





Abstract.
Marine radar is a tool widely used in the study of bird migration, but in most cases it cannot identify detected objects to species. For this reason airspeed is used as one of the main criteria to separate birdlike from insectlike targets, but this criterion has not been tested outside the temperate regions where it was developed. We used a theoretical approach for calculating minimum-power speeds (Vmp) and maximum-range speeds (Vmr) of birds and insects to estimate an interval of airspeeds from which an appropriate value of the criterion can be selected and to propose a criterion based on flight speed for the Isthmus of Tehuantepec in southern Mexico. This approach, with data from 36 species of insects and 60 of passerines either known to occur or similar to species occurring in southern Mexico, found that although Vmp of insects and birds overlaps at 5.5 m sec-1, birds generally fly faster than insects. After combining our results with data from the literature, we conclude that the best airspeed to be used as criterion for distinguishing birdlike from insectlike targets lies within the range of 5.5–9.0 m sec-1 and that 7 m sec -1 is an appropriate airspeed for our study area. These airspeeds are applicable in southern Mexico and in regions where birds and insects have physical dimensions similar to the species addressed in our study.
Resumen.
Los radares marinos son una herramienta ampliamente utilizada para estudiar la migración de aves, pero son polémicos debido a que en la mayoría de los casos no permiten identificar objetos al nivel de especie. Por esta razón, la velocidad aerodinámica de vuelo es utilizada como uno de los principales criterios para separar blancos tipo ave de blancos tipo insecto. Sin embargo, tal criterio no existe para areas fuera de las zonas templadas donde dicho criterio fue desarrollado. Utilizamos un enfoque teorico calculando las velocidades de potencia minima (Vmp) y de alcance máximo (Vmr) de aves y de insectos para estimar un intervalo de velocidades aerodinámicas de vuelo del que pueda elegirse un valor para ser usado como criterio, asi como para proponer un criterio basado en velocidades aerodinámicas para el Istmo de Tehuantepec, en el sur de México. Este enfoque, con datos de 36 especies de insectos y 60 de aves Passeriformes presentes en el Sur de Mexico, o de especies similares a las de esa zona, encontró que aunque la Vmp de aves y de insectos se superpone a 5.5 m sec-1, las aves generalmente son más veloces que los insectos. Después de combinar nuestros resultados con datos disponibles en la literatura, consideramos que la mejor velocidad aerodinámica para ser usada como criterio para separar a las aves de loes insectos se encuentra entre 5.5 y 9.0 m sec-1 y que 7 m sec-1 es una velocidad adecuada para nuestra área de estudio. Estas velocidades son aplicables en el sur de México y en regiones cuya fauna aviar y entomológica tenga dimensiones físicas similares a las de las especies usadas en el presente estudio.
Introduction
Marine surveillance radars have proven to be useful tools for studies of nocturnal bird migration (Cooper et al. 1991, Williams et al. 2001, Harmata et al. 2003, Mabee et al. 2006). They have been used extensively in ornithological research because of their portability, low power requirements, and cost. However, one of the biggest challenges to the widespread application of this tool is to differentiate birds, bats, and insects (Larkin 2005). For this reason the terms “vertebrate-like target,” “birdlike target”, “insectlike target,” or just “target” are commonly used to describe detected objects, in recognition of this difficulty. Nevertheless, some criteria are applicable, and airspeed is a variable that partially distinguishes birds and insects (Larkin 1991, Zaugg et al. 2008), because typical flight speeds of many insects are 2–4 times slower than those of birds (Ellington 1991). Bats' flight speeds, however, overlap with those of passerines (Hedenström et al. 2009), so bats cannot be separated out solely by airspeed and require an additional or different criterion (e.g., flight pattern).
It has been proven that slow-flying targets detected with radar are not birds but insects (Larkin 1991); following this, some authors use an airspeed “threshold” of 6 m sec-1 (Diehl et al. 2003, Mabee et al. 2006, Ruth et al. 2010), considering slower targets to be “insectlike” and faster targets to be “birdlike.” However, the data on which that criterion is based are from temperate regions, where insect diversity differs considerably from that in the tropics, and flight speeds may differ as well. For these reasons, the temperate-region criterion may not be applicable to tropical regions where radar is being used in the study of bird migration. In our studies of the migration of birds through the Isthmus of Tehuantepec in Oaxaca, Mexico (not reported here), we realized the need for a criterion that incorporated the regional knowledge of insects for adequate interpretation of radar studies in a tropical environment. Our objective in this paper is to describe the methods used to define a range of airspeeds that can be used as criterion to distinguish insects from birds in our study area and to show that these airspeeds can be used for other regions.
Airspeeds of birds and insects can be measured directly in the field and then compared. However, theoretical calculations can also be made on the basis of mechanical and aerodynamic principles that apply to all animals that support their weight aerodynamically (Pennycuick 1997). This approach states that mechanical power available in flight muscles is limited, placing both lower and upper limits on the speed any bird can fly. The lower speed, called “minimum power speed” (Vmp ), is the speed a bird can maintain by using the minimum amount of metabolical energy; the upper, called “maximum range speed” (Vmr), is the speed when distance traveled is maximized by unit of work done. Radar studies of the Common Swift (Apus apus) have proven the existence of these flight speeds (Bruderer 1997), which can be obtained through the calculation of the so called “power curves” (Rayner 1999).
These theoretical speeds have been compared with field data from both passerines and nonpasserines, and some similarities between them and observed flight speeds have been found. For instance, Welham (1994) compared flight speeds reported in literature for 48 avian species with predictions of Vmr and found that ∼40% of them generally migrated at that theoretical speed, but Pennycuick (1997) suggested that migrating at Vmr may not represent an “optimal” or even a practical strategy, reporting that the mean airspeeds of 36 species (both passerines and nonpasserines) were distributed around Vmp . Pennycuick (2001) found that two species of passerines that fly by bounding had mean speeds of 1.70Vmp and 1.96E but other species with continuous flapping flew between 0.82 and 1.27Vmp. Although this approach has been developed to analyze birds' flight performance, it can also be applied to insects (Srygley 2003, Pennycuick 2008, Srygley and Dudley 2008, Tennekes 2009). Vmp and Vmr represent a distribution of potential flight speeds, with minimum and maximum flight speeds lying at the limits of this distribution. This theoretical approach provides a range of possible airspeeds that could be used to distinguish insectlike and birdlike targets detected by marine radars.
Methods
For this study, we used data from two different primary sources. Insect data were from measurements made on specimens from two entomological collections in Mexico, whereas bird data were from flight 1.22 for Windows (companion material for Pennycuick 2008, latest version available at http://www.elsevierdirect.com/companion.jsp?ISBN=9780123742995). We used this program to estimate theoretical airspeeds of both insects and birds.
Study Area
Continuing radar observations of nocturnal bird migration over the Isthmus of Tehuantepec, Oaxaca, southern Mexico (Fig. 1), suggest the need for an appropriate criterion to filter birds from insects. The Isthmus of Tehuantepec is a narrow region that separates the Gulf of Mexico from the Pacific Ocean; it is located at the overlap of the Neotropic and Neartic realms and its level of plant endemism is high (Pérez-García et al. 2001). The Isthmus of Tehuantepec is an important corridor for migratory birds moving between North and South America (Binford 1989) and an important stopover site for migratory birds in the fall (Winker 1995a).
Study site and radius used to delimit the distribution of insects used in this study in Oaxaca; square shows the Isthmus of Tehuantepec.

Recording of Bird Data
For airspeeds to be estimated by the aerodynamic approach, data on a bird's wing span, wing area, and weight are needed. Although we know the species of the Isthmus of Tehuantepec from our program monitoring bird migration and from the literature (Binford 1989, Winker 1995a, b), we have not recorded values of the variables mentioned above. Therefore, we used data preloaded in the software FLIGHT 1.22 (Pennycuick 2008) from American or European passerine species (or families) with physical dimensions similar to those present in the isthmus (Fig. 2).
Variables used by FLIGHT 1.22 to estimate flight speeds

Variables used by FLIGHT 1.22 to estimate flight speeds
Recording of Insect Data
We obtained information on regional insect diversity from the Sistema Nacional de Información sobre Biodiversidad (SNIB), operated by the Comisión Nacional para el Conocimiento y Uso de la Biodiversidad (CONABIO) in Mexico. CONABIO provided a list of more than 21 000 records of 1302 insect species from the most comprehensive database of biological diversity in Mexico.
Of the insect records obtained from the SNIB, we used only those of species present within a 65-km radius around La Venta, Oaxaca, covering the surroundings of the study site and out to the borders of the state of Veracruz (Fig. 1). We filtered the data further by including only insect families that might be detected by the radar: the Scarabaeidae, Melolonthidae, Cerambicidae, and Passalidae (Coleoptera) and Uranidae, Saturniidae, Sphingidae, Noctuiidae, and Arctiidae (Lepidoptera.We then visited the following entomological collections: IEXA at the Instituto de Ecología, A.C. (INECOL), Xalapa, and the Colección Nacional de Insectos (CNIN) from the Universidad Nacional Autónoma de México (UNAM) to get images of selected species.
Using a Sony Cybershot digital camera, we photographed insect specimens mounted on a Styrofoam sheet with paper gridded in millimeters as a background for size reference. Then we measured each specimen's wingspan and length with the aid of IMAGEJ (Rasband 1997–2009) and estimated its weight by body relationships (Rogers et al. 1976, Sample et al. 1993, Miller 1997).
Airspeed Estimations
We estimated Vmp and Vmr of insects and birds with the software flight 1.22 for Windows (Pennycuick 2008), which requires data from eight variables (Table 1). Three of these variables are directly related to insectan and avian morphology (all up mass, wingspan, wing area; see Table 1 for definitions), others are environmental (gravity, air density) and the rest (frontal area of body, drag coefficient of body, induced power factor) are aerodynamic constants defined by Pennycuick (1997) as suitable for flight-performance estimations.
Statistical Analyses
We loaded flight 1.22 (Pennycuick 2008) with our insect measurements and estimated insects' theoretical airspeeds. For birds we used the preloaded data for passerines to estimate their theoretical airspeeds. For both groups we calculated airspeeds under the assumption of continuous flapping flight.
We assessed the normality of both insectan and avian Vmp and Vmr by a Shapiro—Wilk W-test; after non-normality was evident, we compared them with a Mann—Whitney U-test, then obtained a third set of airspeeds ( ) by calculating the mean of each estimated value of Vmp and Vmr. We plotted the proportion of species classified incorrectly for each of the three flight speeds (Vmp, Vmr, and ) as a function of the airspeed and defined an interval of potential airspeeds that can be used as a criterion to distinguish between insectlike and birdlike targets.
) by calculating the mean of each estimated value of Vmp and Vmr. We plotted the proportion of species classified incorrectly for each of the three flight speeds (Vmp, Vmr, and ) as a function of the airspeed and defined an interval of potential airspeeds that can be used as a criterion to distinguish between insectlike and birdlike targets.
Results
We calculated minimum power speeds and maximum range speeds (Vmp and Vmr, respectively) for 60 individual insects from 27 species of Lepidoptera and 9 of Coleoptera collected in the Isthmus of Tehuantepec (Table 2) and for 60 species of passerines, 50 belonging to taxonomic families present in the isthmus (Table 3). Insects range from one of the small tiger moths (Hypercompe extrema) of the family Arctiidae (b = 4.5 cm and m = 53.6 mg) to an unidentified large sphingid moth (b = 15.8 cm, m = 3.7 g). For birds, size ranged from warblers like Northern Parula (Setophaga americana, b = 18 cm, m = 8.9 g) to the Carrion Crow (Corvus corone, b = 92.5 cm, m = 553 g), one of the largest passerines. Overall, avian Vmp and Vmr are higher than insects' (Fig. 3).
The minimum power speeds (Vmp) of both insects (W = 0.91, P < 0.001) and birds (W = 0.91, P < 0.001) were not normally distributed, and birds' airspeeds were higher than those of insects (U = 3, Z = 9.43, P < 0.001). They overlapped at 5.5 ± 0.41 m sec-1 (Fig. 3a), a value similar to that for temperate areas. Similarly, insectan (W = 0.91, P < 0.001) and avian (W = 0.90, P < 0.001) maximum range speeds (Vmr) were not normally distributed either, showed the same pattern as Vmp (U = 1, Z = 9.44, P < 0.001), and overlapped at 12.2 ± 0.57 m sec-1 (Fig. 3b). Insectan and avian overlapped at 9.0 ± 0.49 m sec-1 (Fig. 3c). From Figure 3, it is clear that some insects flying at Vmr are faster than many birds flying at Vmp , a fact that helps to explain the two groups' overlap in flight speed reported in literature.
On the basis of our results and reports of insectan and avian airspeeds from the literature, we consider that an airspeed criterion for any region with insects and birds with physical dimensions similar to those considered in this study lies between the insectan and avian overlap at Vmp (5.5 m sec-1) and the overlap at (9 m sec-1). Given the proportion of species classified both correctly and incorrectly with each of the theoretical airspeeds, we consider that an airspeed of ∼7 m sec-1 (the intersection between insectan and avian Vmp) is
well suited for our study area because it classifies correctly all the insects flying at Vmp, most of the insects flying at , all the birds at and Vmr, and most birds flying at Vmp (Fig. 4).
Discussion
One of the biggest challenges with the use of marine radars in the study of bird migration is distinguishing targets that may be insects, birds, or bats. This problem has been approached through the use of multiple criteria that allow them to be separated to some degree. Airspeed is one of the main criteria because birds, in general, fly faster than insects (Ellington 1991). It is, however, known, that flight speeds of insects and birds can overlap (Larkin 1991), and the degree of the overlap depends on the insect species aloft. For this reason we needed information on the local insect fauna (Zaugg et al. 2008) in order to estimate the overlapping airspeed of insects and birds at our study site.
Although direct measurement would be the ideal way to get insectan and avian airspeeds, it is not logistically feasible in most field studies, so we used a theoretical approach as an alternative to estimate airspeeds of local species of insects and birds. This theoretical approach was originally developed for the study of birds' flight performance (Alerstam and Hedenström 1998), but it is applicable to any flying animal (Pennycuick 1972, 2008, Tennekes 2009) because it is based on aerodynamic principles and physical dimensions of animals. The key input for this approach is the physical dimensions of the animals of interest. We used the data for passerines preloaded in FLIGHT 1.22 to estimate airspeeds because we believe the similarity in physical dimensions between species migrating over the Isthmus of Tehuantepec and those included in the software justifies this approach. Insect data, however, were not included in the software so had to be collected from the area of interest.

Insectlike (black bars) and birdlike (gray bars) theoretical speeds calculated with FLIGHT 1.22 (Pennycuick 2008). (a) Minimum power speeds (Vmp), (b) maximum range speeds (Vmr), (c) mean flight speeds (). Values were rounded to the closest 0.5 m sec-1.
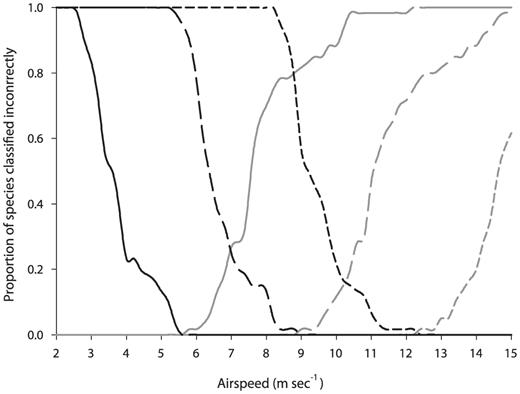
Proportion of species classified incorrectly for each of the three flight speeds as a function of airspeed. Black lines, insects;
gray lines, birds. Vmp, solid lines; , lone-dashed lines; Vmr, Shortdashed lines.
We initially believed that the criterion of 6 m sec-1 used in some temperate regions (Larkin 1991) would not be applicable for radar studies in our tropical region because of differences in the size of insects. For example, Larkin (1991) reported one species of moth, the Cabbage Looper (Trichoplusia ni, wingspan ∼35 mm, Arnett 2000); although we acknowledge that other (probably larger) species might also have been included by Larkin (1991), our study area contained much larger moths, like the Owl Moth (Thysania zenobia) with a wingspan of 137 mm. The wingspan of the insect species we considered ranged from 45 to 158 mm.
From our results it is clear that, as reported in the literature, there is overlap between the theoretical speeds of insects and birds. As a consequence, there is no single best airspeed criterion; instead, one must choose within a range or interval of airspeeds on the basis of characteristics (such as physical dimensions of insects and birds) particular to an area. The choice entails a tradeoff in the proportion of targets correctly classified as birds or insects.
We estimated three airspeeds for every species of bird and specimen of insect analyzed: Vmp, Vmr and . Vmp and Vmr encompass the range of potential flight speeds that the species under consideration (or others of similar dimensions) can reach. The third estimate () is the mean of Vmp and Vmr. Although Vmp and Vmr of birds and insects overlap in an easily
distinguishable way, we considered that those overlap values are not suitable for use as a criterion to separate birds from insects because those results suggest a very low airspeed for Vmp and a very high value for Vmr. Hence we felt an intermediate value such as was warranted.
As insectan (mainly lepidopteran) airspeeds measured in the tropics (Srygley and Dudley 1993,2008, Dudley and Srygley 1994,2008, Dudley et al. 2002) are more similar to our Vmp than to Vmr, and as Bloch and Bruderer (1982) observed that most of migrating birds (mostly passerines and hence with physical dimensions similar to those of the species we considered) fly in the range of 8–18 m sec-1, we propose the overlap of birds and insects at Vmp (5.5 m sec-1) and the overlap at (9 m sec-1) define an appropriate range of airspeeds within which a criterion may be set. In our study area, we believe that an airspeed of ∼7 m sec-1 is an appropriate criterion because it minimizes the inclusion of slow-flying insects and maximizes the inclusion of most birds. By calculating an interval and allowing a suitable airspeed to be chosen as a criterion, investigators can incorporate their understanding and knowledge of flight speeds, composition of local insectan and avian fauna, and their study objectives. We encourage the reader to consider these factors and to choose an airspeed value that best fits his study's objective.
A criterion based on airspeeds will not only minimize confounding factors from insects, it will also help eliminate some bats, as there are records of some bats flying slower than the proposed interval of airspeeds (Hayward and Davis 1964, Vaughan 1966, Patterson and Hardin 1969, Kennedy et al. 1977, Morrison 1980, Sahley et al. 1993, Hopkins et al. 2003). However, although airspeed is a criterion useful for discriminating insectlike targets from birds, we acknowledge that this value will not provide complete certainty for discriminating among all vertebrate targets because bats fly at a wide range of speeds (Winter 1999), so there will always be some flight-speed overlap among birds, insects and bats.
To reduce contamination of the dataset and increase certainty that the analyses actually address the group of interest, however, we strongly recommend the use of additional criteria, such as omitting targets with poor reflectivity and small targets that appear only within ∼500 m of the radar and using behavioral traits to predict a species' timing and pattern of movement. Current research on wing-beat patterns may also prove useful in distinguishing radar targets. Techniques that corroborate the identification of radar targets (e.g., night-vision optics, thermal imagery) are useful and should be used in conjunction with marine radar studies if and when possible.
Acknowledgments
We thank CONABIO for providing information on insect diversity in the state of Oaxaca, to the Colección Nacional de Insectos (CNIN) at the Universidad Nacional Autónoma de México (UNAM), and to the Colección Entomológica (IEXA) at the Instituto de Ecología, A.C. (INECOL), for granting us access to their specimens. Particular thanks to Roberto Arce for his invaluable support in insect identification and beetle handling at IEXA and to Alex Prichard for his suggestions on graphical analyses. Sergio A. Cabrera-Cruz was supported with a scholarship from the Consejo Nacional de Ciencia y Tecnología (CONACYT) during this research. Two anonymous reviewers provided valuable comments that greatly improved the manuscript.
Literature Cited

{kind=link}
{kind=link}
{kind=link}
{kind=link}