date for corn, peanut, and cotton on first, second, and third line, respectively.
Predation by sucking predators was detected in early season corn when O. insidiosus and Geocoris spp. were relatively high (Fig. 3), and percent predation of E. servus stink egg masses was relatively high (Table 7). From 31 July through 14 August, O. insidiosus and Geocoris spp. peaked in mid-season peanut and predation by sucking predators was detected (Fig. 3), and predation of stink egg masses was moderately high (Table 7). On 11 September, Geocoris spp. peaked in cotton and egg predation by sucking predators was 25% (Fig. 3). Also on this date, predation of stink bug egg masses ranged from 23.5 to 50% (Table 7). These results along with observations of these predators preying on stink bug eggs in each crop demonstrate that predation by sucking predators can be attributed to these predator species.
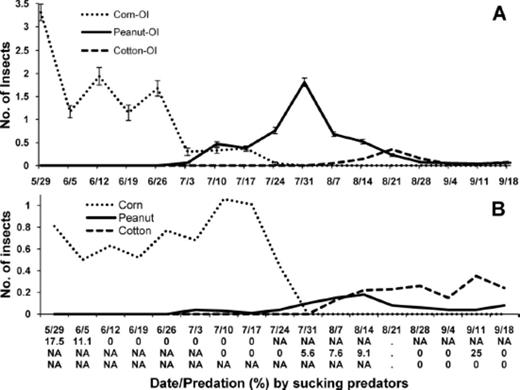
Seasonal occurrence and abundance were determined for chewing predators in each crop in these farmscapes. Density of lady beetles was relatively high in corn on three and 10 July, but these predators were not present in this crop by 17 July (Fig. 4 A). In peanut, these predators peaked on 10 and 17 July, and they peaked again in peanut, but at a lower level, on 7 August. Two weeks later lady beetles peaked in cotton and were present in peanut and cotton for the remainder of the season. These results suggest that lady beetles dispersed from corn into peanut and then into cotton. Adults of S. invicta were present in corn and peanut by 3 July, but peaked first in corn on 17 July and later in peanut on 31 July (Fig. 4 B). After peaking in peanut, density of this predator built up in cotton and remained at relatively high levels in this crop throughout the rest of the growing season. Using the occurrence of S. invicta as an indication of searching activity, these predators were actively searching for stink bug eggs in corn, then peanut, and lastly in cotton.
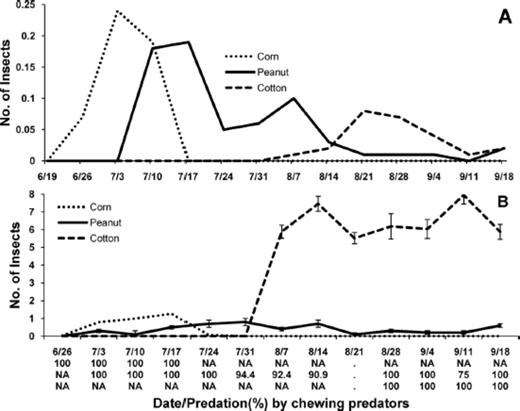
Mean (±SE) number of predators and percent egg predation by chewing predators in corn, peanut, and cotton in corn-peanut-cotton farmscapes for sampling dates in 2008. (A) Lady beetles (H. convergens, C. septempunctata, C. maculata, H. axyridis, S. loewii, and S. ardelio) and (B) S. invicta. Density of S. invicta in peanut was multiplied by 10. Percent egg predation by chewing predators below date for corn, peanut, and cotton on first, second, and third line, respectively.
From 26 June through 24 July, when either one or both groups of chewing predators were present in corn and/or peanut, stink bug eggs in corn and peanut were preyed on exclusively by chewing predators (Fig. 4). From 28 August through 18 September, both lady beetles and S. invicta were present in cotton, and egg predation by chewing predators was very high in this crop during this time. Collectively, these results indicate that egg predation by chewing predators was attributed to lady beetles and S. invicta in each crop. Indeed, both lady beetle larvae and S. invicta were observed feeding on stink bug egg masses in each crop.
Discussion
A wide variety of egg parasitoids was associated with E. servus and N. viridula eggs in corn-cotton, corn-peanut, peanut-cotton, and corn-cotton-peanut farmscapes, but overall the greatest diversity of parasitoids emerged from stink bug eggs in corn. Only T. basalis and T. podisi parasitized eggs of both stink bug species in all three crops over all farmscapes. Four additional species, T. brochymenae, T. euschisti, T. thyantae, and an Ooencyrtus spp., parasitized E. servus eggs in corn. Except for T. euschisti, these same species plus G. obesum parasitized N. viridula eggs in this crop. One other species, T. thyantae, was associated with both stink bug species in peanut, but N. viridula eggs also were parasitized by an Ooencyrtus spp. and G. obesum in this crop. Two additional species, T. euschisti and an Ooencyrtus spp., were associated with E. servus eggs in cotton while one other species, G. obesum, parasitized N. viridula eggs in this crop. For E. servus in corn, multiparasitism occurred for five different sets of parasitoid species whereas for E. servus in other crops, multiparasitism occurred for only two sets of parasitoid species. In farmscapes associated with corn, one to two additional stink bug egg parasitoids were common to two or more of the crops in the farmscape. Stink bug egg parasitoid species diversity in corn apparently enhanced species diversity in farmscapes with corn.
One possible explanation for greater egg parasitoid species diversity in corn compared with peanut and cotton is that parasitoid diversity could be high in the wide variety of early season noncrop host plants surrounding these farmscapes, and these plants may serve as sources of parasitoids for stink bug eggs in corn, one of the first crops available to these pests in these farmscapes. A preliminary study on parasitization of E. servus sentinel egg masses in surrounding vegetation, including wild black cherry, poke salad, chinaberry, oaks, pines, and wild grapevines, has shown that a variety of species, including T. podisi, T. brochymenae, an unknown encyrtid, and an unknown eulophid, parasitize eggs of this pest (P.G. Tillman, unpublished data).
When differences were detected, generally parasitization of stink bug egg masses was higher than predation in corn. Apparently, the greater egg parasitoid diversity observed in corn can result in an increase in parasitization of stink bug eggs in this crop. On many occasions, percent predation was higher than parasitization in peanut in all farmscapes. Therefore, the increase in parasitization of stink bug eggs in corn does not appear to result in higher rates of parasitization of stink bug eggs in peanut in corn-peanut farmscapes. Apparently, predation of stink bug eggs is so intense in peanut that regardless of the crop preceding peanut, predation will be high in peanut. Overall, percent predation per egg mass was higher than parasitization in cotton even though on one occasion percent parasitization was higher than predation in cotton associated with corn. Therefore, the increase in parasitization of stink bug eggs in corn is not always, but can sometimes be reflected in increased parasitization of these eggs in succeeding cotton.
Our findings are similar to previous reports on parasitization of N. viridula and E. servus by T. basalis and T. podisi in a variety of individual agronomic crops, including alfalfa, corn, soybean, peanut, and wheat, and vegetable crops (Buschman and Whitcomb 1980; Yeargan 1979; Orr et al. 1986; Corrêa-Ferreira and Moscardi 1995; Jones et al. 1996; Shepard et al. 1994; Ehler 2002; Tillman 2008, 2010b; Koppel et al. 2009). In a few instances in the current study, though, species predominance did not appear to follow the conventional pattern. For example, in 2005 T. podisi, not T. basalis was the predominant parasitoid emerging from N. viridula eggs in peanut. The reason for this difference in parasitoid species predominance is unknown, but it appears to be unrelated to farmscape composition. In addition, in 2004, frequency of occurrence of T. podisi from E. servus was relatively low in both crops in corn-cotton farmscapes, but when multiparasitism involving this T. podisi was also considered, occurrence of parasitization by T. podisi for eggs of this pest fell within expected levels.
Regarding the remaining egg parasitoids, T. brochymenae has been reported to parasitize E. servus eggs not only in corn (Tillman 2010b), but also in peanut (Tillman 2008). N. viridula has been reported as a host of this parasitoid in corn (Tillman 2010b) and soybean (Corrêa-Ferreira and Moscardi 1995). Both T. euschisti and T. thyantae have been reported to parasitize Euschistus spp. and/or N. viridula eggs in corn, soybean, and peanut (Yeargan 1979, Jones et al. 1996, Tillman 2008, 2010b). An Ooencyrtus spp. has been known to emerge from E. servus eggs in peanut (Tillman 2008) and eggs of both pests in corn (Tillman 2010b). Species of Ooencyrtus have been reported as parasitoids of eggs of both stink bug species in a variety of crops, including soybean, tomato, okra, peanut, and corn (Buschman and Whitcomb 1980; Shepard et al. 1994; Ehler 2002; Tillman 2008, 2010b). G. obesum did not emerge from E. servus and N. viridula eggs in individual corn fields (Tillman 2010b), but it has been recovered from E. servus in peanut (Tillman 2008) and soybean (Orr et al. 1986, Jones et al. 1996).
Several other studies have been conducted to determine predation of E. servus and N. viridula eggs in a variety of individual crops. The majority of these studies have been done in soybean (Yeargan 1979, Ragsdale et al. 1981, Krispyn and Todd 1982, Stam et al. 1987, Shepard et al. 1994, van den Berg et al. 1995) reflecting the significance of soybean as a major host plant of these pests. Other host plants in which stink bug egg predation have been evaluated include alfalfa (Yeargan 1979), macadamia nut orchards (Jones 1995), weeds and tomato and bean crops (Shepard et al. 1994, Ehler 2002), peanut (Tillman 2008), and corn (Tillman 2010b). Species of sucking predators that preyed on stink bug eggs included an Orius spp., an Geocoris spp., and an Podisus spp. Species of chewing predators that fed on stink bug eggs were ants, including Monomorium floricola (Jerdon), Pheidole megacephala (F.), and S. invicta, and lady beetles, including C. maculata, H. convergens, C. septempunctata, and H. axyridis.
N. viridula and E. servus are generalist feeders that exhibit edge-mediated dispersal from corn and peanut into subsequent adjacent crops in farmscapes to find food for adults and sites for oviposition and nymphal development over the growing season (Tillman et al. 2009, unpublished data). Interestingly, stink bug egg parasitoids and predators evidently disperse from crop to crop throughout the seasonal succession of crops in corn-cotton, corn-peanut, peanut-cotton, and corn-peanut-cotton farmscapes to parasitize and prey on egg masses of E. servus and N. viridula. Indeed, a rubidium mark and recapture study conducted in a corn-cotton farmscape demonstrated that O. insidiosus females dispersed from corn into cotton (Tillman et al. 2007). Considering that stink bugs of both species disperse from crop to crop to find food in farmscapes, stink bug egg parasitoids and predators are likely responding to host plant switching by their hosts or prey. Certainly, farmscape ecology of stink bugs has a significant impact on egg parasitoid species diversity and natural biological control of these pests.
The question remains how we use can this information on natural biological control of stink bug eggs in these crops to develop strategies to manage stink bugs in farmscapes. One strategy is to establish multifunctional habitats in these farmscapes to conserve and enhance natural enemies of stink bugs by providing them food sources and protection from insecticides, and by assisting in their movement through the seasonal succession of crops. Addition of nectar-producing plants in such habitats could provide season-long sources of food to natural enemies. The habitat could be strategically placed at the interface of crops within the farmscape to concentrate and enhance these natural enemies at sites were stink bugs generally congregate. This habitat could also assist natural enemies to transition from one crop to another during the period between crops, thus increasing the level and consistency of parasitization and predation in these farmscapes. Establishing the habitat in the spring before agronomic crops are present could possibly encourage species diversity of parasitoids in the farmscape by attracting and nourishing natural enemies that disperse from overwintering sites and early season host plants. This study has shown that natural enemies of stink bugs are prevalent and important biological control agents in farmscapes in which stink bugs develop, and thus an important management strategy for stink bugs in these farmscapes is the strategic addition of multifunctional habitats to conserve and enhance these natural enemies.
Thanks to Kristie Graham and Brittany Giles for technical assistance and Norman Johnson, Ohio State University, for assistance in identifying scelionid parasitoids.
References

{kind=link}
{kind=link}
{kind=link}
{kind=link}