





Abstract
Thrips exhibit different reproductive modes including thelytoky (females produced from unfertilized eggs), arrhenotoky (males produced from unfertilized eggs and females produced from fertilized eggs) and deuterotoky (females and males produced from unfertilized eggs). We investigated patterns of reproductive modes in onion thrips, Thrips tabaci Lindeman, populations and potential effects of the bacterium Wolbachia and temperature on these modes. We also examined the possibility that male-producing T. tabaci populations were resistant to the frequently used insecticides, lambda-cyhalothrin and methomyl. In New York during 2002–2004, T. tabaci populations were sampled from 20 onion fields and reproductive mode was determined by identifying sex of progeny from virgins. Half of the populations were thelytokous and half were a mix of thelytokous, arrhenotokous and deuterotokous individuals, which we refer to as a male-producing population. In two of four cases, the reproductive mode of a population from the same onion field changed across years, suggesting that populations either mix or an external factor caused the change. To address the latter, we speculated that Wolbachia or high temperature mediated reproductive modes. Samples of T. tabaci representing each reproductive mode were examined for Wolbachia using diagnostic polymerase chain reaction (PCR), but it was not detected. Cytological examination of ovaries from two additional thelytokous lines also showed no evidence of Wolbachia. Similarly, high temperature did not affect sex allocation ratios in either thelytokous or male-producing populations. Male-producing T. tabaci populations were not positively correlated with resistance to lambda-cyhalothrin, or tolerance to methomyl. The role of the different reproductive modes in T. tabaci populations in onion fields remains unclear.
Onion thrips, Thrips tabaci Lindeman, is a major pest of agricultural crops worldwide (Lewis 1997). Populations of T. tabaci can escalate quickly when conditions are hot and dry because a generation can be completed in only 2-3 wk (Cranshaw et al. 2005). The most common reproductive mode in T. tabaci populations is thelytoky (Kendall and Capinera 1990), a type of parthenogenesis in which unfertilized eggs develop into females. Thrips tabaci also are known to reproduce through arrhenotoky (Lewis 1973, Kendall and Capinera 1990, Torres-Vila et al. 1994, Gangloff 1999, Vierbergen and Ester 2000, Chatzivassiliou et al. 2002), a type of reproduction in which unfertilized eggs develop parthenogenetically into males and fertilized eggs develop into females (Lewis 1973). Arrhenotokous populations of T. tabaci are most common in the eastern Mediterranean region, where T. tabaci is thought to be indigenous, and this reproductive mode may be the ancestral form of reproduction for this species. Deuterotoky, a relatively uncommon type of parthenogenesis, occurs when unfertilized eggs develop into either males or females (Soumalainen and Saura 1993). Deuterotoky has never been reported in T. tabaci, but occurs in the thrips, Apterothrips apteris (Daniel) (Mound 1992). In New York, Gangloff (1999) observed male T. tabaci in several onion fields in a large commercial onion-producing region in Yates County. The occurrence and prevalence of male-producing T. tabaci populations in other onion-producing regions in New York is not known.
Reproductive modes in thrips and the cytological mechanisms involved in sex determination are not well understood (Moritz 1997). In T. tabaci, the production of males may be a reversion to its ancestral form, or it may be elicited by an external factor. Intracellular proteobacteria from the genus Wolbachia are known to alter reproduction in their arthropod hosts by inducing parthenogenesis, cytoplasmic incompatibility, feminization of genetic males, and male-killing (Werren 1997, Hurst et al. 1999). Although Wolbachia-induced cytoplasmic incompatibilty and male killing are likely the most common phenotype, Wolbachia-induced parthenogenesis has been described in three different Arthropod orders: Acarina, Hymenoptera, and Thysanoptera (Arakaki et al. 2001, Stouthamer et al. 1993, Weeks and Breeuwer 2001). In thrips, Wolbachia infection causes Franklinothrips vespiformis (Crawford) and Taeniothrips inconsequens (Uzel) to become thelytokous, whereas in the absence of Wolbachia they are arrhenotokous (Moritz 1997, Arakaki et al. 2001). The presence of Wolbachia in T. tabaci is not known, but it seems likely. For example, Gangloff (1999) observed male T. tabaci in onion fields, but not in nearby crops such as wheat, alfalfa, and clover. Onions contain antimicrobial properties (Hughes and Lawson 1991) that may cleanse Wolbachia from T. tabaci that use onion as its host. Therefore, it is plausible that thelytokous populations of T. tabaci are infected with Wolbachia, whereas male-producing ones are not infected.
Temperature also can affect the frequency of males encountered in some populations of thrips species, especially if temperatures are high at the time their mothers developed (Moritz 1997). Exposure to high temperature may directly affect sex allocation ratios or indirectly by reducing the susceptibility of infection by sex-altering microbes like Wolbachia. High temperatures have been shown to reduce the effects of Wolbachia, reduce Wolbachia densities or even eliminate Wolbachia infections (Hurst et al. 2000, Snook et al. 2000, Arakaki et al. 2001, Reynolds et al. 2003, Mouton et al. 2006, van Opijnen and Breeuwer 1999). The influence of temperature on parthenogenesis in T. tabaci populations has not been documented.
Thrips tabaci populations from onion fields in New York and Ontario, Canada, have proven resistant to pyrethroid and organophosphate insecticides (Gangloff 1999, Shelton et al. 2003, MacIntyre-Allen et al. 2005). T. tabaci resistance to carbamate insecticides has not been documented in New York. However, moderate to high levels of survival of T. tabaci after exposure to a low rate of methomyl in laboratory assays has been shown, suggesting that genes for resistance to carbamates are present in these populations (A.M.S., unpublished data). In an eastern Colorado onion field, Kendall and Capinera (1990) reported a positive association between insecticide resistance and an arrhenotokous population of T. tabaci. In New York onion fields, arrhenotokous populations of T. tabaci were encountered frequently in August (Gangloff 1999), when damaging populations are targeted intensively with insecticides, and less frequently at other times during the year. Perhaps, there is a tendency for arrhenotokous populations to be resistant to insecticides. If so, scouting onion fields for male T. tabaci, which are morphologically distinct from females, could be an easy and rapid method for identifying insecticide resistance.
The purpose of this project was to identify reproductive modes in T. tabaci populations in New York onion fields and to determine if either Wolbachia or temperature mediated male-producing reproductive modes. Concurrently, we wanted to determine if male-producing populations tended to be resistant to lambda-cyhalothrin and more tolerant to methomyl.
Materials and Methods
Reproductive Modes in T. tabaci Populations.
Sex determination of adult thrips was the criterion used to identify the mode of reproduction for T. tabaci populations in onion (Suomalainen and Saura 1993). Because adult T. tabaci encountered in onion fields may originate from outside these fields, thrips larvae were collected from onions and used as the test insects rather than adults. Also, because thrips may reproduce either sexually or through the several forms of parthenogenesis described earlier, the mode of reproduction used to produce a female collected from the field would be impossible to determine (i.e., a female could originate through sexual reproduction, thelytoky, or deuterotoky). Similarly, a male could have been produced through either arrhenotoky or deuterotoky. To eliminate these possibilities, the mode of parthenogenic reproduction was determined for T. tabaci by identifying the sex of progeny from virgins. To do this, T. tabaci larvae were collected from onion fields and reared in isolation to adulthood, and these virgins were permitted to reproduce parthenogenetically. If a virgin produced all females it was considered thelytokous, whereas a virgin that produced all males was considered arrhenotokous. A virgin that produced a combination of males and females was considered deuterotokous. Because we were primarily interested in comparing biological attributes between thelytokous populations and those that contained males (either arrhenotokous or deuterotokous individuals), we refer to populations that had at least one male as "male producing." No attempt was made to determine if males were reproductively functional.
Thrips tabaci were collected from a total of 20 commercial onion fields representing all major onion-producing regions in New York during 2002-2004. Dry bulb onions were grown continuously in the onion fields sampled for 5-30 yr (i.e., fields not rotated). Between late June and early October, thrips larvae were randomly sampled from 20 to 50 onion plants within a 1-ha area in each field. Larvae and the onions they infested were placed in well-ventilated plastic boxes until pupation. To ensure that test animals would be virgins, pupae were removed from the boxes and placed singly into ventilated, plastic petri dishes (5 cm in diameter). Fifty pupae were selected from each population and nearly 100% were female.
Cabbage leaf disks (3 cm in diameter) were provided as a food source and oviposition substrate for virgins. Although all cabbage leaves appeared to be thrips-free, several steps were taken to ensure that these disks were not contaminated before experimentation. To remove larvae, freshly excised disks were dipped and shaken in a sodium hypochlorite and water solution (0.25% vol:vol). This procedure also removed surface dwelling bacteria and fungi. Because eggs would not have been removed after this procedure, disks were held in tight-fitting plastic boxes and maintained at 29°C in a thrips-free environmental chamber for 1 wk to allow eggs to hatch. The procedure described above to remove larvae was repeated. Each thrips-free cabbage disk was placed in a petri dish that contained white filter paper and one thrips pupa and was sealed with parafilm. Disks supported thrips for ~1 mo (e.g., 27-39 d).
Virgins were removed from petri dishes when their progeny were mostly first instars, 7-10 d after the initiation of the experiment. Two to 3 wk later, sex of the adult offspring was determined using a dissecting microscope. For each virgin, the total number of male and female progeny was recorded. Voucher specimens are kept in a collection by the first author in the Department of Entomology, Cornell University, New York State Agricultural Experiment Station, Geneva, NY.
Temporal Variation in Reproductive Modes.
To determine if the frequency of thelytokous and male-producing populations differed among months in which populations were sampled, data from the 20 populations described above were subjected to a χ2 contingency analysis at P < 0.05 (SAS Institute 2000). Month of sampling was the independent variable and type of reproductive mode was the dependent variable. To determine if reproductive modes in a population changed across years, five nonrotated onion fields were sampled over at least 2 consecutive yr. To determine if the reproductive mode in a population changed within a season, two nonrotated onion fields were sampled twice, once in July and later in August.
Detecting Wolbachia in T. tabaci Populations.
Thrips collected from the commercial onion fields described above were preserved in ethanol and stored at -20°C until DNA was extracted. DNA from a single virgin or up to 10 progeny from a single virgin was extracted using the Puregene DNA purification kit (Gentra Systems, Minneapolis, MN). Total DNA was rehydrated in a final volume of 50 μl. Samples were tested for the presence of Wolbachia using two different sets of Wolbachia-specific primers: (1) the Wolbachia-specific primers (W-Spec), which amplify a region of the 16S rDNA gene, W-Spe cF (CATACCTATTCGAAGGGATAG) and W-Spe cR (AGCTTCGAGTGAAACCAATTC) (Werren and Windsor 2001) and (2) the Wolbachia surface protein gene (WSP) primers 81 F (TGGTCCAATAAGTGATGAAGAAAC) and 691R (AAAAATTAAACGCTACTCCA) (Zhou et al. 1998). Both sets of primers can detect Wolbachia from the major groups found in arthropods. A total of 70 thelytokous, 15 arrhenotokous, and 23 deuterotokous T. tabaci were tested. As a control for DNA template quality, polymerase chain reaction (PCR) was also performed using primers for cytochrome oxidase (COI) LCOI1490 (GGTCAACAAATCATAAAGATATTGG) and HCO2198 (GGTCAACAAATCATAAAGATATTGG) (Herbert et al. 2003). Samples from which no COI amplicons were obtained were excluded. Three different PCRs were performed concurrently on each sample using the three primer pairs. Total reaction volume was 10 μl including 1-μl template DNA with 2.5 mM MgCl2. After 2 min of denaturation at 95°C, 40 cycles of amplification were performed within 30 s at 92°C, 90 s at 55°C, and 30 s at 72°C, followed by a single 5-min extension step at 72°C. To detect potentially low levels of Wolbachia in thrips, the PCR conditions were deliberately more permissive than normally necessary to detect Wolbachia in other arthropods. Appropriate positive and negative controls were always included. To eliminate false positives, a positive response from at least one of the Wolbachia-specific primer sets was followed by a repeated PCR test as well as a reamplification using the product of the first PCR reaction as a template. When possible, amplification products were sequenced. Several control COI PCR products were sequenced to confirm the presence of T. tabaci DNA.
A subset of samples was retested using a nested PCR assay (Weeks et al. 2003). PCR was first performed using general eubacterial 16S primers 16S27 F (AGAGTTTGATCMTGGCTCAG) and 16S1513R (ACGGYTACCTTGTTACGACTT) (Weisburg et al. 1991). Total reaction volume was 25 μl, including 1 μl template DNA with 1.5 mM MgCl2. After 2 min of denaturation at 95°C, 38 cycles of amplification were performed within 1 min at 94°C, 1 min at 52°C, and 1 min at 72°C, followed by a single 5-min extension step at 72°C. The product of this first reaction was diluted (1 μl in 250-μl sterile water) and used as the template for a second PCR reaction using the Wolbachia protocol outlined above. Nested PCR was performed on a total of 24 thelytokous, 7 arrhenotokous, and 7 deuterotokous individuals.
Cytological analysis was performed on two thelytokous lines of T. tabaci collected from New York onion fields in 2005 to confirm the PCR results. Ovaries from 10 females for each line were removed and stained for both DNA and Wolbachia. Thrips were dissected in TBST (50 mM Tris-HCl, 150 mM NaCl, 0.1% Tween, 0.05% NaN3; pH 7.5), and ovaries and other somatic tissue were removed and placed on a lysine-coated glass slide and covered with a drop of TBST with 3.7% formaldehyde. After 30 min, most of the buffer was carefully removed, assuring adhesion of tissue to the slide. Slides were washed in TBST. The ovaries were blocked in TBST with 1% bovine serum albumin (BSA) for 1 h and incubated with 1:500 dilution of mouse anti-HSP60 antibody (Sigma-Aldrich, St. Louis, MO) in TBST-BSA and 1:500 dilution of RNase A (2 mg/ml) for 3 h at room temperature. This antibody specifically recognizes Wolbachia in nematodes and a wide range of arthropod hosts (Taylor and Hoerauf 1999, McGraw et al. 2002, M.E.C., unpublished data). After washing in TBST, ovaries were incubated in a 1:500 dilution of Alexa Fluor 488 labeled goat anti-rabbit antibody (Invitrogen-Molecular Probes, Eugene, OR) in TBST. Subsequently, ovaries were stained with 1 μg/ml DAPI (4′-6-diamidino-2-phenylindole; Invitrogen-Molecular Probes, Eugene, OR) for 5 min, washed, and stained with 5 μg/ml propidium iodide (Invitrogen-Molecular Probes) for 20 min. The propidium iodide solution was removed, and the tissues were covered with mounting media and coverslide. The presence of bacteria was examined using the DAPI, propidium iodide as well as fluorescent-labeled antibodies using a ×100 objective. As a positive control, Wolbachia-infected Drosophila ovaries were dissected, fixed, stained, and visualized at the same time.
Effect of Temperature on Reproductive Mode.
Two additional populations of T. tabaci collected from nonrotated onion fields were used in this experiment. One was male producing and originated near Potter, NY, in 2002, whereas the other was thelytokous and collected near Elba, NY, in 2004. Each population was separated into three subpopulations and maintained on thrips-free cabbage leaf disks at 18, 24, and 29°C and a photoperiod of 16:8 (L:D) and RH 55%. The male-producing and thelytokous populations were reared at these temperatures for one and three generations, respectively. The experimental unit was single virgin. For the thelytokous population, a random subsample of 50 larvae was taken from the first generation to establish a colony for examining the second generation. This procedure was repeated to create a colony for the third generation.
Relationship Between Reproductive Mode and Insecticide Susceptibility.
Thrips tabaci larvae collected from the onion fields in 2002 and 2003 described above were used in this experiment. An equivalent number of thelytokous and male-producing populations were examined. The thrips insecticide bioassay system (TIBS) developed by Rueda and Shelton (2003) and modified by Shelton et al. (2003) was used to determine the susceptibility of larvae to lambda-cyhalothrin (Warrior; Syngenta Crop Protection, Greensboro, NC) and methomyl (Lannate LV; DuPont, Wilmington, DE). TIBS estimates the susceptibility of thrips larvae to an insecticide by determining the percent that survive confinement within an insecticide-treated 0.5-ml microcentrifuge tube after 24 h. Based on earlier work with T. tabaci (Shelton et al. 2003), lambda-cyhalothrin at a concentration of 100 ppm was considered a diagnostic concentration, whereas methomyl at the same concentration was considered the most appropriate concentration to detect the potential for resistance development (A.M.S., unpublished data). For each population and insecticide combination, a total of 100 thrips larvae were subjected to TIBS. However, the experimental unit was 20 larvae, so n = 5, and percent mortality was determined after 24 h.
We predicted that male-producing populations would be resistant to lambda-cyhalothrin and more tolerant to methomyl compared with thelytokous populations. Therefore, we expected mean percent mortality from the diagnostic concentration for male-producing populations to be significantly lower than mortality for thelytokous ones when exposed to these insecticides. This potential association was examined by comparing mean percent mortality for male-producing populations with mortality for thelytokous populations using a one-way analysis of variance (ANOVA) and the procedure PROC GLM at P < 0.05 (SAS Institute 2003).
Results and Discussion
Three reproductive modes, thelytoky, arrhenotoky, and deuterotoky, were documented in T. tabaci populations collected from onion fields in New York. This is the first report of deuterotokous reproducing populations of T. tabaci. One half of the T. tabaci populations were thelytokous, whereas the other half included a mix of thelytokous, arrhenotokous, and deuterotokous individuals (Table 1). This also is the first report documenting multiple modes of parthenogenesis occurring in the same population. Thelytokous and male-producing populations were encountered annually and co-existed in eastern New York (i.e., Orange County) and central and western New York (i.e., Orleans, Oswego, Wayne, and Yates Counties). Previously in Yates County, NY, male onion thrips were captured on yellow sticky cards in 10 commercial onion fields over 3 consecutive yr (Gangloff 1999). However, during this same period and in the same area, no male T. tabaci were captured from other hosts such as wheat (three fields sampled), alfalfa (three fields sampled), or red clover (one field). This observation suggests that a factor or factors elicit the production of males in onion fields and not in other crops. Gangloff (1999) did not study the possibility that male T. tabaci occurred in other onion-producing regions in New York.
Sex allocation ratios of progeny produced by virgin T. tabaci from commercial onion fields in New York during 2002-2004
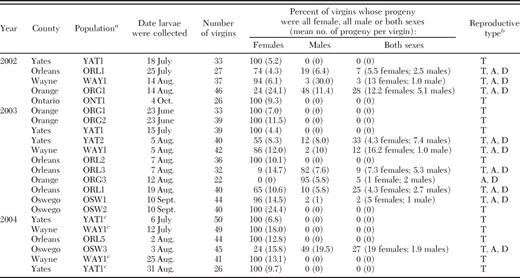
Based on these sex ratios, the mode or modes of parthenogenesis for each population was identified.
Onion field in which larvae (future virgin females) were collected.
Thelytoky (T), arrhenotoky (A), and deuterotoky (D). c These populations were sampled twice, once in July and later in Aug.
Sex allocation ratios of progeny produced by virgin T. tabaci from commercial onion fields in New York during 2002-2004
Based on these sex ratios, the mode or modes of parthenogenesis for each population was identified.
Onion field in which larvae (future virgin females) were collected.
Thelytoky (T), arrhenotoky (A), and deuterotoky (D). c These populations were sampled twice, once in July and later in Aug.
The average number of offspring produced per T. tabaci virgin was nearly identical for each reproductive mode (Table 1). Arrhenotokous virgins averaged 10.6 offspring (range: 1-30), whereas deuterotokous virgins produced an average of 8.8 female (range: 1-19) and 3.0 male offspring (range: 1-7.4). Each thelytokous virgin produced an average of 11.0 offspring (range: 4.4-24.4; Table 1). In male-producing populations, arrhenotokous and deuterotokous individuals averaged 32.2% (range: 2-95%) and 15.1% (range: 3-33%) of the total, respectively (Table 1).
Reproductive modes in T. tabaci populations varied temporally in several respects. For instance, the frequency of thelytokous and male-producing populations encountered during certain months significantly differed (χ2 = 9.94; df = 4,16; P = 0.04; Fig. 1). Ninety percent of the male-producing populations were detected from late July through August, and none were detected in June or October. In contrast, thelytokous populations were encountered from June through October and were prevalent in June and during the first half of July (Fig. 1). In two of four T. tabaci populations, reproductive modes varied across years (Table 2). Although sample size was small, these results suggest that populations either mix in some fields or an external factor changes the reproductive mode (e.g., a thelytokous population becomes a male-producing one or vice versa). The reproductive mode in two of two T. tabaci populations did not change within a season (Table 2).
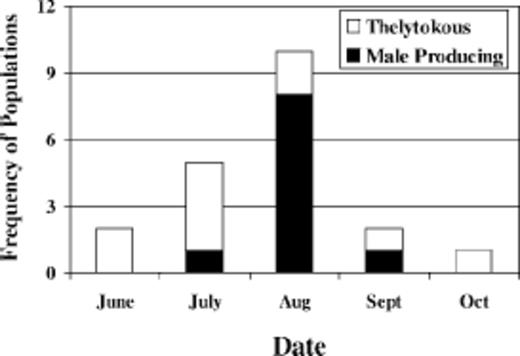
Number of thelytokous and male-producing populations of T. tabaci encountered during the season in New York onion fields from 2002 to 2004 (N = 20). Male-produc-ing populations included arrhenotokous and deuterotokous individuals.
Reproductive modes of T. tabaci populations from conunercial onion fields across seasons and within a season in New York during 2002-2004

Onion field in which larvae (future virgin females) were collected.
Male producing refers to a population that includes arrhenotokous and deuterotokous individuals.
Reproductive modes of T. tabaci populations from conunercial onion fields across seasons and within a season in New York during 2002-2004
Onion field in which larvae (future virgin females) were collected.
Male producing refers to a population that includes arrhenotokous and deuterotokous individuals.
Wolbachia were not detected in T. tabaci. A total of 70 thelytokous, 15 arrhenotokous, and 23 deuterotokous T. tabaci virgins or pooled progeny tested negative for Wolbachia, indicating that it is not required for altering parthenogenesis. This is based on two lines of evidence, PCR amplification, and cytological examination of ovaries. In no cases did both Wolbachia-specific primer sets indicate the presence of Wolbachia. In most cases where one or the other primer sets resulted in amplicons, Wolbachia was excluded as a template source because of either PCR product size, or the resulting sequence, or both. With one exception, the result of DNA sequencing of PCR products was either sequences with no known homology to Wolbachia or a chromatogram with multiple peaks—both consistent with the amplification of thrips DNA and not the Wolbachia genes for which the primers were designed. In one exception, sequenced PCR products were identified as Wolbachia. Repeated PCR using the same template DNA as well as others collected at the same site failed to repeat these results. Therefore, the source of this Wolbachia was likely a contamination. Sequencing and a subsequent BLAST search of several different COI PCR products confirmed that the DNA was intact and from T. tabaci. Although at least one additional bacterium has been shown to induce parthenogenesis in arthropods (Zchori-Fein et al. 2001), attempts to identify such organisms with our specimens have failed (J.L.W., unpublished data).
Cytological examination of the two thelytokous lines of T. tabaci using the DNA stains and the Wolbachia-binding fluorescent-labeled antibody revealed no Wolbachia in thrips ovaries. In the one case where bacteria were clearly evident, it was apparently spilling out of a portion of the digestive tract. Unlike Wolbachia, these bacteria were distinctly rod shapped and not recognized by the anti-Hsp60 (Wolbachia binding) antibody. The DNA staining method also has the potential to reveal other endosymbiotic bacteria, but none were observed within ovaries. Taken together, these results indicate that Wolbachia (or other endosymbiotic bacteria) are unlikely to be required for parthenogenesis induction in T. tabaci.
Temperature had no effect on parthenogenesis, at least over the range of temperatures examined (18-29°C; Tables 3 and 4). Sex allocation ratios in the male-producing and thelytokous populations were similar at each temperature. For the male-producing population, the percentage of males produced at each temperature was similar (Table 3). For the thelytokous population, no males were produced at any temperature over three consecutive generations (Table 4). Because only two populations were evaluated in this study, additional populations should be examined before temperature is dismissed as a potential parthenogenesis-altering factor. Alternatively, an external factor may not elicit male-producing forms of parthenogenesis in populations of T. tabaci. Rather, such populations may be descendents from the ancestral form of this species.
Sex of progeny produced by virgin T. tabaci from a male-producing population that originated from a commercial onion field near Potter in Yates County, New York in 2002 and maintained at each of three temperatures

Sex of progeny produced by virgin T. tabaci from a male-producing population that originated from a commercial onion field near Potter in Yates County, New York in 2002 and maintained at each of three temperatures
Sex of progeny produced by virgin T. tabaci from a thelytokous population that originated in a commercial onion field near Elba in Orleans County, New York in 2004 and maintained at each of three temperatures for three consecutive generations

Sex of progeny produced by virgin T. tabaci from a thelytokous population that originated in a commercial onion field near Elba in Orleans County, New York in 2004 and maintained at each of three temperatures for three consecutive generations
Male-producing populations of T. tabaci were not more resistant to lambda-cyhalothrin or more tolerant to methomyl compared with thelytokous populations. Mean percent mortality for male-producing and thelytokous populations exposed to lambda-cyhalothrin did not differ (P > 0.7; Fig. 2A). Similarly, mean percent mortality for male-producing and thelytokous populations exposed to methomyl did not differ (P > 0.9; Fig. 2B). Perhaps even more important than the absence of differences between means is that some populations of each reproductive mode were either clearly resistant or clearly susceptible to the insecticide, thereby disproving the hypothesis that reproductive mode can be used to predict resistance status.
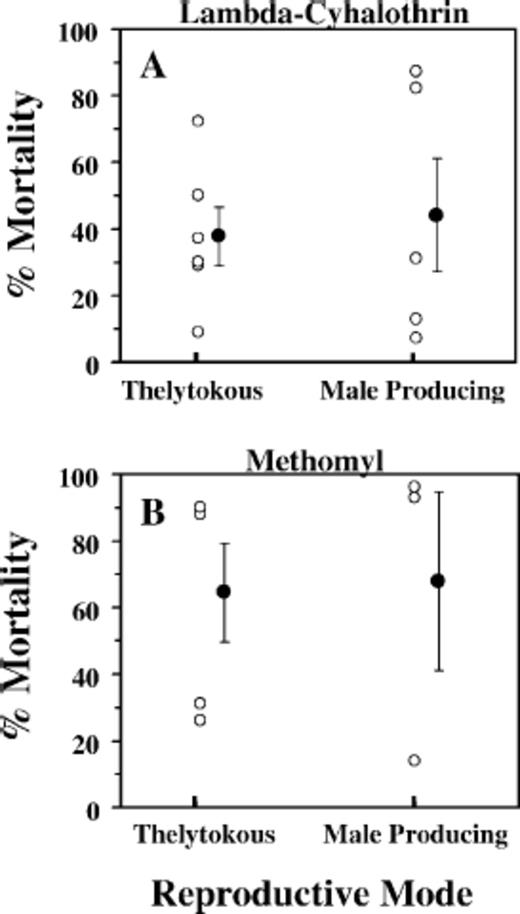
Percent mortality of T. tabaci larvae after a 24-h exposure to a diagnostic concentration of lambda-cyhalo-thrin at 100 ppm (A) or a 100 ppm concentration of metho-myl (B). Larvae originated from thelytokous and male-producing populations in New York onion fields during 2003 and 2004. Mean ± SEM) percent mortality is represented by the off set solid circles. Each open circle represents one population in which n = 300-500 larvae in the insecticide bioassay.
Male-producing populations used in the TIBS assays included thelytokous individuals comprising between 0 and 96% of the total individuals in the population. Therefore, the possibility existed that populations containing a high proportion of thelytokous individuals could be more susceptible than those containing a high proportion of arrhenotokous and deuterotokous individuals. Such a scenario would confound the ability to examine the potential association between male-producing populations and resistance to or more tolerance to insecticides. However, this scenario did not occur in our study. For both insecticides, mortality levels in male-producing populations were not correlated with the percentages of thelytokous thrips in these populations (lambda-cyhalothrin: F = 0.25; df = 1,3; P = 0.65; methomyl: F = 0.42; df = 1,1; P = 0.63).
The absence of a positive correlation between male-producing T. tabaci populations and susceptibility to insecticides indicates that scouting onion fields for male T. tabaci will not be helpful for detecting or predicting resistance. Thus, New York onion growers will need to continue relying on TIBS results for estimating T. tabaci's susceptibility to insecticides. Results from TIBS have indicated resistance occurs at the field rather than regional level and may change within the season. Although TIBS has been useful, it is currently used on only a small percentage of the onion acreage in New York. The reason for this is that TIBS is a new technique primarily developed for research purposes. Commercial use of this would require laboratory facilities and appropriate training. Perhaps in the future, crop consultants who scout large acreages of onions will invest in using TIBS.
The role of the different reproductive modes in T. tabaci populations in New York onion fields remains unclear. Understanding these reproductive modes may provide insight into certain biological attributes of a population such as the rapidity in which a population could develop insecticide resistance. Also, it would be interesting to know if these various reproductive modes occur in T. tabaci populations from other onion-growing regions and vegetable cropping systems.
We thank J. Plate for conducting the thrips insecticide bioassay system, K. Straight for maintaining thrips colonies, and G. English-Loeb for assisting with some of the statistics. The New York State Agricultural Experiment Station and USDA/CSREES-Hatch supported the research in this project.
References

{kind=link}
{kind=link}