





Abstract.
A combination of elements (e.g., crops and pasture lands, strips of roadside vegetation, woodlots, temporary wetlands) increases the heterogeneity of rural landscapes. Agricultural landscapes range from homogeneous, dominated by a single element (i.e., pure cropland or pure pastoral farming) to heterogeneous, dominated by two or more elements (i.e., mixed farming). In this study we characterized landbird assemblages in various agricultural landscapes (cropland, pastoral, and mixed landscapes) in the Pampas of central Argentina, surveying along transects to quantify species richness, composition, and abundance. Mixed landscapes were more heterogeneous than pastoral areas and cropland. Species richness, particularly of generalists, was greater in mixed landscapes. Richness of grassland specialists (species of increased conservation concern in the Pampas) was lower in croplands than in pastoral and mixed landscapes. An indicator-species analysis supported the association of some grassland specialists (Rhea americana, Asthenes hudsoni, Embernagra platensis, Pseudoleistes virescens) with pastoral landscapes. Our results suggest that maintaining landscape heterogeneity throughout the Pampas (i.e., mixed landscapes) is important for preserving species richness of birds. But several threatened grassland specialists inhabiting the Pampas, such as Rhea americana and Asthenes hudsoni, are found exclusively in pastoral landscapes, so the long-term survival of these species will depend on the conservation of this landscape type.
Resumen.
La combinación de elementos del paisaje (e.g., cultivos, pastizales, vegetación de bordes de caminos, arboledas y humedales temporarios) incrementa la heterogeneidad y define diferentes tipos de paisajes rurales, desde paisajes homogéneos dominados por un único elemento (i.e., paisaje de cultivo o ganadero) hasta paisajes heterogéneos dominados por dos o más elementos (i.e., paisajes mixtos). En este estudio caracterizamos los ensambles de aves terrestres en diferentes tipos de paisajes rurales de la Pampa argentina (paisajes de cultivos, ganaderos y mixtos). Las aves terrestres fueron muestreadas con transectas para determinar la riqueza de especies, la composición y la abundancia en cada tipo de paisaje rural. El paisaje mixto tuvo una mayor heterogeneidad que los paisajes de cultivos y ganaderos. El paisaje mixto tuvo una mayor riqueza de especies que los paisajes de cultivos y ganaderos, en especial de especies generalistas. Por otro lado, el paisaje de cultivos tuvo la menor riqueza de especialistas de pastizal (aquellas con problemas de conservación en la Pampa) que los paisajes ganaderos y mixtos. Un análisis de especies indicadoras destacó la asociación entre algunas especies de pastizal (Rhea americana, Asthenes hudsoni, Embernagra platensis, Pseudoleistes virescens) y el paisaje ganadero. Nuestros resultados sugieren la importancia de mantener la heterogeneidad del paisaje en los agroecosistemas de la Pampa argentina (i.e., paisajes mixtos) para preserver la riqueza de aves. Sin embargo, cabe señalar que varias especies amenazadas de pastizal que habitan en los pastizales de las pampas, como R. americana y A. hudsoni, se encuentran exclusivamente en paisajes ganaderos, por lo que su supervivencia a largo plazo dependerá de la conservación de este tipo de paisaje.
Introduction
In agricultural areas, various features of the landscape (e.g., crops and pasture lands, strips of roadside vegetation, woodlots, temporary wetlands) provide important resources for birds and other wildlife that require herbaceous plants, bushes, and trees for feeding, nesting, and shelter (Farina 1997, Mazerolle and Villard 1999, Hinsley and Bellamy 2000, Fuller et al. 2004). The combination of such elements increases the landscape's heterogeneity (Haslem and Bennett 2008) and defines various types of agricultural landscapes, from homogeneous, dominated by a single element (i.e., pure cropland or pure pasture lands) to heterogeneous, dominated by two or more elements (i.e., mixed landscapes). Within a specific agricultural landscape, the richness and abundance of birds may be increased by particular components of cropland, pastures, or mixed farming (Murphy 2003, Askins et al. 2007, Herzon and O'Hara 2007). For instance, in heterogeneous landscapes, mixed farming may play an important role in birds' population dynamics, since many generalist species may use different habitat types for early and late nesting (Atkinson et al. 2002).
Intensively exploited agricultural landscapes support fewer species and fewer individuals of specialist species than do landscapes under less intensive use (Herzon and O'Hara 2007). For instance, in homogeneous landscapes, livestock grazing is expected to have a less negative effect on most bird populations because it is less intensive and intrusive (i.e., no chemical applications or mechanical treatments) than crop production (Chamberlain et al. 2000, Peterjohn 2003, Askins et al. 2007, Norris 2008).
The associations among landbird species in different agricultural landscapes may vary considerably depending on their life-history attributes (Hansen and Urban 1992) such as residence status (Murphy 2003), feeding habits (Robinson et al. 2001), or habitat specialization (Atauri and de Lucio 2001, Söderström et al. 2003). More interestingly, Murphy (2003) pointed out a correlation between birds' life-history traits and use of specific landscape features in the eastern and central United States, showing that richness and abundance of grassland birds were negatively affected by an increase in agricultural land use.
The Pampas of Argentina were formerly an extensive grassland that has become one of the largest agricultural regions of the world (Soriano et al. 1991). As elsewhere in South America (Morton et al. 2006), the establishment of agroecosystems in the Pampas has substantially modified the original vegetation (Baldi et al. 2006, Bilenca et al. 2009) as well as the composition and abundance of native bird species (Wilson 1926, Daguerre 1936, Narosky and Di Giacomo 1993). During recent decades, moreover, there has been a deeper change in the pattern of land use, more intense agricultural practices being substituted for rotation between crops and pastures (Viglizzo et al. 2011). This change was based principally on the introduction of no-tillage cultivation and an herbicide-tolerant genetically modified soybean, which has promoted the conversion of pasture lands into cropland and led to the decline in number of livestock and extent of pasture lands (Baldi and Paruelo 2008, Viglizzo et al. 2011). Although the structure of modern farmlands has been generally simplified, disparate levels of spatial homogenization were superimposed on the original landscape (Baldi et al. 2006).
Recent studies of the avifauna of central Argentina have shown that the population density of several species of birds decreased as the proportion of cropland in the landscape increased (Filloy and Bellocq 2007, Codesido et al. 2008, Cerezo et al. 2011). Codesido et al. (2011) detected a reduction in distribution of grassland birds in association with land-use changes, in particular with the loss of grasslands and increase of crops. The association between attributes of bird species and various agricultural landscapes has been well documented in other temperate-zone agroecosystems (Atkinson et al. 2002, Murphy 2003, Askins et al. 2007, Herzon and O'Hara 2007). In the Pampas, some studies have attempted to quantify similar associations but covered restricted geographic scales (Cerezo et al. 2011) or considering specific groups (e.g., diurnal raptores, Pedrana et al. 2008) only. However, no studies in the Pampas evaluated the associations between bird assemblages and various agricultural landscapes.
We hypothesized that species richness and abundance of landbirds should be higher in heterogeneous landscapes than in homogeneous landscapes (Benton et al. 2003). Therefore, according to this hypothesis, we predict that species richness and abundance of landbirds should be higher in mixed landscapes than in landscapes of either cropland or pastures alone. As a complement, we also hypothesized that species richness and abundance of specialist birds should be higher in agricultural landscapes under less intensive use (Chamberlain et al. 2000). Therefore, according to this hypothesis, we predict that species richness and abundance of specialist birds should be higher in pastoral landscapes than in cropland. Thus the objectives of our study are (1) to compare species richness and abundance of generalists, wetland specialists, and grassland specialists (the species of greatest conservation concern in the rural Pampas) in various agricultural landscapes and (2) to identify the indicator species of birds in each category of landscapes.
Methods
Study Area
Our study area in the Pampas region of central Argentina extends over about 225 000 km2 (500 km from north to south, 450 km from east to west; 33–39 °S, 57–63° W; Fig. 1). In this region, July mean temperature ranges from 7.5 to 9.5 °C, January mean temperature from 21.5 to 23.5 °C, and annual precipitation from 800 to 1000 mm. Previously, the main vegetation types were prairies and steppes, dominated by grasses (e.g., Stipa, Piptochaetium, Aristida, Melica, Briza, Bromus, Eragrostis, and Poa), interspersed with marshes and other communities as governed by local edaphic conditions, but the area has been heavily modified by long-term human use (Soriano et al. 1991, Burkart et al. 2005). Rotation between crops and pastures has been now replaced by rotation between crops (wheat and soybean, primarily) or by a sustained production of one crop (Aizen et al. 2009). Originally, trees were absent, entering the region only marginally near the ecotones with the neighbouring Delta and Espinal ecoregions (Burkart et al. 1999; Fig. 1). In the Pampas, stands of tall trees are novel, and their introduction has added structural complexity to the rather two-dimensional grasslands of this region. Nowadays, woodlands of both native and exotic species have been established in some places of the region as riparian vegetation or along roadsides (Ghersa et al. 2002).
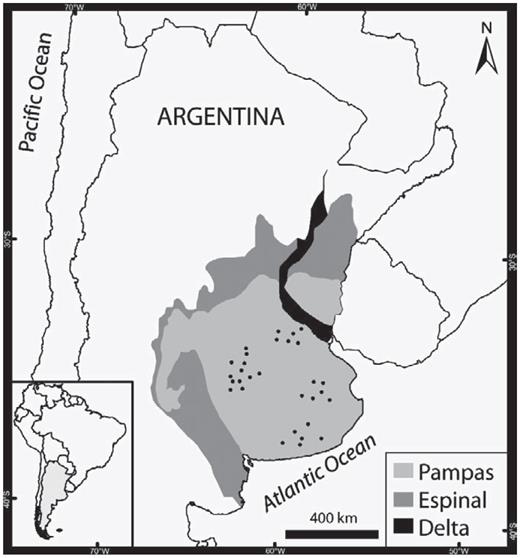
Location of the 30 survey transects in the Pampas of central Argentina.
Land-Use Intensity and Agricultural Landscapes
We surveyed along 30 transects located on secondary and tertiary roads in agroecosystems of the Pampas (Fig. 1). Each transect was 20 km long with permanently marked survey points located every kilometer. At each point (within a 200-m radius), we recorded the cover of various land uses, considering the following broad categories common in the study region and characteristic of many agricultural landscapes around the world (Fuller et al. 1997, Haslem and Bennett 2008, Walk et al. 2010): grasslands (including pastures, fallow fields, and fields of natural vegetation), croplands (including cultivated, stubble, plowed, and/or harrowed fields), woodlots (either natural or plantations of trees >0.2 ha), roadside vegetation, and wetlands (including areas with floating or rooted vegetation on the banks of streams, ponds, or along roadsides). For each transect, we calculated the percentage of each land-use type as the summed cover by land-use type at 20 points along each of the 30 transects.
In the Pampas, crops and pastures are the dominant covers (the sum of both coverages is always >80% of area). Therefore, we defined three agricultural landscapes based on coverage of pastures and crops as follows. (1) Cropland landscapes (homogeneous): transects along which crop fields cover >50% of the area (Fig. 2a), (2) pastoral landscapes (homogeneous): transects along which grassland fields cover >50% of the area (Fig. 2b), and (3) mixed landscapes (heterogeneous): transects that did not meet either of the criteria above (Fig. 2c). As a result of applying this classification, we obtained ten transects for each of the three categories.
Landbird Sampling
Following the point-count method (Bibby et al. 2000), Codesido surveyed along each transect twice in summer (January or February), 2006–2008. Counts took place either during the 3 hr after dawn or the 3 hr before sunset. Each transect consisted of 20 points. For 5 min, all landbirds seen or heard within a 200-m radius around each point were identified and counted, resulting in a total sampling effort of 100 hr. We did not count birds on windy or rainy days. We classified all species by their nesting habitat as grassland specialists, wetland specialists, or generalists (see Codesido et al. 2011, 2012). We excluded from our analyses species that do not breed in the Pampas, either nearctic migrants such as the Swainson's Hawk (Buteo swainsoni), Upland Sandpiper (Bartramia longicauda), and Cliff Swallow (Petrochelidon pyrrhonota) or austral migrants from Patagonia such as the Austral Negrito (Lessonia rufa; Narosky and Di Giacomo 1993).
Data Analysis
To examine the spatial independence of species richness on the transects, we ran a Mantel test for spatial autocorrelation (Legendre and Legendre 1998). We correlated two dissimilarity matrices based on species richness and geographical position per transect. We also tested for spatial autocorrelation for bird abundance by correlating dissimilarity matrices of bird abundance and geographical position. We constructed two sets of matrices, one for each variable, based on the properties of the Euclidean distance index. This tests whether species richness and bird abundance along transects in close proximity are more similar than those that are spatially distant. We calculated probability values by comparing the observed dissimilarity of these matrices to that of 1000 random permutations of both matrices.
We calculated the heterogeneity for each transect in each category of landscape by the complement of Simpson's index (Magurran 1988),  , where pi is the proportion of each cover type in the transect. We controlled the effect of rare species (i.e., rare species may unduly influence the results) by considering only those species recorded on more than 10% of transects (Söderström et al. 2003). For species richness we used the average of the two surveys of each transect. Similarly, we calculated the abundance of each species in each category of landscape. We compared the richness and abundance of landbirds by category with two separate one-way ANOVA. Values were square-root transformed when necessary to satisfy assumptions of normality and homogeneity of variance (Zar 1999). Then we used the Tukey test in order to determine whether means in each category of landscape differed significantly (Zar 1999).
, where pi is the proportion of each cover type in the transect. We controlled the effect of rare species (i.e., rare species may unduly influence the results) by considering only those species recorded on more than 10% of transects (Söderström et al. 2003). For species richness we used the average of the two surveys of each transect. Similarly, we calculated the abundance of each species in each category of landscape. We compared the richness and abundance of landbirds by category with two separate one-way ANOVA. Values were square-root transformed when necessary to satisfy assumptions of normality and homogeneity of variance (Zar 1999). Then we used the Tukey test in order to determine whether means in each category of landscape differed significantly (Zar 1999).

Typical count points representative of the landscape types in the Pampas of central Argentina. (a) Field of soybeans (Glycine max) in the cropland landscape. In the center, note the lack of roadside vegetation between crop and road, (b) Grassland in a pastoral landscape. Cattle grazing may create grasslands with greater horizontal and vertical structural diversity. Note the presence of roadside vegetation along the fence rows, (c) Field of corn (Zea mays) in a mixed landscape. Note the presence of roadside vegetation along the fence rows.
We used species' presence/absence along each transect to calculate the Bray—Curtis similarity index among categories of landscape, then compared bird assemblages by category with an analysis of similarity (ANOSIM; Clarke 1993). ANOSIM compares the level of similarity among transects of a given category of landscape to that among transects of all categories and determines if the former is greater than expected by chance. ANOSIM also estimates pairwise comparisons, so it can be used to determine species turnover by category. We tested results from ANOSIM for significance with a randomization procedure. Then we used nonmetric multidimensional scaling to compare species composition by transect and category of landscape. This analysis portrays similarities and differences among samples graphically so that points that are closer are more similar in terms of species composition. For Anosim and multidimensional scaling we used PAST (Hammer et al. 2001). To identify species characteristic of each landscape, we used an indicator-species analysis (Dufrêne and Legendre 1997). This analysis calculates an indicator value for each species on the basis of its relative frequency and relative abundance in each category. Indicator values can range from 0 (no indication) to 100 (perfect indication). A perfect indication for a given landscape means that the species was recorded on all transects within that landscape and was not observed in any other landscape. We tested indicator values for significance with a Monte Carlo randomization procedure, which compares the observed indicator values to alternative values calculated from the same data and randomly assigned to landscape type. We report only species whose indicator values are significant (P ≤ 0.01) and ≥40% (Azpiroz and Blake 2009). For indicator-species analysis we used PC-ORD version 4 (McCune and Mefford 1999).
Results
Agricultural Landscapes and Heterogeneity
Mantel tests confirmed no spatial autocorrelation between transects in either species richness or bird abundance (P > 0.05 in all cases; transects were spatially independent). In cropland landscapes, crop cover was 67% on average, whereas in pastoral landscapes mean grassland cover was 62% (Fig. 3). The categories of landscapes also differed in that mixed and pastoral landscapes had greater cover of roadside vegetation than did cropland landscapes (ANOVA, F2,27= 4.9, P < 0.05; Fig. 3), whereas pastoral landscapes had greater cover of wetlands than did the other two landscape types (ANOVA, F2,27= 8.7, P < 0.01; Fig. 3). As expected, the heterogeneity of mixed landscapes was greater (S = 2.92 ± 0.13) than that of pastoral and cropland landscapes (S = 2.24 ± 0.12 and S = 1.98 ± 0.09, respectively; ANOVA,F2,27 = 18.8,P <0.0001).
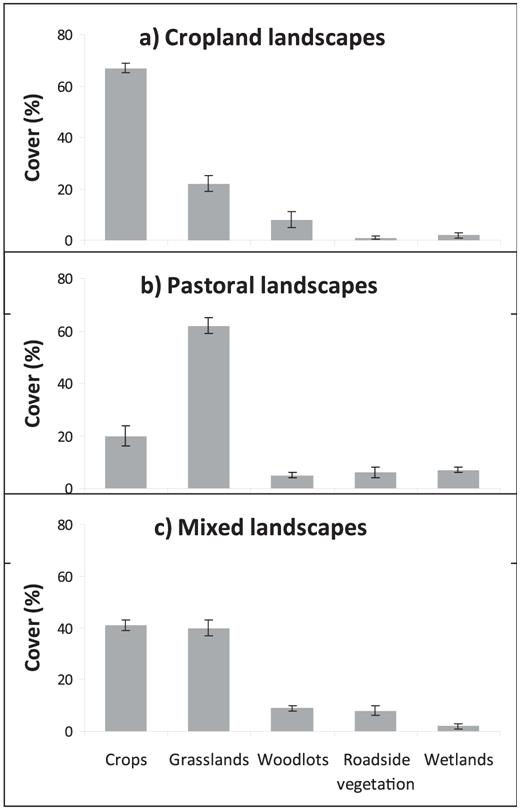
Mean cover of habitats (± SE) in (a) cropland landscape, (b) pastoral landscape, and (c) mixed landscape in the Pampas of central Argentina.
Bird Assemblages
Rare species excluded, we recorded 67 species of breeding landbirds, of which 49 were generalists, 13 grassland, and 5 wetland specialists (Appendix, available at http://dx.doi.org/10.1525/cond.2012.120011). Patterns of species richness differed by landscape type. Species richness was greater in mixed landscapes than in pastoral and cropland landscapes (ANOVA, F2,27= 8.3, P = 0.0016; Fig. 4a). Species richness of generalists was greater in mixed landscapes than in pastoral landscapes (ANOVA, F2,27 = 4.9, P = 0.015; Fig. 4a). Richness of grassland specialists was lower in cropland landscapes than in pastoral or mixed landscapes (ANOVA, F2,27= 7.9, P = 0.002; Fig. 4a).
The abundance of birds was greatest in pastoral landscapes, lowest in cropland landscapes, and intermediate in mixed landscapes (ANOVA, F2,27 = 4.9, P = 0.015; Fig. 4b). The abundance of grassland specialists was also lower in cropland landscapes than in pastoral and mixed landscapes (ANOVA, F2,27= 5.1, P = 0.014; Fig. 4b). Pastoral landscapes had an abundance of wetland specialists greater than that of cropland and mixed landscapes (ANOVA, F2,27 = 4.03, P = 0.03; Fig. 4b).
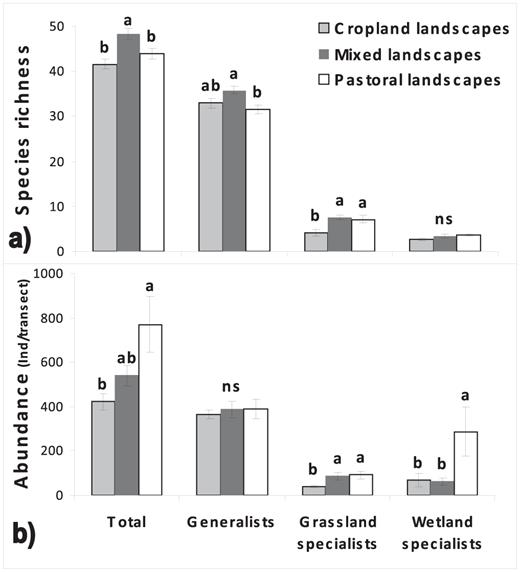
Mean (± SE) of (a) species richness and (b) abundance of generalists, grassland specialists, wetland specialists, and all species pooled in cropland, mixed, and pastoral landscapes in the Pampas of central Argentina. Different letters indicate significant differences (Tukey test; P < 0.05).
Anosim results confirmed that the bird communities of these categories of landscape differed significantly (global R = 0.17, P = 0.003). In terms of spatial turnover by pairwise comparisons, the only significant differences in terms of species composition were between cropland and pastoral landscapes (R = 0.26, P = 0.009), whereas the species composition of mixed landscapes did not differ significantly from that of either of the other two landscape types (P > 0.05). The ordination by multidimensional scaling grouped transects by landscape type along the second ordination axis with pastoral and cropland landscapes occupying extremes (Fig. 5)
Indicator-species analysis identified 11 species with significant indicator values; one was associated with cropland landscapes, four with mixed landscapes, and six with pastoral landscapes (Table 1). Four out of the six indicator species for pastoral landscapes, the Great Pampa-Finch (Embernagra platensis), Brown-and-Yellow Marshbird (Pseudoleistes virescens), Greater Rhea (Rhea americana), and Hudson's Canastero (Asthenes hudsoni), are grassland specialists, whereas indicator species for the other landscape types are all generalists (Table 1).
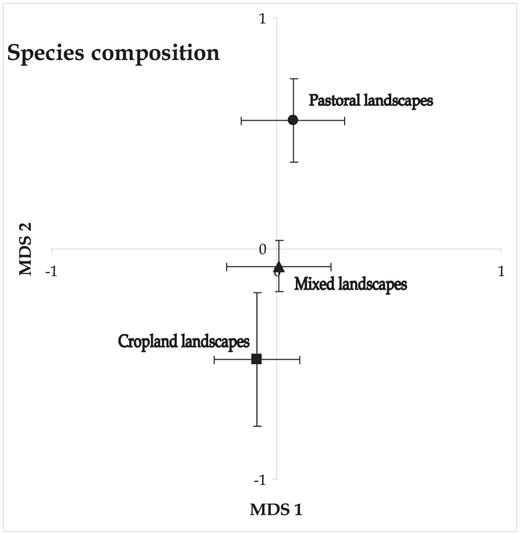
Nonmetric multidimensional scaling based on species' presence/absence per 30 sample transects within three categories of agricultural landscape. Symbols represent the mean and standard error of transects in each landscape type, to enhance visual interpretation of the ordination.
Discussion
Landscape Heterogeneity
Overall, our results suggest that landscape heterogeneity affects landbird richness in the agroecosystems of the Pampas of central Argentina. The positive influence of landscape heterogeneity on faunal diversity in agricultural landscapes has been recognized (Atauri and de Lucio 2001, Herzon and O'Hara 2007, Haslem and Bennett 2008) and in Europe is considered a key factor influencing the conservation of farmland birds (Benton et al. 2003). In the Pampas, mixed landscapes were more heterogeneous and supported greater bird richness. Mixed landscapes had both generalist and specialist species because of greater landscape heterogeneity that provides different niches and multiple ways to exploit environmental resources. Mixed farming increased the abundance of generalists, that is, indicator species such as the American Kestrel (Falco sparverius), Rufous-collared Sparrow (Zonotrichia capensis), Shiny Cowbird (Molothrus bonariensis), and Bay-winged Cowbird (Agelaiodes badius), and these positive responses should be associated with more diverse components of the habitat because of increased opportunities for foraging and nesting (Atkinson et al. 2002). Additionally, in mixed landscapes, some species of grassland specialists use patches of grassland for foraging or nesting, as pointed out by other studies (Bucher and Nores 1988, Best et al. 1995, McMaster and Davis 2001). However, in this regard it should be pointed out that mixed landscapes are not able to keep all grassland specialists, as some grassland specialists such as the Greater Rhea and Hudson's Canastero are sensitive to landscape context and are exclusively dependent on pastoral landscapes (Table 1).
Indicator values (as percentage of perfect indication) and values of Monte Carlo test of significance of observed maximum indicator values. Only values significant at least at P ≤ 0.01 and ≥40% are shown; gen, generalist; gra, grassland specialist.
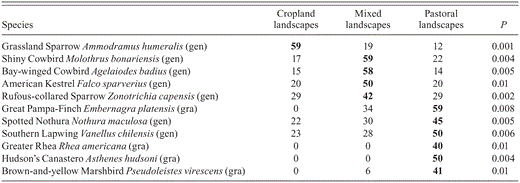
Indicator values (as percentage of perfect indication) and values of Monte Carlo test of significance of observed maximum indicator values. Only values significant at least at P ≤ 0.01 and ≥40% are shown; gen, generalist; gra, grassland specialist.
Land-Use Intensification
The use of land for annual crops seems more detrimental to the avian community than is livestock grazing in the agroecosystems of the Pampas. According to our results, total abundance as well as abundance and richness of grassland species are lower in croplands than in pastoral landscapes. Grassland specialists being scarce in or absent from cropland suggest that this landscape type provides limited opportunities for grassland-nesting birds—not surprising since operations in agricultural fields, especially crop harvesting, can reduce nesting success (Best et al. 1995). During summer and autumn fields are covered with summer crops, mostly soybean, that have low vertical structure, and after harvest the land is used for winter crops, mostly wheat, resulting in exploitation year round (Viglizzo et al. 2011). Likewise, intensive agricultural practices have reduced roadside vegetation, resulting in a serious loss of avian biodiversity (Szpeiner et al 2007, Di Giacomo and Lopez de Casenave 2010, Codesido and Bilenca 2011).
Studies in Britain and northwestern Europe have pointed out that some generalists with greater habitat breadth are more tolerant of changing agricultural practicees (see Siriwardena et al. 1998, Robinson and Sutherland 2002). In some cases, this pattern is likely to be related to requirements for foraging; for instance, granivorous birds such as the Wood Pigeon (Columba palumbus; see Inglis et al. 1990) tend to be associated with annual crops, which habitat tends to be rich in seeds. In this regard, our results are in agreement with those of Filloy and Bellocq (2007), who also found that the abundance of the Grassland Sparrow (Ammodramus humeralis) is positively associated with the increase of croplands in the landscape of the Pampas. This granivorous species may fit successfully into high-intensity agriculture that provides abundant food such as cereal grains.
In the pastoral landscapes, grasslands shelter the greatest number of nesting specialists because these grasslands most closely resemble the pre-settlement conditions of the Pampas (Cerezo et al. 2011, Codesido et al. 2011). Land used for cattle grazing (at low to moderate rates) can provide a wide range of opportunities for birds that forage on invertebrates and seeds (Perkins et al. 2000), and pastures may offer the greater horizontal and vertical structural diversity that can support a greater abundance and diversity of grassland-breeding birds. Some of the indicator species of pastoral landscapes are generalists, such as the Southern Lapwing (Vanellus chilensis) and Spotted Nothura (Nothura maculosa), wich feed and nest on the ground in multiple types of open habitats (Bucher and Nores 1988, Stotz et al. 1996). Other indicator species are grassland specialists such as the Great Pampa-Finch, Brown-and-Yellow Marshbird and Hudson's Canastero, which place their nests in tall grass (Isacch and Martinez 2001, Isacch et al. 2003, Cozzani and Zalba 2009), and others such as the Greater Rhea, that nest in short grass surrounded by tall grass (de la Peña 2005). This also suggests that nest-site availability plays an important role in determining bird communities in this landscape type. This factor is thought to be key in shaping bird assemblages in agricultural landscapes in general (Söderström et al. 2003, Murphy 2003, Askins et al. 2007, Isacch and Cardoni 2011).
In the agroecosystems of the Pampas, the limited areas of woodlots, roadside vegetation, and wetlands are critical to many species of landbirds; in particular, the extent of roadside vegetation is a good predictor for both the whole group and the grassland specialists (Codesido and Bilenca 2011). Many species detected in our study use roadside vegetation either for breeding or foraging (Szpeiner et al. 2007, Di Giacomo and Lopez de Casenave 2010, Codesido and Bilenca 2011, Cerezo et al. 2011). Additionally, within pastoral landscapes temporary wetlands support greater abundances of wetland specialists such as the White-faced Ibis (Plegadis chihi) and Cattle Egret (Bubulcus ibis), which forage in the these zones as well as in grassland (see also Canevari et al. 1991, Codesido et al. 2012). In this regard, note that some grassland specialists, such as the Spectacled Tyrant (Hymenops perspicillatus), Warbling Doradito (Pseudocolopteryx flaviventris), Bay-capped WrenSpinetail (Spartonoica mahtroides), Sedge Wren (Cistothorus platensis), and Brown-and-Yellow Marshbird use also marsh vegetation associated with temporary wetlands (Isacch et al. 2004, Isacch and Cardoni 2011).
Conservation Implications
Overall, the effect of agricultural intensification and homogenization of the landscape toward cropland on the avifauna of the Pampas of central Argentina will depend on the sensitivity and plasticity of each species (Fernández et al. 2003, Filloy and Bellocq 2007, Pedrana et al. 2008). In this study, we found that the abundance and richness of most specialized grassland birds have declined over large areas of the Pampas that have been converted to intensive crop production. Our results suggest that maintaining heterogeneity throughout the Pampas (i.e., mixed landscapes) is important for preserving the species richness of birds. However, it should be pointed out that several threatened grassland specialists inhabiting the Pampas, such as the Greater Rhea and Hudson's Canastero (AOP/SADS 2008), are found exclusively in pastoral landscapes. These results highlight the importance of pastures for the conservation of native birds, especially when we consider that soybean cultivation is expanding, leading to greater homogeneity in the region's vegetation (Baldi and Paruelo 2008, Bilenca et al. 2008, Aizen et al. 2009, Viglizzo et al. 2011). In the same vein, a notable result of our study is that many species typical of grassland, such as the Long-winged Harrier (Circus buffoni) and Spectacled Tyrant, were absent or scarce along our transects, emphasizing the need to promote the conservation or restoration of native grasslands, in order to assure conservation of native avifauna. Failing that, the region's birds will consist only of generalists and an impoverished subset of grassland species.
Although large-scale changes in farming practices that increased the richness and abundance of grassland birds will be difficult to achieve within cropland landscapes, smallerscale measures with some elements of the countryside may increase habitat diversity significantly. For instance, maintaining networks of roadside vegetation (e.g., undisturbed roadside vegetation or fence rows) may be crucial for promoting habitat complexity and biodiversity in intensively managed agricultural landscapes (Szpeiner et al. 2007, Poggio et al. 2010, Codesido and Bilenca 2011, Cerezo et al. 2011).
Acknowledgments
We thank Lucas and Carlos Leveau for their assistance during field work. We appreciate the improvements in English usage made by Peter Lowther through the Association of Field Ornithologists' program of editorial assistance. This work was supported by the Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET—Argentina; PIP 2010-2012 GI11220090100231), Universidad de Buenos Aires (UBACyT; grants X282, X406, and 20020090100070), and Instituto Nacional de Tecnología Agropecuaria (INTA—Argentina; PNECO 1302).
Literature Cited

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}