





Abstract.
The Dunlin (Calidris alpina) is a polytypic shorebird with complex patterns of distribution and migration throughout its holarctic range. We analyzed mark-re sighting data obtained between 1977 and 2010 from birds captured at two major staging areas in western Alaska to test the hypothesis that the migration patterns of Alaskan populations are a mixture of parallel and chain, similar to those of Dunlin populations in the western Palearctic. Birds marked on the Yukon—Kuskokwim Delta were found wintering in both Asia and North America, which documented the unexpected mixing of C. a. arcticola from northern Alaska and C. a. pacifica from western Alaska and contradicted our initial prediction of parallel migration pathways for these two subspecies. In its North American winter range C. a. pacifica segregated according to location of marking, confirming our prediction of a chain migration pattern within this population. Individuals of C. a. pacifica marked on the delta were resighted significantly farther north, mostly in southern British Columbia and Washington, than birds marked on the second, more southerly staging area on the Alaska Peninsula, which were resighted primarily in the San Francisco Bay area of northern California. We recommend additional studies use a combination of intrinsic and extrinsic markers to quantify the strength of migratory connectivity between breeding, staging, and wintering areas. Such information is needed to guide conservation efforts because the Dunlin and other waterbirds are losing intertidal habitats at an unprecedented rate and scale, particularly in the Yellow Sea and other parts of Asia.
Resumen.
Calidris alpina es un ave playera politípica con patrones complejos de distribución y migración a lo largo de su rango holártico. Analizamos datos de marca-avistamiento repetido obtenidos entre 1977 y 2010 de aves capturadas en dos grandes áreas de parada en el oeste de Alaska para evaluar la hipótesis de que los patrones migratorios de la población de Alaska son una mezcla de migración en paralelo y en cadena, similar a aquellos de las poblaciones de C. alpina en el oeste paleártico. Las aves marcadas en el Delta de Yukon—Kuskok wim fueron encontradas invernando en Asia y América del Norte, lo cual documentó la mezcla inesperada de C. a. arcticola del norte de Alaska y C. a. pacifica del oeste de Alaska y contradijo nuestra predicción inicial de corredores de migración paralela para estas dos subespecies. En su rango de invernada de América del Norte, C. a. pacifica se segregó de acuerdo a la localización de marcado, confirmando nuestra predicción de un patrón de migración en cadena dentro de esta población. Ejemplares de C. a. pacifica marcados en el delta fueron vistos de nuevo significativamente más al norte, mayormente en el sur de la Columbia Británica y Washington, que las aves marcadas en la segunda área de parada más al sur en la Peninsula de Alaska, las cuales fueron vistas de nuevo principalmente en el área de la Bahía de San Francisco del norte de California. Recomendamos que estudios adicionales usen una combinación de marcadores intrínsecos e extrínsecos para cuantificar la fuerza de la conectividad migratoria entre áreas reproductivas, de parada e invernales. Se necesita esta información para guiar los esfuerzos de conservación debido a que C. alpina y otras aves acuáticas están perdiendo hábitat intertidal a una tasa y escala sin precedentes, particularmente en el Mar Amarillo y otras partes de Asia.
Introduction
Understanding the seasonal geographic structuring of migratory bird populations is fundamental to allocating resources used in conservation efforts. This requires knowledge not only of specific breeding and wintering areas but also of the location and role of other sites used during transit between these seasonally important periods. Understanding migratory connectivity, the degree to which periods of the annual cycle are geographically linked within a population, is crucial for determining where and when a population is limited (Webster et al. 2002, Boulet and Norris 2006). Migratory shorebirds in particular present challenges in assessing connectivity primarily because species have such varied migration strategies (Piersma 1987, Colwell 2010, Warnock 2010). These differ by distance flown, number of stopover or staging sites used, season, age, sex, and population. Assessing migratory connectivity and population structure in the face of such variation is made even more challenging by the fact that many shorebird species nest in remote regions of the north that are difficult to access and where birds are dispersed over large areas in relatively low numbers.
The Dunlin (Calidris alpina) is a well-studied migratory shorebird with a circumpolar breeding range. It winters mostly in estuaries of the North Temperate Zone (Cramp and Simmons 1983, Piersma et al. 1996, Warnock and Gill 1996, Fernández et al. 2010). The species exemplifies the challenges faced in assessing seasonal connectivity in migratory birds because of its high level of geographic structuring on the breeding grounds and its complex migration pathways. Molecular studies of mitochondrial DNA (mtDNA) have revealed strong evidence of five major phylogeographic groups across the species' breeding range, lineages that reflect historical isolation of populations in tundra refugia during the late Pleistocene Epoch and a more recent divergence within the Beringian region of the North Pacific Basin (Wenink et al. 1996, Buehler and Baker 2005).
Finer geographic structuring among breeding populations of the Dunlin has been documented through morphometrics and plumage characteristics, although the number of subspecies that should be recognized (up to 11) is disputed (cf. Todd 1953, MacLean and Holmes 1971, Browning 1977, 1991, Greenwood 1986, Tomkovich 1986, Nechaev and Tomkovich 1988, Engelmoer and Roselaar 1998). Dunlins maintain strong site fidelity to both breeding and natal areas (Holmes 1966, Soikkeli 1967, 1970, Tomkovich 1994), and this behavior is thought to have contributed to the high level of differentiation among populations (Lappo 1996, Wenink et al. 1996). Multiple studies of Dunlins breeding in the western Palearctic have shown strong population structuring along migration routes and on wintering grounds, although mixing of breeding populations does take place (Greenwood 1984, Wenink and Baker 1996, Wennerberg 2001, Lopes et al. 2006). A system of parallel migration (sensu Salomonsen 1955) appears to be the general pattern among western palearctic Dunlins, with aspects of chain migration and leap-frog migration in different parts of the species' range (Wennerberg 2001). Migratory connectivity has significant implications for conservation at various spatial scales, ranging from specific estuaries to entire flyways (Lopes et al. 2006).
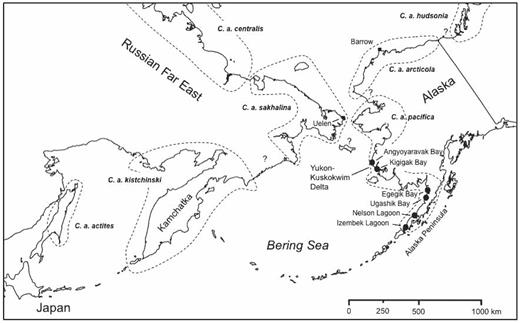
Beringia and the North Pacific region showing the principal study areas in Alaska (Yukon—Kuskokwim Delta and Alaska Peninsula) and sites within each (filled circles) where most Dunlins were captured and marked between 1976 and 2010 (details in Table 1). Dashed lines enclose the general breeding ranges of the various subspecies of the Dunlin that nest in the region (after Lappo and Tomkovich 1998).
Although the population structuring of Dunlins breeding in the western Palearctic has been well studied, much less is known about such structuring among Dunlins breeding in Alaska and the Russian Far East. Five subspecies based on morphometric and plumage characteristics are recognized from this region, including C. a. arcticola in northern Alaska, C. a. pacifica in western Alaska, C. a. sakhalina in Chukotka, C. a. kistchinski on the Kamchatka Peninsula, and C. a. actites on northern Sakhalin Island (Fig. 1). No mtDNA haplotypes from the Russian Far East or Alaska have been found among birds migrating or wintering in the western Palearctic (Wenink and Baker 1996, Wennerberg 2001), nor is there evidence of intracontinental genetic mixing between Dunlins of Alaska and the central Canadian Arctic in either their breeding or winter ranges (Wenink et al. 1996). These findings suggest that Dunlins breeding in Alaska and the Russian Far East are restricted to migration flyways and winter ranges in the Pacific basin, where Dunlins occur at temperate latitudes in both North America and Asia (summarized in Piersma et al. 1996).
Migratory connectivity within the Pacific basin is less clear, however. Samples of nonbreeding Dunlins from both sides of the basin contain a mixture of haplotypes from breeding areas in northern and western Alaska and the Russian Far East (Wenink and Baker 1996). The few recoveries of banded birds connecting breeding and nonbreeding grounds in this region linked northern Alaska to eastern Russia (n = 5) and Taiwan to Sakhalin Island (n = 1; Norton 1971, Greenwood 1984). Analyses of plumage and morphometric characteristics suggest that Dunlins breeding in northern Alaska and the Russian Far East share common migration routes to nonbreeding areas in eastern Asia, whereas those breeding in western Alaska migrate to nonbreeding grounds along the Pacific coast of North America (MacLean and Holmes 1971, Tomkovich 1986). Such analyses were complicated by the subspecies' substantial overlap in measurements and the lack of characteristic differences in basic plumage (Tomkovich 1986).
In 1976, we began a study in western Alaska of the autumnstaging and migration ecology of Dunlins of the subspecies C. a. pacifica, which nest in coastal habitats throughout western Alaska, are common there on intertidal areas during autumn, and spend the winter mostly along the Pacific coast of North America south of Alaska (in Warnock and Gill 1996, Page et al. 1999). Our initial objective was to determine the importance of coastal habitats in Alaska to autumn-staging Dunlins by assessing the abundance, habitat use, and temporal occurrence of birds across broad geographic areas, and these results have been reported previously (e.g., Gill and Jorgensen 1979, Gill and Handel 1990, Handel and Gill 1992). Our second objective was to establish links between areas where Dunlins stage in western Alaska and sites where they winter through analysis of resightings and recoveries of birds marked during our multiple studies in the region. A recent impetus to continue this research was to assess the prevalence and potential pathways of intercontinental transmission of avian influenza viruses among migratory birds (Ip et al. 2008). We hypothesized that migration of Alaskan Dunlins should reflect the general pattern of parallel migration of populations in the western Palearctic (Wennerberg 2001), with C. a. pacifica following an eastern flyway within the Pacific basin, segregated from the western flyway of C. a. arcticola. We also predicted that, like other Dunlin populations with longitudinal migration routes (Wennerberg 2001), C. a. pacifica should follow a pattern of chain migration, with birds from more northerly staging areas wintering farther north along the Pacific coast of North America than those from more southerly staging areas.
Study Area
Our study encompassed two areas of western Alaska: (1) the outer coast of the central Yukon—Kuskokwim River delta (YKD) and (2) the northern coast of the Alaska Peninsula (AP), which lies about 700 km SSW of the YKD (Fig. 1). Both the YKD and AP are characterized by numerous large estuaries and embayments (50–100 km2) of extensive intertidal mud and sand flats. Dunlins use these intertidal areas between July and late October (Gill and Jorgensen 1979, Gill and Handel 1990, Handel and Gill 1992) after nesting on nearby graminoid meadows (Holmes 1971, Warnock and Gill 1996). Over the study region we captured and marked Dunlins staging in autumn at two principal sites, Angyoyaravak Bay (YKD; 61° 16′ N, 165° 40′ W) and Nelson Lagoon (AP; 56° 00′ N, 161° 07′ W). Lesser numbers were captured and marked in 1995 at another YKD site (Kigigak Island; 60° 51′ N, 165° 01′ W) and at three other large estuaries along the AP (Fig. 1; Table 1): Izembek Lagoon (55° 18′ N, 162° 56′ W) in 1993 and 1994, Ugashik Bay (57° 29′ N, 157° 35′ W) in 2006, and Egegik Bay (58° 12′ N, 157° 31′ W) from 2004 to 2006. During these and intervening periods observers were present at several other western Alaska estuaries (Fig. 1), where they attempted to locate marked Dunlins.
Number of Dunlins captured by site, year, and type of marking, 1977–2010.
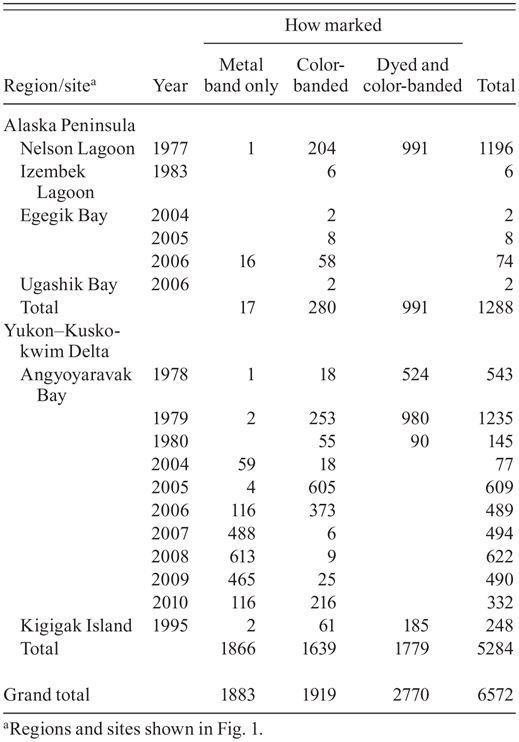
Number of Dunlins captured by site, year, and type of marking, 1977–2010.
Methods
Capture and Marking
During the post-breeding period (mid-July to mid-October) we captured Dunlins primarily by using propelled nets (rocket or bungee) and mist nets placed at or near high-tide roosts that form annually along the immediate coast (Handel and Gill 1992); a much smaller number of birds was captured at these roosts with walk-in traps and hoop nets. Efforts at capture were fairly constant at all sites, averaging about once every 2–3 days, but gaps of 10–15 days arose when birds shifted use of roost sites because of periodic changes in tidal regimes and adverse winds (Handel and Gill 1992).
Each captured bird received a numbered metal band. Most birds received one or more additional markers, including various colors of plastic leg bands, colored leg flags (plain or engraved) and picric acid applied as a dye to the plumage of the breast, flanks, or vent (Table 1). These special markers were applied in combinations unique to the year and location of banding; some birds received additional marks according to age (adult or juvenile), according to period of banding (usually changed at 2- to 3-week intervals), and, later in the study, according to putative subspecies (see beyond). Although having dyed plumage increases a bird's chance of being detected away from the site of capture (Handel and Gill 1983), we stopped applying picric acid in the mid-1990s because we suspected it increased risk of predation by gulls and jaegers. For most birds captured (>95%) we recorded mass to the nearest 0.5 g, wing length to the nearest 1 mm, and exposed culmen to the nearest 0.1 mm. We scored fat levels on a modified version (scale 0–5) of the scale of Prater (1975) and aged the birds by plumage (in Warnock and Gill 1996, Choi et al. 2010).
Statistical Analyses
To assess the occurrence of marked Dunlins away from capture sites we analyzed both recoveries (a bird was recaptured and the band number was recorded) and resightings (a bird was seen and all or part of its band combination was recorded). Recovery data were obtained from the U.S. Geological Survey's Bird Banding Laboratory (current through January 2012) and included all birds banded by us and recovered elsewhere plus birds banded elsewhere and recovered at our study sites. Data on resightings included records of birds we marked that were seen away from Alaska plus birds marked by others in the winter range and seen by us at our Alaskan study sites.
We chose a conservative approach in compiling our database of resightings, especially in dealing with marked birds seen away from the capture site. Our main concern here was how to treat potentially duplicate sightings of individuals since most of our birds could be identified only to cohort (place and period of capture). Where we had multiple records of a particular combination of markers within 20 km of each other and on different dates, we treated these as a single bird. If two or more similarly marked birds were seen simultaneously by a single observer or by separate observers in different portions of an embayment about the same time on a given day, we treated these as different individuals.
We considered records from first arrival in late September to November to represent late-autumn distribution and those from December to March to represent winter distribution. We used Mann—Whitney U tests to evaluate differences in the latitudinal distribution of resightings and recoveries on the wintering grounds between late September and March (1) between birds marked on the two staging areas (YKD vs. AP) or (2) between autumn and winter for either group. To examine distributional patterns of resightings and recoveries of marked birds in the winter range, we considered all reports from April to May separately because birds may have already begun northward migration (Page 1974, Warnock et al. 2004, Brazil 2009). We present summary statistics of latitudinal distribution as medians and interquartile ranges (IQR; 25–75%).
Results
Capture and Marking Effort
Between 1977 and 2010 we captured and marked 6572 Dunlins during 13 different field seasons (Table 1); 4689 (71%) of these were color-banded (some also dyed) and thus were more likely to be seen than a bird marked only with a metal band (Handel and Gill 1983). Among the color-marked birds about 73% were from the YKD and 27% from the AP (Table 1). Most birds marked on the AP were adults (93%), while less than half (42%) of those marked on the YKD were adults. From the pool of color-marked birds (4689) we received 133 reports of our birds seen away from Alaska (through 2011). After we eliminated potentially duplicate records of the same individuals, the number was reduced to 66, for a minimum rate of resighting of 1.4%. An additional 12 of our banded birds were recovered by others away from Alaska. We also recorded at our Alaskan study sites 18 Dunlins that had been marked by others away from the breeding range (6 recoveries and 12 resightings). Using all of these categories combined, we had records of 96 birds from which to assess migratory connectivity between our staging areas in Alaska and nonbreeding areas elsewhere.
Distribution of Resightings and Recoveries
Two distinct distributional patterns emerged from the 66 resightings and 12 recoveries of birds we banded in western Alaska (Table 2). First, Dunlins that we marked on the YKD were reported from both North America (n = 37; 21 September–23 May) and Asia (n = 24; 13 October–20 May), whereas Dunlins we marked on the AP were seen only in North America (n = 17; 13 October–23 February). The second pattern to emerge was a distinct difference in latitudinal distribution along the Pacific coast of North America between birds marked on the two staging areas (Fig. 2). During both late autumn and winter, birds from the YKD recorded in North America occurred significantly farther north (median = 49.0° N; IQR = 3.2°; n = 33) than those from the AP (median = 38.0° N; IQR = 3.8 °; n = 17; U = 70.0; P = 0.03). We found no evidence of a change in latitudinal distribution between late autumn and winter for birds from either the YKD (U = 129.5, n1 = 19, n2 = 17, P = 0.31) or AP (U = 29.5, n1 = 12, n2 = 7, P = 0.29). Dunlins from the YKD were reported from southeastern Alaska to Baja California, Mexico, but were clustered in southern British Columbia and northern Washington. Reports of AP birds were concentrated around the San Francisco Bay area of northern California but ranged from northern Washington to Monterey Bay, California. We had no records of AP birds during spring (April–May), but six YKD birds were reported along the coast of North America between Oregon and southeastern Alaska during this period, including three that had also been recorded at least once at those sites during winter.
Number of Dunlins marked at staging sites in western Alaska and resighted or recovered in their winter range in North America and Asia, and of those marked in the winter range and recovered or resighted at our Alaskan study sites, 1977–2010.

Number of Dunlins marked at staging sites in western Alaska and resighted or recovered in their winter range in North America and Asia, and of those marked in the winter range and recovered or resighted at our Alaskan study sites, 1977–2010.

Distribution of records (sightings and recoveries) along the Pacific coast of North America of Dunlins linked to capture efforts on the Yukon—Kuskokwim Delta (YKD; left panel) and the Alaska Peninsula (AP; right panel). Insets show expanded views of areas where records were concentrated. Filled circles = locations where birds marked on the YKD or AP were sighted away from Alaska; unfilled circles = locations where birds marked on the YKD or AP were recovered away from Alaska; unfilled triangles = locations where marked birds were seen away from Alaska in spring (April–May); filled squares = locations away from Alaska at which birds were marked by others and seen in Alaska; unfilled squares = locations away from Alaska at which birds were marked by others and recovered in Alaska. Numbers within or adjacent to symbols denote multiple records.
The Dunlins marked on the YKD and reported from Asia (Fig. 3) were distributed from northern Sakhalin Island, Russia, south to Taiwan. Unlike the pattern along the coast of North America, however, resightings of birds in Asia shifted significantly southward between late autumn, when the distribution was centered near Japan and the Republic of Korea (median = 35.2° N; IQR = 4.5°; n = 10), and winter, when the distribution was concentrated closer to Taiwan (median = 26.5° N; IQR= 11.8°; n = 11; U = 25.0; P = 0.04). Six resightings during May (five in the Republic of Korea and one in Japan) indicated a subsequent northward shift.
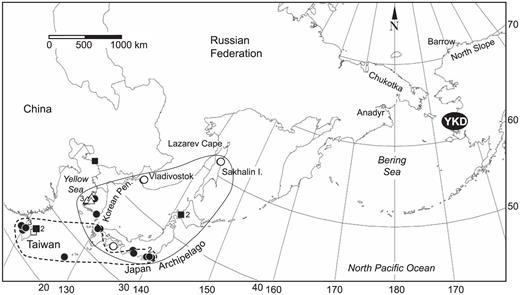
Distribution in Asia of records of Dunlins linked to the Yukon—Kuskokwim Delta, Alaska. Filled circles = locations where birds marked on the YKD were sighted away from Alaska; unfilled circles = locations where birds marked on the YKD were recovered away from Alaska; unfilled triangles = locations where marked birds were seen away from Alaska in spring (April–May); filled squares = locations away from Alaska at which birds were marked by others and seen on the YKD; unfilled squares = locations away from Alaska at which birds were marked by others and recovered on the YKD. Numbers within or next to symbols denote multiple records. Lines encircling regions depict where marked birds were recorded during autumn (September–November; solid line) and during winter (December–March; dashed line).
Distributional patterns of Dunlins banded elsewhere and recorded at our study sites in Alaska were similar to those of our marked birds. On the AP, we recorded four marked Dunlins at Nelson Lagoon and five at Izembek Lagoon, all of which had been banded in northern California. On the YKD, however, we recorded one Dunlin from mainland China, three from Taiwan, two from Japan, one from British Columbia, and two from Oregon (Figs. 2 and 3).
Discussion
Alaska-Marked Birds Seen in Asia
The numerous resightings in Asia of Dunlins marked on the YKD were unexpected because the delta lies within the core breeding range and sites of staging of C. a. pacifica—a population thought to winter only in North America (Todd 1953, MacLean and Holmes 1971, Browning 1977, 1991). This meant either that some portion of the population of C. a. pacifica migrates to Asia, contrary to MacLean and Holmes' (1971) morphometric assessment, or that birds from another population move to western Alaska to stage with C. a. pacifica. Mixing of populations, whether in the winter range or at staging sites, contradicted our initial hypothesis of segregated, parallel migrations within the Pacific basin.
Recent evidence confirms that at least the subspecies C. a. arcticola from northern Alaska occurs among Dunlins staging on the YKD in autumn: (1) 7 Dunlins (4 adults, 3 juveniles), among 24 fitted with VHF radios at nests (adults) or along the coast (juveniles) in northern Alaska in 2005 were detected on the central YKD between 26 August and 7 October that year (Taylor et al. 2011), and (2) in 2008 and 2009 we recorded 4 Dunlins on the YKD (1 recapture, 3 resightings) that had been individually color-marked on the North Slope of Alaska (R. Lanctot and R. Gill, unpubl. data). Although we have no evidence of C. a. sakhalina staging on the YKD, we cannot at this time preclude that some may also occur there. Tomkovich (1986) identified as C. a. pacifica several “long-billed” Dunlins nesting sympatrically with C. a. sakhalina at Uelen, Russia, only 90 km across the Bering Strait from the breeding grounds of C. a. pacifica in Alaska (Fig. 1). Thus it is plausible that some examples of C. a. sakhalina could migrate with C. a. pacifica from Russia to stage on the YKD with C. a. arcticola from northern Alaska and locally breeding C. a. pacifica. Mixing of populations staging on the YKD may provide a mechanism for Dunlins with Beringian haplotypes (breeding only in Asia; Wennerberg 2001) to winter in both Asia and North America (Wenink and Baker 1996) despite the apparent lack of gene flow. Analysis of additional genetic markers may reveal whether the sharing of two of seven Alaska haplotypes by C. a. pacifica and C. a. arcticola (Wenink and Baker 1996) is due to current gene flow or to incomplete lineage sorting of mtDNA markers among these most recently differentiated populations (cf. Pruett and Winker 2010).
Our data confirm that some Dunlins wintering in Asia stage on the YKD as early as mid-August, and some radiotagged birds from northern Alaska have been detected there until 7 October (Taylor et al. 2011), but we do not yet know what proportion of the population stages on the YKD or elsewhere in western Alaska. Our findings and the few recoveries of C.a. arcticola banded in its breeding range in northern Alaska (Norton 1971, North American Bird Banding Laboratory data) suggest that C. a. arcticola migrates south in at least two stages, the first being a potentially nonstop flight to sites in southeast Russia, Japan, and Korea, regions of the earliest (October and November) resightings and recoveries of our marked birds. By early August foods on northern Alaska tundra are in decline (Holmes 1966), and Dunlins shift feeding to intertidal areas, where they then become uncommon by early September (Connors 1984, Taylor et al. 2010). The rich intertidal flats of the YKD attract large numbers of shorebirds, not only from local breeding populations but also from areas farther north throughout Alaska and Russia, to prepare for trans-Pacific migrations (Gill et al. 1994, 2009, Alerstam and Gudmundsson 1999, Handel and Gill 2010, Lindström et al. 2011, Battley et al. 2012).
Our analysis of Alaska birds seen in Asia suggested a directed southward movement after marked birds arrived there. The movement of birds south from the coasts of Russia and northern Japan corresponds with the onset of freezing temperatures in those areas beginning in December (http://www.worldclimate.com/). We cannot discount, however, that the distribution of our resightings may have suffered from a spatiotemporal bias in search effort in Asia (cf. Lanctot et al. 2009) or reflected annual variation or anthropogenic change to habitats through our study. Assessing the movements in Asia and defining the bounds of the winter range of C.a. arcticola is particularly challenging because at least three other subspecies of the Dunlin (kistchinski, sakhalina, actites) are thought to share winter ranges along the central coast of eastern Asia (Tomkovich 1986, Bamford et al. 2008, Lanctot et al. 2009, Xiangyu et al. 2009, Choi et al. 2010; Z.-J. Ma, pers. comm.). Studies in progress using stable isotopes and light-level geolocator technology (R. Lanctot, pers. comm.) on populations of all nearctic subspecies (arcticola, pacifica, hudsonia) of the Dunlin should help answer these questions.
Wintering Area Segregation in North America
The second major finding of our study was that C. a. pacifica segregated in its winter range, as we predicted, largely according to where the birds were marked, with those from the YKD, the more northerly staging area, residing throughout winter on average much farther north than birds from the AP If the birds stage at areas near tundra where they breed, this pattern implies chain migration, which is predicted to evolve when the cost of migration is related to distance or when there is an advantage of early arrival in breeding or wintering areas and a strong latitudinal gradient in suitability of breeding or wintering habitat (Lundberg and Alerstam 1986). The origin of all Dunlins at the two staging areas, however, is not completely certain. Movement of some proportion of C.a. arcticola from northern Alaska to the YKD suggests that this rich staging area might also draw C. a. pacifica from areas north of the YKD as well as locally. An ancillary study of Dunlins radio-marked as they staged on the YKD showed that a small proportion (9%) moved to estuaries of the AP before migrating to wintering grounds, with juveniles having a greater tendency to drift southward than adults (Warnock et al. 2013). Limited mixing on staging areas could account for the incomplete segregation of marked birds in the winter range, including Dunlins from the AP seen in Oregon and Washington and birds from the YKD seen south of Oregon.
Unlike Dunlins migrating to Asia, those wintering in North America were largely resident after arrival until the return migration in spring. The distribution of our resightings mirrored the winter distribution found along the Pacific coast of the contiguous United States, with concentrations in Washington—Oregon estuaries and in San Francisco Bay (Page et al. 1999). Other studies have also reported little winter movement other than at local scales (Brennan et al. 1985, Buchanan 1988, Butler 1994, Warnock et al. 1995, Sanzenbacher and Haig 2002). Buchanan et al. (1986), however, did note influxes of immature birds into Grays Harbor, Washington, in November and concluded that some immatures have a protracted southward migration in November.
Conservation Implications
Knowing how migratory populations are geographically linked between seasonal periods is fundamental to understanding not only basic processes such as population regulation but also where and when in the annual cycle conservation measures should be implemented (Webster and Marra 2005, Boulet and Norris 2006, Martin et al. 2007). In different portions of their range, particularly wintering and stopover sites, Dunlins are currently being subjected to serious perturbations, most human-caused, that could induce significant populationlevel changes. In North America these include (1) loss of intertidal habitats to mariculture in Mexico, (2) restoration of salt ponds to vegetated salt marsh in Mexico and California, (3) spread of exotic Spartina spp. onto intertidal flats in California, Washington, and British Columbia, (4) human disturbance of roost sites, and (5) changing agricultural practices (Fernández et al. 2010, Sutherland et al. 2012). In Asia, by far the most immediate and serious threat is the reclamation of intertidal habitats throughout the Yellow Sea for industrial expansion, which is occurring at an unprecedented pace and scale (Barter 2002, Birds Korea 2010, Rogers et al. 2010, Yang et al. 2011, MacKinnon et al. 2012). This monumental loss of intertidal habitat will affect many species of waterbirds, including the Dunlin, that rely on this globally important area for migration and wintering, and conservation measures that address habitat loss in the Yellow Sea are being discussed at the international level.
Quantitative frameworks of economic modeling can be used to optimize strategies for protecting habitat along entire flyways (e.g., Klaassen et al. 2008) and throughout the annual cycle (e.g., Sheehy et al. 2011). Such models require information on the cost to protect sites used by the population as well as the relative quality, importance, and interdependence of those sites. Our study has documented linkages between staging areas in western Alaska and wintering areas along the Pacific coast of North America for C. a. pacifica and of the importance of the YKD as a staging area for C. a. arcticola, which winters in Asia, but quantifying the strength of these linkages will require larger sample sizes. Now available for small birds are inexpensive miniaturized light-level geolocators that can track their movements throughout an annual cycle, potentially providing a more robust and less biased assessment of migratory connectivity than do banding studies. Data from geolocators, however, need to be physically recovered from the tags, and, because latitudinal estimates are based on estimates of day length, locations can be difficult to determine with accuracy at high latitudes, around solar equinoxes, and under variable environmental conditions (Lisovski et al. 2012), reducing their utility for shorebirds like the Dunlin that breed in the Arctic and migrate near the autumnal equinox. Although resighting and recovery data from banding studies are generally sparse, they still provide the most spatially accurate information on bird migration. We recommend a combination of extrinsic and intrinsic markers be used to determine: (1) what proportion of C.a. arcticola stages on the YKD before continuing on migration to Asia; (2) the magnitude and duration of use of stopover and wintering sites in Asia, including the Yellow Sea, by C. a. arcticola and other subspecies of the Dunlin breeding in the Russian Far East; and (3) the sites of breeding of C. a. pacifica wintering in southern California and Mexico. Quantifying the strength of such linkages at the population level will assist in optimizing conservation efforts for this species.
Acknowledgments
A study extending over four decades was obviously supported by many people and institutions. Financial support for field work from 1976 to 1982 was provided to the U.S. Fish and Wildlife Service (USFWS) through the Bureau of Land Management as part of the Alaskan Outer Continental Shelf Environmental Assessment Program, from 1983 to 1996 by the USFWS, and from 1997 to 2010 by the U.S. Geological Survey (USGS), the “Beringia 2005” expedition of the Swedish Polar Research Secretariat, and the interagency (USFWS and USGS) program for surveillance sampling for highly pathogenic avian influenza. Principal logistical support was provided by the Yukon Delta and Izembek national wildlife refuges. Numerous individuals helped with capturing and marking Dunlins over the years, but we owe special recognition to P. Battley, F. Broerman, J. Conklin, S. Davis, T. DeGange, M. Dementyev, T. Donnelly, M. Green, C. Harwood, Å. Lindström, J. Lawonn, H. Lemke, M. Maftei, B. McCaffery, C. McCaffery, D. Melville, J. Nelson, M. Petersen, A. Riegen, M. Sexson, D. Stojanovic, J. Terenzi, L. Tibbitts, and D. Veitch. We especially thank C.-Y. Chiang, J. Choi, P. Sanzenbacher, and N. Warnock for allowing us to incorporate sightings of their marked birds into our database. Equally important to marking birds was having them resighted. The late Mark Barter maintained a database of Dunlins resighted away from or in east Asia, an invaluable cross-reference for our records. The following we thank for their observations or for passing along observations by others, with names in italics responsible for multiple records: D. Aldercroft, N. Arce, P. Ashman, R. Bayer, the late L. Binford, J. Buchanan, E. Cardiff, R. Carmona, T. Chandik, C. Hsuan, C.-Y. Chiang, J. Choi, P. Connors, L. Cooper, G. Dorsey, N. Drumheller, A. Edwards, M. Egger, R. Erickson, J. Evens, S. Feys, R. Gates, H. Gibbs, R. Gordon, T. Harris-Haller, S. Harrison, Mr. Hattori, N. Hentze, S. Herman, B. Hill, T. Hiroi, M. Hyakutake, J. Ireland, K. Irwin, S. Jamieson, H. Johansen, S. Johnston, Y.-K. Ju, G. Kaiser, M. Kashiwagi, N. Kishimoto, A. Kumagai, Y. Kurasashi, R. Lanctot, P. La Tourrette, J. Lawonn, R. LeValley, B. MacDonald, M. Mahafey, N. Mamagata, T. Manoles, J. Michaels, C. Minton, J. P. Myers, T. Nakayama, H. Nehls, P. Nietlisbach, G. Page, C. Park, D. Paulson, D. Roberson, D. Rogers, R. Ronson, S. Shanewise, Y. Shigeta, D. Shuford, J. Smith, W. Somerville, P. Springer, L. Stenzel, E. Stopps, P. Suchanek, K. Suzuki, Mr. Takenaka, W. Turnbull, G. Van Vliet, T. Wahl, R. Widrig, K. Wohl, L.-L. Wu, and D. Yu. We thank R. Browning, J. Choi, Z.-J. Ma, P. Tomkovich, and N. Warnock for sharing information and insights. J. Terenzi worked his magic with ArcGIS, D. Bystrak provided data from the USGS Bird Banding Laboratory, and P. Unitt made available historic references. Use of trade names is for descriptive purposes only and does not imply endorsement by the U.S. government. We thank L. Tibbitts, R. Lanctot, J. Pearce, T. Piersma, and two anonymous reviewers for helpful comments.
Literature Cited

{kind=link}
{kind=link}
{kind=link}