





Abstract.
Large-scale disturbances, by affecting habitat suitability, can influence bird populations and communities. Although we know that disturbances can lead to population decreases and shifts in community structure, our understanding of finer-scaled mechanisms leading to these patterns is limited. We focused on why some territories remain occupied by a species following a large-scale disturbance, while others are abandoned. The Swainson's Warbler (Limnothlypis swainsonii) is especially susceptible to flooding because of its preference for nesting in thick understory vegetation and foraging in leaf litter. In 2008, a major flood inundated a study site where we have studied Swainson's Warblers since 2004. In 2008 and 2009, we resurveyed 42 Swainson's Warbler territories that were occupied between 2005 and 2007 and examined changes in habitat structure and composition and prey abundance influencing territory abandonment. Occupancy declined between the preflood and postflood surveys. Territory abandonment was associated with decreases in leaf litter cover, increases in cover of green vegetation, and decreases in density of total understory stems and giant cane (Arundinaria gigantea) between the preflood and flood periods. Despite the partial recovery of habitat structure and prey abundance after the flood, Swainson's Warbler occupancy did not recover in subsequent years. Multiple habitat variables, including leaflitter cover, density of woody stems, understory density, and prey abundance have been identified as important for Swainson's Warblers, and these results suggest that leaf litter may be especially important in influencing habitat-selection decisions following flood disturbance.
Resumen.
Los disturbios de gran escala, al afectar la aptitud del habitat, pueden influenciar a las poblaciones y comunidades de aves. Aunque sabemos que los disturbios pueden llevar a disminuciones poblacionales y cambios en la estructura comunitaria, nuestro entendimiento de los mecanismos de sintonáa fina que llevan a estos patrones es limitado. Nos enfocamos en entender por qué una especie continúa ocupando algunos territorios luego de un disturbio de gran escala, pero abandona otros. Limnothlypis swainsonii es una especie particularmente susceptible a las inundaciones debido a sus preferencias por anidar en la vegetación densa del sotobosque y por forrajear en la hojarasca. En 2008, una gran creciente inundó un sitio de estudio donde habíamos estudiado a L. swainsonii desde 2004. En 2008 y 2009, volvimos a muestrear los territorios de 42 individuos de L. swainsonii que estuvieron ocupados entre 2005 y 2007 y examinamos los cambios en la estructura y composición del hábitat y la influencia de la abundancia de presas en el abandono del territorio. La ocupación disminuyó entre los muestreo previos y posteriores a la inundación. El abandono del territorio estuvo asociado con la disminución de la cobertura de la hojarasca, el aumento de la cobertura de la vegetación verde y la disminución en la densidad total de tallos del sotobosque y de caña gigante (Arundinaria gigantea) entre los dos muestreos. A pesar de la recuperación parcial de la estructura del habitat y la abundancia de presas luego de la inundación, la ocupación de L. swainsonii no se recuperó en los años subsiguientes. Múltiples variables del hábitat, incluyendo la cobertura de la hojarasca, la densidad de tallos leñosos, la densidad del sotobosque y la abundancia de presas han sido identificadas como importantes para L. swainsonii, y estos resultados sugieren que la hojarasca puede ser particularmente importante en influenciar las decisiones de selección de habitat luego del disturbio casado por la inundación.
Introduction
Natural and anthropogenic disturbances play a fundamental role in maintaining the habitat and landscape heterogeneity many birds require (Brawn et al. 2001). Understanding the effects of these disturbances may improve our knowledge of birds' ecology and can have important conservation implications (Hunter et al. 1987). Studies of the effects of disturbance on birds have often focused on population- and community-level changes such as declines or increases in densities of particular species (Knopf and Sedgwick 1987, Knutson and Klaas 1997). Less emphasis has been placed on the actual mechanisms leading to these population- and community-level responses, namely, the responses of individual birds to habitat change and their decisions about whether to abandon or occupy a given area. Disturbances do not necessarily affect all previously occupied territories within a given area equally, and individual responses to relative changes in various habitat attributes may provide insight into which aspects of habitat are most important for a given species.
Flooding is a disturbance that can affect birds by destroying nests, changing foraging substrates and the availability of food resources, or altering habitat structure and composition (Hunter et al. 1987). In particular, flooding may kill understory vegetation (Christman 1984, Yin 1998, Benson and Bednarz 2010) and reduce the diversity and abundance of ground-dwelling arthropods (Uetz et al. 1979). Given this large influence on the lower strata of forests, understory-dependent and ground-foraging species are especially vulnerable to major floods (Knopf and Sedgwick 1987, Knutson and Klaas 1997, Anich and Reiley 2010, Benson and Bednarz 2010). However, when multiple habitat components change simultaneously with flooding, it is often difficult to pinpoint which specific habitat attributes birds assess when selecting habitat.
One understory-dependent species that is sensitive to flooding is the Swainson's Warbler (Limnothlypis swainsonii). This species breeds in the southeastern United States, primarily in bottomland hardwood forests, winters in the Caribbean basin, and inhabits areas with a dense understory (Anich et al. 2010b). Pairs are socially monogamous; males maintain fidelity to a breeding site and defend their territory against conspecific males (Anich et al. 2010b). The Swainson's Warbler is uncommon and local throughout its breeding range and is ranked among the highest-priority species for conservation (USFWS 2008). Swainson's Warblers forage for arthropods in leaf litter, typically by flipping dead leaves and poking their bills into litter on the ground (Meanley 1970, 1971, Barrow 1990, Graves 1998). Within bottomland hardwood forests, this species generally breeds in the areas least prone to floods and may be affected by extreme levels of flooding (Meanley 1966, Graves 2001, 2002, Benson et al. 2011). Furthermore, many of the areas of highest elevation within bottomland hardwood forests have been converted to agricultural fields (Twedt and Loesch 1999), and the depth and duration of flooding of the remaining bottomland forest may be increased as a result of existing flood-control levees (Megonigal et al. 1997, Graves 2001, Anich and Reiley 2010).
On the basis of previous work, we know that the overall density of the understory, abundance of woody stems in the understory (especially of giant cane, Arundinaria gigantea), litter cover and depth, arthropod abundance, and lack of green vegetation at ground level are associated with habitat selected by Swainson's Warblers (Benson et al. 2009, Brown et al. 2009, 2011, Anich et al. 2012). However, these studies were done during a relatively dry period where flooding was not an important factor. In selected study areas, we also have demonstrated that flooding affects Swainson's Warbler habitat and can reduce the population of the area flooded (Anich and Reiley 2010, Benson and Bednarz 2010). Some birds persist in flood-affected areas despite changes in habitat, although the specific factors associated with continued occupancy or abandonment of a territory are unknown.
To determine how flooding affects habitat suitability, and what specific habitat changes are associated with territory abandonment, we investigated the relationship between changes in habitat and prey abundance and the changes in territory occupancy following a prolonged flood. Specifically, we surveyed for Swainson's Warblers at sites that were occupied during a previous study of nest-site selection and breeding ecology (Benson et al. 2009), and we examined what habitat changes were most associated with territory abandonment. We expected territories with greater reductions in understory habitat, leaf litter, and prey abundance to be more likely to be abandoned.
Study Area
White River National Wildlife Refuge is one of the largest contiguous tracts of remaining bottomland hardwoods in Arkansas and the southeastern U.S. (Twedt and Loesch 1999). Our study site was at a relatively high elevation, encompassing approximately 31 km2 near Alligator Lake in the southern portion of the refuge. The typical habitat used by Swainson's Warblers was dominated by sugarberry (Celtis laevigata), sweetgum (Liquidambar styraciflua), box elder (Acer negundo), elm (Ulmus spp.), American sycamore (Platanus occidentalis), and hickory (Carya spp.). Common understory species included spicebush (Lindera benzoin), boxelder, giant cane, greenbriar (Smilax spp.), Virginia creeper (Parthenocissus quinquefolia), peppervine (Ampelopsis arborea), and grape (Vitis spp.). In the refuge, Swainson's Warblers generally occupy areas at relatively higher elevation and less prone to flooding (Brown et al. 2009, Benson et al. 2011).
Our study site was unaffected by flooding from 2005 to 2007, but during the early spring of 2008, significant amounts of rain in the upper White River basin led to above-average flows of the White River and extreme flooding of many low-elevation areas. The crest in 2008 was the fifth highest on record at 10.53 m, approaching the previous high crest of 11.13 m recorded in 1973 (U.S. Army Corps of Engineers 2008). This flood inundated most of the refuge during April and early May 2008, including our entire Swainson's Warbler study site. The flooding coincided with the species' arrival and territory settlement, but some birds persisted in the tree canopies until flooding began to subside in May (Anich and Reiley 2010).
Methods
From late April to early August 2005 through 2007, Benson et al. (2009) used passive and song-playback surveys to locate territorial Swainson's Warbler males at White River National Wildlife Refuge. After locating territorial males, researchers searched for nests in those territories, found 91 active nests, and recorded data on vegetation characteristics (Martin et al. 1997, Benson et al. 2009) and arthropod abundance at each location. In 2008 (the “flood year”) and 2009 (the “postflood year”), we surveyed for Swainson's Warblers at 42 territories occupied from 2005 to 2007 and recorded vegetation and arthropod data within these territories (Benson et al. 2009, Brown et al. 2011).
Avian Surveys and Vegetation Sampling
Between 15 April and 15 June 2008 and 2009, we resurveyed 42 previously occupied territories three and two times per season, respectively, using song playback (Bednarz et al. 2005, Brown et al. 2009). We defined a territory as occupied if a Swainson's Warbler was detected within 100 m of its original detection location. Before the flood, the mean size of a Swainson's Warbler at our study site was 9.38 ha (Anich et al. 2009), and territories were generally adjacent to at least one neighboring territory throughout our study site (Nick Anich, unpubl. data). Repeated surveys within a season allowed us to account for imperfect detection (MacKenzie et al. 2006). In July of each year, we sampled vegetation characteristics at randomly selected locations of nests that were active between 2005 and 2007 (Benson et al. 2009) and fell within a 200-m buffer centered around survey locations (2008–2009). We chose a 200-m buffer because it encompasses a Swainson's Warbler's average home range size of 9.38 ha in Arkansas (Anich et al. 2009, 2010a).
We recorded data on vegetation structure and cover by using modified BBIRD protocols (Benson et al. 2009). The habitat variables we analyzed were chosen on the basis of published literature (Meanley 1971, Graves 2001, Benson and Bednarz 2010) and our judgment of relevance to the Swainson's Warbler and likelihood of being affected by flooding (Table 1). Specifically, we estimated cover of forbs, grass, green (combination of forbs and grass cover), litter, understory density, and bare ground within a 5-m radius, and we recorded litter depth and density of cane, vine, shrubs, and total understory stems. Litterdepth (±1 mm) was measured 5 m from each nest location in the four cardinal directions, as was the number of cane, vine, and shrub stems within a 1-m2 quadrat. We estimated density of the understory with a coverboard (Nudds 1977) placed at the plot's center point. An observer recorded the percent of the board covered in five height intervals (0–0.5 m, 0.5–1.0 m, 1.0–1.5 m, 1.5– 2.0 m, and 2.0–2.5 m) from a lateral distance of 11.3 m in each of the four cardinal directions. For analyses, we combined these readings into a mean understory-density variable.
Measurements ( , SE) of habitat and abundance of prey arthropods and results of statistical tests for differences among years within Swainson's Warbler territories at White River National Wildlife Refuge in eastern Arkansas, 2005–2009.
, SE) of habitat and abundance of prey arthropods and results of statistical tests for differences among years within Swainson's Warbler territories at White River National Wildlife Refuge in eastern Arkansas, 2005–2009.
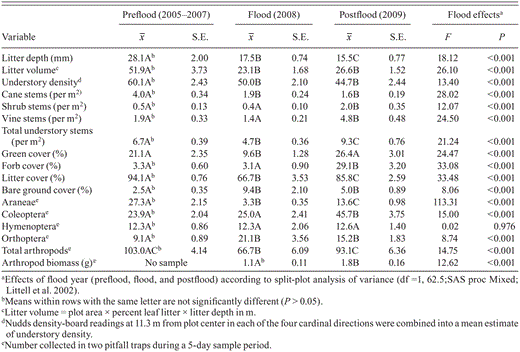
Measurements (, SE) of habitat and abundance of prey arthropods and results of statistical tests for differences among years within Swainson's Warbler territories at White River National Wildlife Refuge in eastern Arkansas, 2005–2009.
Arthropod Sampling
In addition to resampling vegetation characteristics at locations of past nests, we also placed pitfall traps to estimate arthropod abundance and biomass (Brown et al. 2011). Two pitfall traps were placed at randomly chosen paired locations 5 m north and south or east and west from the plot's center. Pitfall traps consisted of a 473-mL plastic cup with the rim of the cup flush with the top layer of soil. We placed a piece of wood from the forest floor, approximately 3 cm wide by 12 cm long, across the top of each cup to deflect precipitation and large debris from entering the cup while allowing arthropods to enter. We filled the pitfall traps with approximately 90 mL of a solution of 50% propylene glycol and 50% water for preserving the samples and collected the traps were after 5 days of sampling. Samples were transferred into labeled whirl-pack bags until they could be transported back to the lab for sorting and identification. When analyzing samples, we identified adult arthropods to order (Tripplehorn and Johnson 2005). We measured biomass to the nearest 0.1 mg after drying the samples in an oven at 50 °C for 48 hr (Duguay et al. 2000).
Data Analyses
Using mixed-model ANOVA (SAS Institute, Cary, NC; proc MIXED; Litteil et al.1996), we examined differences in habitat and arthropod-abundance variables before, during, and after the flood at both occupied and unoccupied territories. To account for potential nonindependence of territories sampled in multiple years, we treated territory as a random effect. We accounted for potentially heterogeneous variances by allowing residual variance to vary among flood categories. We compared the time periods pairwise to identify values significant (P ≤ 0.05) for the main effect.
To determine what habitat and prey-availability variables had the most influence on changes in territory occupancy from preflood to flood to postflood periods, we used multi-season occupancy models (MacKenzie et al. 2006) in program MARK (White and Burnham 1999). Surveys took place up to three times each year, and initial site occupancy for the preflood period was fixed at 1 because all areas of interest were, by definition, occupied by Swainson's Warblers during our previous research (Benson et al. 2009). Because all sites were initially occupied, we focused on factors influencing probability of extinction, the probability a formerly occupied site would become unoccupied.
To evaluate factors influencing extinction probability, we developed a set of a priori models based on factors that we believed to be important from previous research and personal observations (Table 2). To minimize the number of models considered, we first evaluated candidate models for detection probability, then evaluated them for extinction probability by using the best-fit detection model based on Akaike's information criterion adjusted for small sample sizes (AICc). Because we were interested in factors influencing territory abandonment insuccessive years (i.e., extinction probability), we represented all habitat covariates as changes in values between periods (e.g., increase in bare ground cover between preflood and flood years). We ranked candidate models by AICc and computed model weights (Burnham and Anderson 2002). In cases where no model was overwhelmingly supported (wi > 0.9), we used model averaging to examine the effects of explanatory variables on territory abandonment. Prior to fitting models, we evaluated correlations among variables and did not include any highly correlated variables (|r| > 0.70) in the same model.
Results from the best-fitting multi-season models of Swainson's Warbler occupancy as a function of temporal changes in habitat and food abundance at White River National Wildlife Refuge in eastern Arkansas, 2005–2009.

Results from the best-fitting multi-season models of Swainson's Warbler occupancy as a function of temporal changes in habitat and food abundance at White River National Wildlife Refuge in eastern Arkansas, 2005–2009.
Candidate models for variation in detection probability included constant within and among years, varying by year, varying within a season, and varying by year and within a season. Candidate models for extinction probability included only habitat and arthropod-abundance variables that we expected to be most important to Swainson's Warbler from previous habitat-selection studies and those that we expected to have the most influence on site occupancy (Table 2). In particular, these variables included litter volume, litter depth, litter cover, cover of green vegetation, density of shrub, vine, and cane stems, understory density (Graves 2001, Anich et al. 2010a, Benson and Bednarz 2010b), and abundance of spiders (Araneae) and beetles (Coleopterans), prey Swainson's Warblers select (Savage et al. 2010). Because habitat and arthropod abundance changes from the preflood to flood periods were more dramatic than during the flood and postflood periods, we focused primarily on models incorporating changes in habitat and arthropod abundance during this first interval, although some models incorporated changes during both intervals.
Results
Vegetation structure and composition was strongly affected by flooding and varied among the three time periods for most variables (Table 1, Fig. 1). Between the preflood and flood periods, there were significant decreases in green cover, littercover, litter depth, and litter volume, whereas bare ground cover increased (Table 1). Litter depth continued to decrease from the flood to postflood period, and litter cover had not recovered to preflood levels by the postflood year (Fig. 1). Bare ground cover was significantly greater in the flood and postflood years than during the preflood period (Table 1). There were fewer cane stems and total woody stems in the understory, as well as decreased understory density during the flood period than during the preflood period (Table 1). During the postflood year, however, the density of vine stems, shrub stems, and total woody stems in the understory increased significantly over their values during both the preflood and flood periods, although cane-stem density and understory density remained unchanged from the flood to the postflood year (Table 1). Forb cover did not change from the preflood to the flood periods, but it increased significantly by the postflood year (Table 1). Even though, on average, the vegetation structure and composition of territories changed from the preflood to the flood period, the magnitude of change varied dramatically from territory to territory (Table 3).
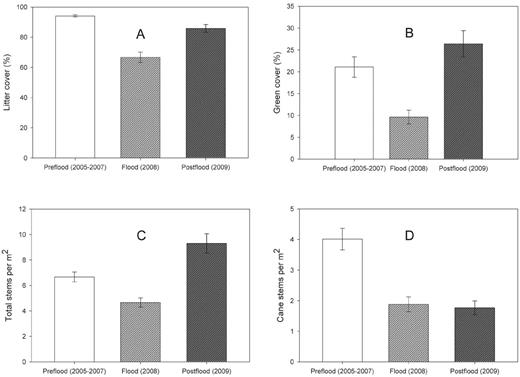
Changes in (A) litter cover (%), (B) green cover (%), (C) total understory stems per m2, and (D) cane stems per m2 in 5-m-radius plots within areas occupied and formerly occupied by Swainson's Warblers at White River National Wildlife Refuge during three periods: preflood years (2005–2007), the flood year (2008), and 1 year postflood (2009). Error bars are ± 1 SE.
There were also substantial changes in arthropod abundance as a result of the flood. Total arthropod abundance decreased from the preflood to the flood year, but postflood numbers were not significantly different from preflood values, indicating that overall abundance of arthropods hadrebounded one year after the flood (Table 1). The abundance of spiders decreased with the flood and recovered somewhat by the postflood year but did not increase to preflood levels (Table 1). Coleoptera were more abundant during the postflood period than during the preflood and flood periods. Oddly, the abundance of Orthoptera increased from the preflood to the flood period. Finally, total biomass of arthropods was significantly lower in the flood year than in the postflood year (Table 1). As stated above, the magnitude of change varied dramatically among individual territories (Table 3).
Ranges of the changes in habitat and prey-abundance variables between the preflood period (2005–2007) and flood period (2008) at White River National Wildlife Refuge in eastern Arkansas.
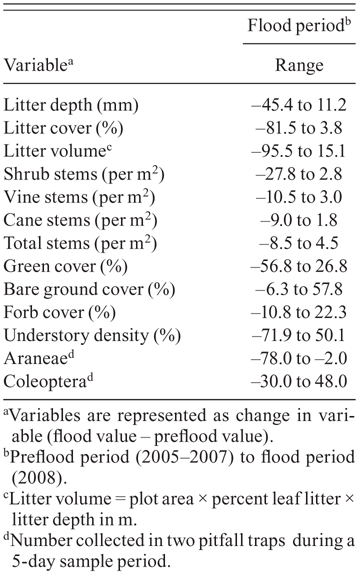
Ranges of the changes in habitat and prey-abundance variables between the preflood period (2005–2007) and flood period (2008) at White River National Wildlife Refuge in eastern Arkansas.
In 2008 and 2009 we resurveyed 42 territories that were occupied in one or more years from 2005 to 2007. Of those surveyed, we detected Swainson's Warblers at 21 (50%) in 2008 and 19 (45%) in 2009. Our best model for probability of Swainson's Warbler detection indicated differences by year and sample period within the postflood year (wi = 0.87). Detection probability was fixed at 1.0 for the preflood period (2005–2007), was 0.64 during the flood year, and was 0.92 and 0.72 for the first and second visits during the postflood year, respectively. Based on model-averaged estimates, occupancy had decreased to 68% (SE = 0.06) and 46% (SE = 0.08) in the flood and postflood years, respectively
Because all territories were occupied during the preflood period, colonization during the first interval was fixed at zero. No territories were colonized between the flood and postflood years. Of the 23 models evaluated, 13 were better predictors of extinction probability than the constant model (Table 2). The top three models indicated that changes in litter cover, green cover, and total understory stems from the preflood to the flood periods influenced the probability of territory abandonment (Table 2); these variables appeared in models that accounted for 84% of the Akaike weight. Because no model was overwhelmingly supported (wi > 0.9), we used model averaging to examine the effects of explanatory variables on territory abandonment. Decreases in litter cover increased extinction probability (Fig. 2a). As green cover increased, extinction probability increased (Fig. 2b). In addition, as total stem density and cane stem density decreased, extinction probability increased (Fig. 2c, d), although densities of cane stems appeared to be of little influence relative to green cover, litter cover, and total stem density. Models containing the remaining variables accounted for only 0.19 of the summed Akaike weight, with no individual variable accounting for more than 0.07 of this summed weight. Moreover, parameter estimates for these variables were near zero, with confidence intervals overlapping zero, indicating that these variables had little influence on territory abandonment.
Discussion
The flooding of our study area had a substantial effect on vegetation structure and composition, the abundance of arthropods, and, ultimately, numbers of Swainson's Warblers. Whilethe effects were dramatic, they were uneven, leading to heterogeneity in habitat change. Along with our previous work in this area, this investigation provided us with an opportunity to examine which factors had the most influence on which territories remained occupied and which were abandoned.
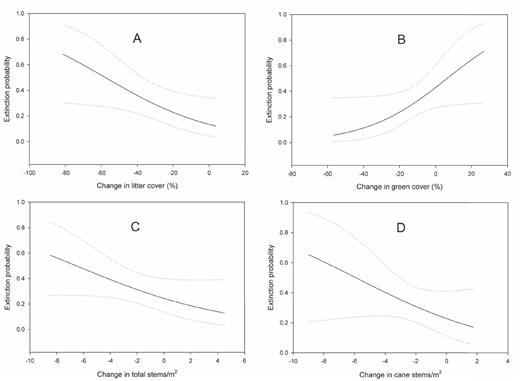
Model-averaged predicted probability of loss (with 95% confidence intervals) of Swainson's Warbler territories as a function of (A) change in litter cover (%), (B) change in green cover (%), (C) change in total understory stems m-2, and (D) change in cane stems m-2from before (2005–2007) to after a major flood (2008) at White River National Wildlife Refuge, Arkansas. We generated estimates while holding all other variables from competing models at their mean values.
Previous studies have reported that Swainson's Warblers avoid standing water during the breeding season or abandon territories in response to prolonged flooding (Meanley 1966, Graves 2001, Klaus 2004). Somewhat contrary to these observations, Anich and Reiley (2010) documented Swainson's Warblers defending flooded territories, and birds continued to use those territories after the flooding had subsided. Nonetheless, Anich and Reiley (2010) observed reduced occupancy consistent with other studies documenting the effects of flooding on ground or understory-dependent birds (Hunter et al. 1987, Knopf and Sedgwick 1987, Knutson and Klaas 1997).
In our study, reduced occupancy by Swainson's Warblers appeared to be the result of changes in habitat, although territories were affected by flooding differently. We observed reductions in litter, cane, shrubs, and arthropod abundance, and increases in bare ground and forbs, changes consistent with previous observations of the effects of floods on habitat structure (Uetz et al. 1979, Yin 1998, Benson and Bednarz 2010). Interestingly, the variables most associated with persistence or abandonment of a territory were not necessarily the same variables previously identified as the most important in influencing habitat use. Specifically, Brown et al. (2009) found density of cane stems the best predictor of presence or absence of Swainson's Warblers, while Benson et al. (2009) found understory density and canopy cover most important for distinguishing nest sites from random locations. Although the reduction in cane-stem density appeared to contribute somewhat to territory abandonment after flooding, understory density was not influential. Change in litter cover, which is known to be important for Swainson's Warblers (Meanley 1971, Brown et al. 2009, Anich et al. 2012), was likely more influential than past studies reported because flooding created a wider range of this variable (preflood 74.5–100.0% and postflood 9.0–98.3%) for birds to select. Similarly, change in the cover of green vegetation greatly influenced territory abandonment; this variable differed at nest and random sites (Benson et al. 2009), but not among used and unused areas within home ranges (Anich et al. 2012) or at occupied and unoccupied sites (Brown et al. 2009). Increased green cover, in the flood and postflood periods, was most likely related to increases in ground-level cover of grasses, forbs, and vines that are consistently and negatively associated with habitat use by this species (Benson et al. 2009, Brown et al. 2009, Anich et al. 2012).
Despite being sensitive to flooding, the Swainson's Warbler is a disturbance-dependent species (Anich et al. 2010b). Even though flooding and other forms of disturbance affect the species negatively in the short term, some form of disturbance is ultimately necessary to create or maintain the dense understory it requires. Longer-term changes related to flooding could improve Swainson's Warbler habitat by stimulating understory growth (Benson and Bednarz 2010). Indeed, floods deposit nutrients in floodplains, set back forest succession, and may stimulate early successional species (Junk et al. 1989). Despite being strongly affected in the flood year, arthropod abundance and most characteristics of understory vegetation had rebounded after one year, with some understory components actually exceeding preflood levels. Though the habitat appeared to be rebounding in some respects, surveys at all previously occupied territories documented further decreases in Swainson's Warbler occupancy (24%; n = 42 preflood territories) in 2010.
One plausible explanation for the lack of postflood recovery in Swainson's Warbler occupancy is the lack of regeneration of cane. Even though Swainson's Warblers use non-cane habitats with the appropriate structure (Meanley 1971, Graves 2001, 2002), cane has historically been the plant that provided the appropriate habitat and nest substrates at our study site (Benson et al. 2009, Brown et al. 2009). During continuing surveys in 2010, there appeared to be fewer cane stems than 1 year after the flood (BMR, pers. obs.). Even though infrequent flooding may promote cane growth (Platt and Brantley 1997), the cane's rhizome networks die when inundated for more than a few weeks (Gagnon 2009); inundation at our site in 2008 exceeded 1 month. Additionally, cane requires canopy openings to benefit from disturbances (Gagnon and Platt 2008). Our study area was characterized by late-successional forest (>90% canopy cover; Anich 2008), and flooding did not change canopy cover appreciably ( = 87%) during the flood and postflood seasons. Alternatively, decreased occupancy after the flood may have resulted from poor site fidelity in response to low reproductive success during the flood and postflood years (Doligez et al. 1999, Hoover 2003). Although we lack data on reproductive success during the flood or postflood periods, the observed changes in habitat may have led to lower nest success than in previous years (Benson et al. 2010) and may have increased emigration.
Less than 26% of the original bottomland hardwood forests of the Mississippi Alluvial Valley remain, and most of the highest-elevation forests have been converted to agriculture (Twedt and Loesch 1999). Additionally, the remaining higher-elevation bottomland forests have been subjected to anthropogenic changes in natural flooding regimes that increase the duration and depth of inundation (Megonigal et al. 1997, Graves 2001, Anich and Reiley 2010). Swainson's Warblers tend to occupy these higher-elevation sites (Brown et al. 2009, Benson et al. 2011) and are often restricted to seasonally inundated buffer zones along rivers and swamps (Graves 2001). Given the rarity of this habitat, these higher-elevation bottomland forest areas should be identified and receive targeted management, especially for understory-dependent birds. Beyond minimizing the effects of floods where possible, management should promote development of a thick understory, as through trees of uneven ages (Twedt and Somershoe 2009). This management, by increasing the light necessary for cane and understory shrubs to respond after a flood, may allow vegetation to be more resistant to flood-related habitat changes.
Acknowledgments
Funding for this research was provided by the U.S. Fish and Wildlife Service. R. Hines, S. Reagan, and D. Sharp of the White River National Wildlife Refuge provided accommodation and access. We thank T. McKay for entomological assistance. We thank N. Anich, C. Roa, A. Zachary, S. Rune, A. Overfield, and A. Beeler for their assistance in the field.
Literature Cited

{kind=link}
{kind=link}