





Abstract
Mississippi Kites (Ictinia mississippiensis) nesting in the Mississippi Alluvial Valley, USA, have consistently exhibited poor reproductive success, reduced average clutch sizes, and evidence of food stress during brood-rearing, raising concerns about population viability. Unlike populations elsewhere, kites nesting in the bottomland forests of this region face dynamic, anthropogenically altered hydrologic conditions that may be affecting the availability of important prey. Therefore, we quantified nestling diets and examined factors thought to be directly influencing the types and proportions of prey delivered to kite nestlings. Specifically, we sought to identify variables affecting the delivery of annual cicadas, the dominant prey item fed to kite chicks in numerous systems, as cicada emergence from subterranean burrows is known to be delayed by flooding. Using time-lapse video, we documented nestling diets and evaluated predictors of diet variability in east-central Arkansas, USA. We found that the delivery of cicadas increased with day of year, and was greatest during the driest of 4 study years. In contrast, the delivery of dragonflies, the numerically dominant prey item, declined with day of year, but increased with water level, and was lowest during the driest year. Although water level was not a strong predictor of the delivery of cicadas, interannual variation in the pattern of cicada deliveries suggests that flooding reduced the availability of this prey item to kites. Also, despite diverse nestling diets, the provisioning of dragonflies and a variety of other arthropods suggests that kites responded functionally to an absence of cicadas. The temporal patterns in prey deliveries that we detected imply that kite nestling diets in bottomland forests of the Mississippi Alluvial Valley may be influenced by water-level impacts on arthropod phenology and abundance.
Resumen
Los individuos de la especie Ictinia mississippiensis que anidan en el valle aluvial del río Mississippi en los Estados Unidos han mostrado constantemente bajas tasas de éxito reproductivo, tamaños de nidada reducidos y evidencia de estrés alimenticio durante la cría de los pichones, lo que ha despertado preocupación sobre la viabilidad de la población. A diferencia de poblaciones de otros lugares, los individuos que anidan en los bosques inundables de esta región enfrentan condiciones hidrológicas dinámicas y alteradas por el hombre, lo que podría afectar la disponibilidad de sus presas. Cuantificamos la dieta de los pichones de I. mississippiensis y examinamos los factores que podrían influir directamente en el tipo y proporción de presas que les son ofrecidas. Específicamente, quisimos identificar variables que influyen directamente en la disponibilidad de cigarras anuales, la presa más importante para los pichones en varias poblaciones, ya que se sabe que la salida de las cigarras de sus madrigueras subterráneas se retarda por las inundaciones. Usando vídeo por lapsos de tiempo documentamos la dieta y evaluamos los predictores de la variabilidad de la dieta en el centro-oriente de Arkansas, EEUU. Encontramos que la entrega de cigarras se incrementó con el día del año y fue mayor durante el año más seco de quatro años de estudio. En contraste, la entrega de libélulas (la presa más abundante) disminuyó con el día del año y se incrementó con el nivel del agua, y fue menor en el año más seco. Aunque el nivel del agua no fue un predictor importante de la entrega de cigarras, la variación entre años en el patrón de entrega de cigarras sugiere que las inundaciones reducen la disponibilidad de esta presa. Igualmente, a pesar de la dieta diversa de los pichones, la entrega de libélulas y otros artrópodos sugiere que I. mississippiensis responde funcionalmente a la ausencia de cigarras. Los patrones temporales en la entrega de presas que detectamos implican que la dieta de los pichones en los bosques inundables del valle aluvial del río Mississippi podrían verse influenciados por el impacto del nivel del agua en la fenología y abundancia de artrópodos.
Palabras clave: bosques inundables de tierras bajas, cigarra, dieta, entrega de presas, inundación, Ictinia mississippiensis, vídeo por lapsos de tiempo
Introduction
The Mississippi Kite (Ictinia mississippiensis, hereafter “kite”) is a medium-sized raptor and Neotropical migrant that breeds in portions of the southern United States (Parker 1999). Populations of kites breeding in bottomland hardwood forests of the Mississippi Alluvial Valley (MAV) have consistently exhibited low reproductive success and appear unable to fledge more than one chick per nesting attempt (Evans 1981, Barber et al. 1998, St. Pierre 2006, Bader and Bednarz 2009, Chiavacci 2010). In contrast, kites nesting in the Southwest and Great Plains experience notably higher nest success rates and are capable of fledging 2 chicks per nesting attempt (Parker 1974, Glinski and Ohmart 1983, Shaw 1985, Gennaro 1986). Reasons for these disparities between populations remain unknown, but food availability has been suggested as a cause of the difference in the frequency of 2-egg clutches (Parker 1974). In particular, productivity within parts of the MAV seems to be food-limited (Evans 1981, Bader and Bednarz 2011), as the occurrence of siblicide and cannibalism in the rarely produced 2-chick broods in Arkansas (Bader and Bednarz 2009, Chiavacci 2010) is indicative of food stress (Newton 1979); such behaviors have not been witnessed in populations elsewhere. Indeed, kites nesting in east-central Arkansas likely represent a population sink (Bader and Bednarz 2009). Because the amounts and types of food delivered to nestlings strongly affect nestling quality, nestling survival, and, ultimately, the annual productivity of adults (e.g., Simons and Martin 1990, Rodenhouse and Holmes 1992, Naef-Daenzer and Keller 1999), identifying diet composition and factors influencing variability in nestling diets may illuminate reasons for the poor reproductive performance of kites in the MAV.
Although not an obligate insectivore (Parker 1988), the majority of the Mississippi Kite's diet consists of insects (Parker 1999), with annual (i.e. nonperiodical) cicadas representing the primary prey delivered to chicks (Glinski and Ohmart 1983, Shaw 1985, Botelho et al. 1993, Bader and Bednarz 2011). Indeed, the nest success of kites in Arizona was found to be related to the volume of cicada-preferred vegetation around nest sites, demonstrating this insect's potential effect on kite reproduction (Glinski and Ohmart 1983). Likewise, some songbirds rely so heavily on cicadas during the breeding season that they time their nesting to coincide with the availability of this prey (Rosenberg et al. 1982). If this is also the case for Mississippi Kites, then nestling quality and reproductive success may be strongly influenced by the temporal correspondence between the provisioning of nestlings and cicada availability. However, the life cycle of cicadas results in their seasonal appearance being driven by environmental conditions. Nymphs develop underground for several years (Karban 1986), and the timing of their emergence into aboveground habitats is dictated partly by soil temperature (Smith et al. 2006). Consequently, factors affecting soil temperature (e.g., moisture content) directly influence when cicadas emerge (e.g., Andersen 1994), and thus when they are available to nesting kites.
Bottomland hardwood forests and their hydrologic cycles in the United States have been altered through fragmentation, waterway channelization, and the construction of water control structures (e.g., Pashley and Barrow 1993, Sharp 2012). Such alterations can affect the severity and frequency of flooding events (Harris and Gosselink 1990, reviewed in King et al. 2009), possibly making conditions within bottomland forest habitats more dynamic than those experienced historically (Sharp 2012). In fact, several recent studies have highlighted the negative consequences that severe flooding can have on songbirds in the MAV (Anich and Reiley 2010, Benson and Bednarz 2010, Reiley et al. 2013). These altered flood regimes may also be directly affecting the availability of cicadas to breeding kites, as inundated soils delay cicada emergence by lowering soil temperatures (Andersen 1994). Current evidence suggests that cicadas are emerging later in the MAV than elsewhere: Whereas cicadas were the primary prey delivered during the first 4 weeks of nestling development in Arizona (Glinski and Ohmart 1983), they were not the dominant prey in Arkansas until chicks were about 3 weeks of age (Bader and Bednarz 2011). Because of the Mississippi Kite's reliance on cicadas during brood-rearing, and the potential effects that cicadas can have on reproductive success, factors impacting the availability of cicadas may be contributing to the poor reproductive performance of kites in the MAV. Therefore, our objectives were to: (1) identify variables useful for predicting the types and proportions of prey items delivered to nestlings; and (2) determine the potential influence of intra- and inter-annual variation in hydrologic conditions on the delivery of cicadas relative to other prey items.
Methods
Study Area
We studied kites at White River National Wildlife Refuge (34°23′N, 91°05′W) in east-central Arkansas, USA, near the White River's confluence with the Arkansas River Canal. The refuge covers approximately 64,700 ha and is one of the largest remaining contiguous bottomland hardwood forests in the MAV. It consists primarily of bottomland hardwood forest, with some upland hardwood forest, scattered fallow and agricultural fields, hundreds of natural and man-made lakes, and many bayous and sloughs. The landscape surrounding the refuge is dominated by agricultural crops, primarily rice and soybeans.
Nest Searching and Video Monitoring
We searched for nests from early April, shortly before kites began arriving at the refuge, through early July in 2008 and 2009. Although there were likely active nests with chicks present after early July that we failed to locate, we ceased nest-searching at that time because the initiation of other aspects of our study precluded us from continuing focused nest searches. We searched for adult kites from a motorboat or kayak. Upon locating adults, we observed them for behaviors indicative of nesting (e.g., copulation, carrying nesting material and/or prey) and attempted to follow them to their nests. Upon locating a nest, we determined whether it was active based on the presence of an adult, and established its status based on adult behavior. We subsequently checked nests with a spotting scope or binoculars every 3–4 days from a distance of 50–100 m.
To quantify the diet of nestling kites, we deployed time-lapse video recording systems at a subset of nests. We used 2 types of video recording systems. One system consisted of an infrared camera (PC177IR-1color, Supercircuits, Liberty Hill, Texas, USA) mounted on a limb approximately 50 cm above, and facing directly down into, the nest. The other was a custom-designed infrared video recording system (Fieldcam: Field Television System: LDTLV/Box/Versacam/IR60, Fuhrman Diversified, Seabrook, Texas, USA). The latter system incorporated an 18× optical zoom and 4× digital zoom, enabling us to place the camera up to 20 m from a nest. We placed cameras by climbing the nest tree or an adjacent tree (depending on the camera system) using a climbing harness, tree spikes, and lanyards. Videos were recorded on either Sony VHS time-lapse recorders (SVT-LC300, New York, New York, USA), Supercircuits VHS time-lapse recorders (NCL3300, Liberty Hill, Texas, USA), or miniature digital video recorders (DVRs; Cox et al. 2012). For VHS recorders we used an 8-hr taperecording at 8 frames per second, and for DVRs we employed an 8 gigabyte secure digital high capacity (SDHC) card recording at 6 frames per second. Systems were powered by 1–3 deep-cycle marine batteries. We replaced VHS tapes and SDHC cards every 3 days and batteries as needed. We used a portable viewing monitor during nest checks to determine the status of the nest and to ensure the camera was properly focused on the nest. We monitored nests with cameras until they fledged or failed; cameras were then moved to other active nests. We are certain that we did not monitor prey deliveries at multiple nests of the same pair in a year because Mississippi Kites raise only one brood a year, although they may renest following failure early in incubation (Evans 1981). In Arkansas, most kites lay eggs from mid-May through early June, fledge chicks in late July or early August, and begin migrating out of the area in late August. Such a timeline would all but preclude kites from completing another nesting attempt following failure during even the early nestling stage in initial nests.
Video Identification of Prey
Video data were reviewed on a time-lapse VCR (analog video) or computer (digital video) to quantify prey deliveries. When reviewing prey deliveries, we attempted to identify prey items to the lowest taxonomic level possible. In some cases, we were able to identify body parts of an item (e.g., legs, abdomen, wings), but could not classify it to order. We classified such items into 1 of 3 general categories: arthropod, bird, and other vertebrate. We were able to categorize some prey as vertebrate due to the relatively longer time needed for adults to feed or nestlings to consume such items (particularly larger vertebrates), in addition to the typical behavior with which kites consumed vertebrates (i.e. forcefully pulling at tissue with their bills) relative to invertebrates, which were easily pulled apart. However, if the time taken to consume the item was relatively long, but the feeding behavior did not suggest that a vertebrate item was being consumed, we labeled the item as unidentified. Observations of feathers enabled us to identify items as birds, but we found it more difficult to identify other vertebrate prey to class level. In addition to identifying the prey item, we recorded year, day of year, and nestling age for each delivery. To minimize reviewer bias, all video data were reviewed by one observer (S. J. Chiavacci).
Statistical Analyses
To identify factors useful for predicting the proportions of prey items delivered to nests, we separated prey items into 5 categories: cicadas (Cicadidae), dragonflies (Odonata), other arthropods, vertebrates, and unknown. Because of the numerical dominance of cicadas and dragonflies in the diet relative to all other types of arthropods, we analyzed these 3 groups separately. To examine variation in prey proportions on a broader temporal scale, increase sample size, and improve the predictive strength of our models, we supplemented our 2008 and 2009 data with prey delivery data from 2 nests monitored in our study site in 2004 and 5 nests monitored in 2005 (Bader and Bednarz 2011). Bader and Bednarz (2011) employed the same field methods for locating, monitoring, and selecting nests at which to deploy cameras, and used the same analog video systems and video review methods as our study. One person (T. Bader) reviewed the 2004 and 2005 video data. For each nest used to examine predictors of prey items delivered, we tallied prey deliveries for each day that the nest was active and then calculated the proportions of cicadas, dragonflies, other arthropods, vertebrates, and unknown items delivered on each of those days. We then examined the proportions of these prey items delivered to nestlings using generalized linear mixed models with binomial distributions and logit link functions (PROC GLIMMIX; SAS Institute 2008, Littell et al. 2006), with the predictor variables of nestling age, year, day of year, and water level. We examined the effect of nestling age because Bader and Bednarz (2011) proposed that adults may feed nestlings certain prey items based on their ability to consume such items without adult assistance (i.e. tearing food apart), which is likely to be more pronounced when chicks are younger and smaller. For nests at which we did not record hatching on video, we assigned ages to chicks by (1) estimating the time elapsed since the start of incubation (if known), and (2) comparing feather growth patterns between chicks of known and unknown ages. We thought that day of year might explain variability in prey deliveries because of this kite's reliance on ephemeral insect populations, which likely vary in abundance and thus availability at different times of the breeding season. Also, because not all nestlings hatch at the same time, variation in the day of year when kites are feeding chicks may help to explain some diet variations. The White River flooded large portions of the refuge in 2008, 2009, and, to a lesser extent, 2004. We therefore examined daily White River water levels obtained via the U.S. Army Corps of Engineers' river gauge located in St. Charles, Arkansas, near the middle of the river's route through the refuge (U.S. Army Corps of Engineers 2008). To address nonindependence among samples (i.e. deliveries within nests), we treated nest as a random effect. We examined support for a priori candidate models by calculating Akaike's Information Criterion adjusted for small sample sizes (AICc), ranking each model using differences in AICc values (ΔAICc), and generating model weights (wi; Burnham and Anderson 2002). For the analysis of each prey category, we evaluated the global model for overdispersion using the ratio of the goodness-of-fit chi-square statistic to its degrees of freedom.
In all cases we considered models within 4 ΔAICc units of the top model to be competitive, and chose to examine the predictive strength of variables contained within this competitive model set. We evaluated all models for the existence of uninformative parameters (i.e. “pretending variables”; Anderson 2008) and did not use these models in model-averaging or when drawing inference from models (Arnold 2010). In cases of model selection uncertainty, we calculated model-averaged parameter estimates for the variables contained within the set of models deemed competitive. Prior to fitting models, we examined correlations between all covariates and excluded from the same model those that were highly correlated (|r| > 0.7). Means are reported ± SE. All analyses were performed in SAS, version 9.2 (SAS Institute 2008).
Results
We located 41 kite nests during 2008 and 2009, 32 of which eventually contained incubating adults. Seventeen nests hatched 1 nestling, and 2 hatched 2 nestlings. We recorded video at 17 (2008 = 6, 2009 = 11) of 27 nests that survived 7 days beyond the start of incubation; we did not set up cameras earlier than 7 days after the start of incubation to avoid nest abandonment. We excluded 3 of 27 nests from those eligible to receive cameras because of tree features that made climbing unsafe. Of the 17 nests at which we recorded data, we excluded 7 from dietary analyses due to poor video quality (n = 3) or nest failure before or during the first 3 days of the nestling stage (n = 4, including one 2-chick brood), thus providing little or no dietary data. We quantified prey deliveries at the 10 nests for which we had good quality video (4 in 2008 and 6 in 2009) and at which we began recording video when nestlings were ≤1 day of age. We recorded 363 nest days of video data, comprising 5,275 hr of video during which prey deliveries could occur (i.e. daylight hours). We failed to record video during 533 hr (9%) of the nestling period, primarily due to equipment failure, but video loss was evenly distributed among nests and the nestling period. Of the 10 nests used for dietary analyses, 8 fledged 1 chick each and 2 were depredated when nestlings were 14 and 39 days of age. One nest contained 2 chicks until the younger, which was 16 days of age, was killed and consumed by its older sibling. We did not treat this nest differently than single-chick nests.
We observed 9,017 prey deliveries at these 10 nests, 6,617 (73%) of which we were able to identify as at least arthropod or vertebrate. Of the prey items identified to 1 of our 3 general categories, 5,987 were classified to order and 1,641 were further classified to family (Table 1). We documented 8 orders and 23 families of arthropod, and 5 orders and 7 families of vertebrate (Table 1). In addition, we were able to classify 591 (7%), 31 (<1%), and 8 (<1%) prey items that we could not identify to order into the more general categories of arthropod, vertebrate, and unidentified bird species (likely passerines), respectively. We failed to identify 2,400 (27%) prey items. Failure to identify items was due to various reasons, including items being too small to identify to at least class (46%), our view being blocked by the body of an adult or nestling (40%), poor quality video (13%), and items being fed to nestlings out of the view of the camera (1%). Of the nests that fledged chicks, mean age at fledging was 39.6 ± 1.2 days. Mean total items delivered day−1 peaked when nestlings were 21.4 ± 2.2 days of age. The mean sum of prey deliveries to nests that fledged chicks (n = 8) or survived to the mean fledging age of 39 days (n = 1) was 972 ± 59. The frequency of prey items in the diet varied considerably among nests for all 5 prey groups (Figure 1). Nestling diet data recorded in 2004 and 2005 included 7 nests and 2,849 prey deliveries, the majority of which were cicadas (52.1%) and dragonflies (26.1%). Nest monitoring during 2004 and 2005 began at various nestling ages (x̄ = 11 ± 3 days of age; range = 1–24).
Prey items delivered to Mississippi Kite nestlings in White River National Wildlife Refuge, Arkansas, USA, 2008–2009. Prey were separated into broad categories (arthropod, vertebrate, unknown) and, when possible, identified further into order and family.
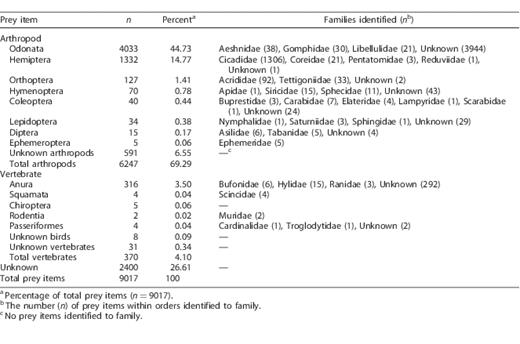
Prey items delivered to Mississippi Kite nestlings in White River National Wildlife Refuge, Arkansas, USA, 2008–2009. Prey were separated into broad categories (arthropod, vertebrate, unknown) and, when possible, identified further into order and family.
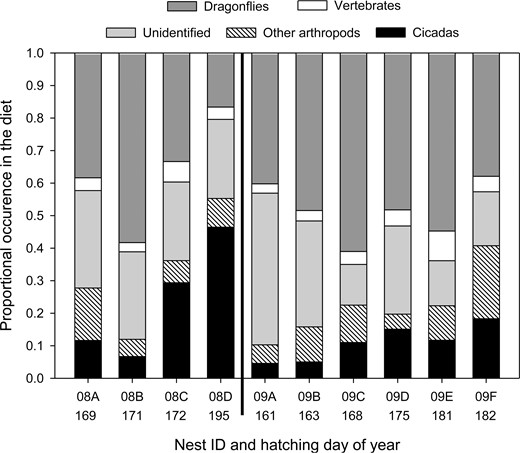
Mississippi Kite nestling diet at 10 nests in White River National Wildlife Refuge, Arkansas, USA, in 2008 (nests to left of solid black line) and 2009 (nests to right of solid black line). In 2008, hatching day 169 = June 17 and hatching day 195 = July 13. In 2009, hatching day 161 = June 10 and hatching day 182 = July 1.
White River water levels were above the long-term average during 2004, 2008, and 2009, and lower than the average in 2005 (Figure 2). The variables nestling age and day of year (|r| = 0.71) and water level and year (|r| = 0.79) were highly correlated and so were excluded from occurring together in the same model. We did not detect evidence of overdispersion in our global models (ĉ ≤ 1) and proceeded to examine the relative support of candidate models.
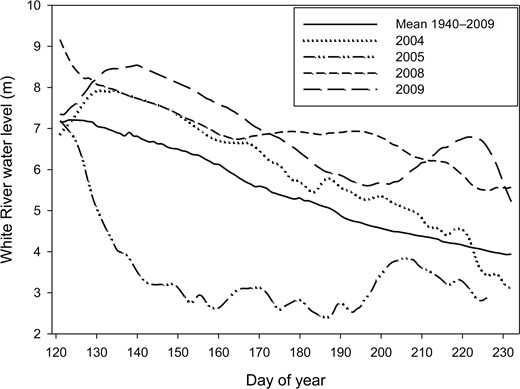
White River water level from May 1 (day 120) to August 20 (day 232) in 2004, 2005, 2008, and 2009, and mean river water level from 1940 to 2009. Water levels were obtained from the Army Corps of Engineers' river gauge located in St. Charles, Arkansas, which is situated near the middle of White River National Wildlife Refuge, Arkansas, USA.
The model that best predicted the proportion of cicadas delivered to nestlings included the effects of day of year and year, and had 4.8 times more support than the second-best model based on an evidence ratio of Akaike weights (Table 2). After model-averaging, we found that the proportion of cicadas delivered increased consistently with the day of year during all years, but the proportions of cicadas delivered to nestlings were much greater in 2005, the year with below-average water levels, than in years with above-average water levels (Figure 3); cameras were deployed later in 2004, accounting for the lack of cicada data before day 202.
Results of generalized linear mixed models predicting the proportion of cicadas, dragonflies, other arthropods, vertebrates, and unknown prey items delivered to Mississippi Kite nestlings in White River National Wildlife Refuge, Arkansas, USA, in 2004, 2005, 2008, and 2009. Models were ranked using differences in Akaike's Information Criterion adjusted for small sample size (ΔAICc). Bold ΔAICc values and associated model weights (wi) denote competitive models (i.e. ΔAICc < 4) within prey item model sets. Prey delivery data from 2004 and 2005 are from Bader and Bednarz (2011).
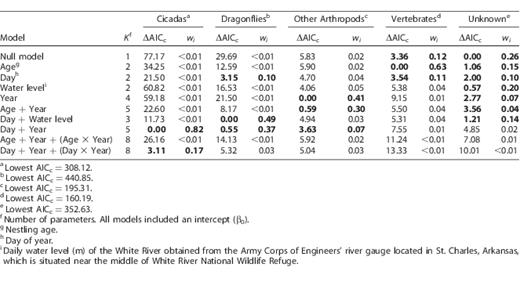
Results of generalized linear mixed models predicting the proportion of cicadas, dragonflies, other arthropods, vertebrates, and unknown prey items delivered to Mississippi Kite nestlings in White River National Wildlife Refuge, Arkansas, USA, in 2004, 2005, 2008, and 2009. Models were ranked using differences in Akaike's Information Criterion adjusted for small sample size (ΔAICc). Bold ΔAICc values and associated model weights (wi) denote competitive models (i.e. ΔAICc < 4) within prey item model sets. Prey delivery data from 2004 and 2005 are from Bader and Bednarz (2011).
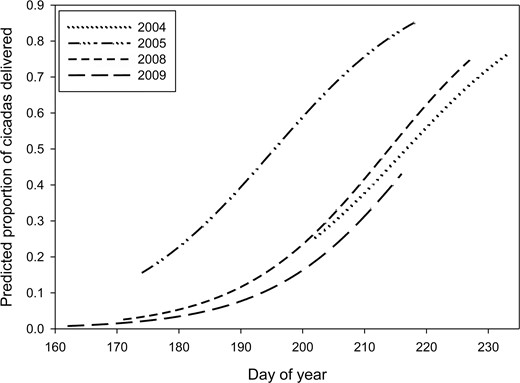
Model-averaged predicted proportions of cicadas delivered to Mississippi Kite nestlings in relation to day of year in White River National Wildlife Refuge, Arkansas, USA, in 2004, 2005, 2008, and 2009. Day 160 = June 9 and day 230 = August 18. Prey delivery data from 2004 and 2005 are from Bader and Bednarz (2011).
Dragonfly deliveries were best predicted by a model including the main effects of day of year and water level, which received slightly more support than the second-best model (day of year + year; Table 2). We examined the model-averaged predicted values of day of year, water level, and year separately, while holding other variables constant at their mean values. The proportion of dragonflies delivered decreased with day of year, but increased with water level (Figure 4). Additionally, 2005 (the driest year) had the lowest predicted proportion of dragonflies delivered (0.24 ± 0.05), with greater proportions delivered in years with above-average water levels (2004: 0.34 ± 0.14; 2008: 0.41 ± 0.05; 2009: 0.51 ± 0.04).
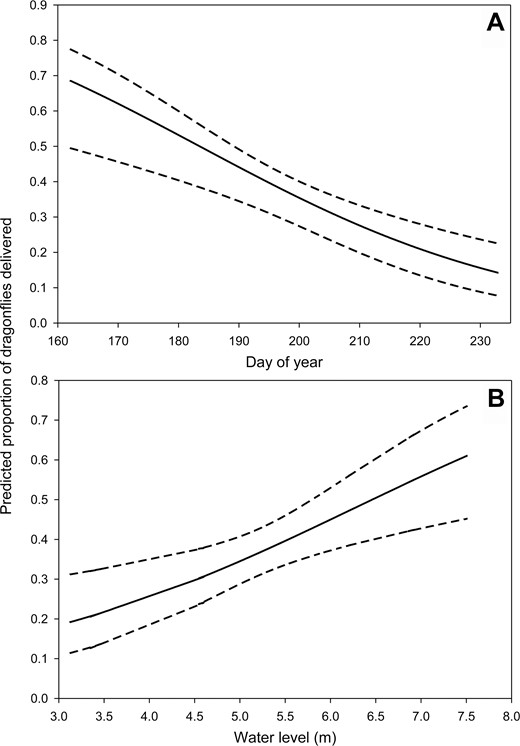
Model-averaged predicted proportion and 95% confidence limits of dragonflies delivered to Mississippi Kite nestlings in relation to (A) day of year (day 160 = June 9 and day 230 = August 18) and (B) water level in White River National Wildlife Refuge, Arkansas, USA, in 2004, 2005, 2008, and 2009. We held water level and year at their mean values when examining the influence of day of year. Likewise, year and day of year were held at their mean values when examining the effect of water level. Prey delivery data from 2004 and 2005 are from Bader and Bednarz (2011).
The delivery of other arthropods was best explained by a year effect. Following model-averaging, we found that the lowest proportion of other arthropods was delivered in 2004 (0.01 ± 0.01), and increased in 2005 (0.03 ± 0.02), 2008 (0.09 ± 0.02), and 2009 (0.10 ± 0.02). Although the presence of nestling age and day of year in competitive models explaining the delivery of other arthropods suggested that these variables were important, the parameter estimates for both nestling age (β̂ = −0.02; 95% CI: −0.04 to 0.01) and day of year (β̂ = −0.01; 95% CI: −0.03 to 0.01) overlapped zero, indicating that these factors added little explanatory power to the more simple model with only a year effect. The proportion of vertebrates delivered was best explained by nestling age; this model had 5.4 times more support than the second-best model containing only the intercept (null model; Table 2). Vertebrate deliveries decreased as nestlings aged (β̂ = −0.04; 95% CI: −0.08 to −0.01). No variables that we examined were good predictors of the delivery of unidentifiable prey items (Table 2).
Discussion
Mississippi Kites nesting in east-central Arkansas, USA, delivered a wide variety of prey items to their nestlings, although dragonflies and cicadas constituted nearly 60% of all prey deliveries. We found that the types and proportions of prey delivered to nestlings varied with nestling age, water level, and within and among years. These patterns illuminate potential reasons for differences in nestling diets among kite populations. Additionally, our findings suggest that environmental conditions within the Mississippi Alluvial Valley (MAV), specifically the severity and frequency of flooding events, may be influencing the availability of cicadas, which, in turn, could be contributing to the poor productivity of kites observed in the region (e.g., Bader and Bednarz 2009).
The proportion of cicadas in nestling diets varied within and among years. Differences among studies in the timing of cicada deliveries to Mississippi Kite nests indicate that temporal variability in when cicadas are available is most likely driving these patterns. In Arizona, USA, Apache cicadas (Diceroprocta apache) were the predominant prey during approximately the first 4 weeks of nestling development (day of year range during which deliveries were observed: 182–232; Glinski and Ohmart 1983). However, at about 5 weeks of age (ca. days 215–222) kites switched to delivering primarily beetles, likely because declines in cicada abundance drove kites to prey more heavily on beetles (Glinski and Ohmart 1983). In contrast, in Arkansas, USA, kites delivered mainly dragonflies during the first 2 weeks of nestling development, then switched to primarily cicadas when nestlings were about 3 weeks of age (ca. days 203–210 in 2004 and days 187–194 in 2005; day of year range during which deliveries were observed: 174–233), which Bader and Bednarz (2011) suggested may have been due to the physical inability of nestlings to consume cicadas without the aid of an adult. We did not find support for Bader and Bednarz's (2011) hypothesis, as our nestling age model received no support within our model set predicting cicada deliveries. Although we also observed a shift from dragonfly to cicada deliveries during 2008 and 2009, upon combining our data with those of Bader and Bednarz (2011), we found that cicada deliveries were best predicted by year and day of year. Thus, it appears that kites in Arkansas respond functionally to an absence of abundant cicadas early in the breeding season by preying more heavily on dragonflies and other arthropods in years of reduced cicada numbers. The same functional response appears to have occurred in Arizona, except in reverse order; cicadas made up the majority of the diet earlier in the nestling stage, but were replaced with beetles as cicada numbers declined (Glinski and Ohmart 1983). This is similar to behavior observed in the closely related Plumbeous Kite (Ictinia plumbea), for which the delivery of dragonflies and lizards to nestlings depended on the proportion of cicadas in the diet (Seavy et al. 1997). While we did not directly measure prey abundance, we propose that Mississippi Kites would have selected annual cicadas over other large arthropods (i.e. dragonflies) when given a choice, as annual cicadas collected in Arkansas (n = 40, x̄ = 0.657 ± 0.027 g dry mass, male and female cicadas combined) had >1.5 times the biomass of the largest dragonflies delivered to nests (i.e. aeshnids; n = 10, x̄ = 0.399 ± 0.038 g dry mass; S. J. Chiavacci personal observation).
Unlike cicada deliveries, dragonfly deliveries were positively related to water level and negatively related to day of year. Water level may have influenced the frequency of dragonflies in the diet through increased vulnerability to kite predation. Flooding likely created an abundance of suitable breeding habitat for dragonflies during flood years (i.e. sites for ovipositing eggs; Corbet 1999) possibly leading to increased dispersal of dragonflies (Batzer and Wissinger 1996) typically concentrated around localized water bodies. As kites may obtain over 40% of their prey within 150 m of their nests (Glinski and Ohmart 1983), more water around nest sites may have increased the number of dragonflies readily available to kites. However, day of year showed a stronger relationship with the proportion of dragonflies in kites' diets. This may illustrate a declining reliance on and predation of dragonflies, due to a preference for, and increasing availability of, cicadas. Indeed, dragonfly proportions were lowest, whereas cicada proportions were highest, in kites' diets in 2005 (the driest year in our analysis; Figure 2), lending support to our notion that cicadas emerged earlier in 2005, resulting in kites preying less on dragonflies. Alternatively, this pattern may imply that dragonflies declined in abundance as the season progressed or under dry conditions, and that their presence in the diet of nestlings was proportional to their availability rather than due to a reduced preference by kites.
Variability in the availability of cicadas may also explain interannual patterns in the delivery of other arthropods. For example, other arthropods were delivered in much higher proportions in 2008 and 2009 than in 2005. We suspect that, had 2004 data been collected earlier in the season, the proportion of these prey in the diet in that year may also have been greater than that in 2005. If true, this would suggest that kites broadened their diet in the absence of abundant cicadas (i.e. their preferred prey), a general dietary pattern found in other raptors (Steenhof and Kochert 1988). In contrast, the proportion of vertebrate deliveries varied little among years in our system, implying that predation of vertebrates was unaffected by the flooding that likely dictated predation of some invertebrate prey. Unlike deliveries of other prey, however, vertebrate deliveries were weakly related to nestling age, possibly reflecting the increased nutritional needs of kite nestlings early in their development (Robinson 1957). The greater delivery of vertebrates early in nestling development may enable adults to more efficiently meet the energetic demands of young nestlings, by delivering larger and likely more energy-rich prey during a period of relatively rapid growth (1–10 days of age; Robinson 1957).
Although the causal mechanisms that led to variation in cicada deliveries are unknown, we propose that flooding and its effects on soil temperature influence the within-season timing of cicada availability. In preparation for emergence, annual cicada nymphs burrow toward the soil surface and use soil temperature as a cue for when to move above ground (Smith et al. 2006). Factors that lower soil temperature, such as flooding, lead to delayed nymph emergence from areas that are inundated (Andersen 1994). We surmised that such a situation affected the proportions of cicadas in the diets of kite nestlings during years with extensive flooding (2004, 2008, and 2009), and used the level of the White River as an indirect measure of flooding extent. Our failure to find a link between river water level and cicada deliveries was likely due to the complexity of flooding patterns in White River National Wildlife Refuge, which are influenced by three major rivers, numerous water control structures, and the physical features of the refuge (e.g., floodplain width, microtopography; Benson et al. 2011). For example, whereas higher water levels upstream result in headwater flooding, rises in the Mississippi River to the south cause the White River to back up in the southern portion of the refuge, leading to nonuniform patterns in flooding extent (Sharp 2012). Flooding severity is exacerbated by a large levee bordering the southern half of the refuge to the east (Sharp 2012). Lastly, the temporal relationship between river water level and cicada emergence may not have corresponded because high water levels early in the year may have had lasting effects on emergence timing through differences in water retention and persistence in some areas. To have explored these possibilities would have required our collecting data on a much finer scale. However, the largest proportions of cicadas were delivered to nests in 2005, the year with the lowest White River water level (Figure 2), providing support for our assertion that flooding conditions affected cicada emergence timing (i.e. availability) and, thus, the proportions of cicadas fed to nestlings. Indeed, the river level in 2005 was considerably lower than the long-term average (Figure 2) and may have resulted in cicadas emerging earlier than usual, making 2005 a good year with respect to the temporal availability of cicadas to brood-rearing kites.
Our results illustrate the need for a better understanding of how variability in nestling diets may be contributing to depressed reproductive performance of kites in White River National Wildlife Refuge (Bader and Bednarz 2009) and elsewhere in the MAV (Evans 1981, Barber et al. 1998). Black Kites (Milvus nigrans), a wetland-dependent species, have been found to deal with unpredictable inundation levels and subsequent effects on prey via adjustments in clutch size, hatching success, and brood reduction (Sergio et al. 2011). Similar unpredictability in prey availability due to abiotic factors such as flooding may partly explain why Mississippi Kite nests in the MAV only rarely contain 2-egg clutches, why kites have not been observed to fledge more than one chick per nesting attempt (Evans 1981, Barber et al. 1998, St. Pierre 2006), and why chicks express facultative siblicide and cannibalism (Bader and Bednarz 2009, Chiavacci 2010). Also, if the frequent delivery of cicadas throughout the majority of the nestling period in Arizona (Glinski and Ohmart 1983) holds true for much of the Southwest and Great Plains, the presumably greater availability of cicadas during this period may explain why kites more frequently fledge 2 chicks in these areas (Parker 1974, Shaw 1985, Gennaro 1986). To our knowledge, very little is known about factors influencing the phenology of annual cicadas in the MAV, despite the fact that they represent an important food resource for many breeding birds (e.g., Rosenberg et al. 1982). As well as altering the timing of cicada emergence, flood duration could also be affecting cicada availability through reduced nymph survival. Although cicada nymphs seem capable of surviving short-term flooding while underground (Marlatt 1907, Andersen 1994), it remains unclear whether persistent, long-term inundation causes nymph mortality. Such information would be valuable for further assessing our hypothesis that anthropogenically altered flooding regimes are influencing the availability of cicadas to brood-rearing kites by affecting emergence timing and possibly nymph survival. Lastly, expanding the study of Mississippi Kite nestling diet and reproductive success to include the largely unstudied breeding areas east of the MAV, as well as recently colonized urban and suburban habitats, would provide useful comparative information about this species' diet and reproductive performance.
Acknowledgments
This project was funded by the U.S. Fish and Wildlife Service (USFWS) and the Arkansas Game and Fish Commission (AGFC) through a State Wildlife Grant, and by the Arkansas Audubon Society Trust through a Student Research Award to S. J. Chiavacci. We thank Karen Rowe (AGFC) for her support throughout this project, and Richard Hines (USFWS) and the staff of White River National Wildlife Refuge for their assistance. T. Bader graciously provided data for our analyses and assistance during the early stages of this project. Comments by two anonymous reviewers improved an earlier version of this manuscript. We thank J. Everitts for measuring cicada and dragonfly biomass. B. Bawden, N. Chiavacci, D. Noel, and B. Schneider assisted with fieldwork.
Literature Cited
Author notes
Current address: Illinois Natural History Survey, Prairie Research Institute, University of Illinois, Champaign, Illinois, USA
Current address: Department of Biological Sciences, University of North Texas, Denton, Texas, USA

{kind=link}
{kind=link}
{kind=link}
{kind=link}