





Abstract
With more than 50 described subspecies occurring throughout Southeast Asia and Micronesia and a wide array of morphological and ecological variation, the Island Thrush (Turdus poliocephalus) is one of the most variable known bird species. However, this bird has been studied surprisingly little from an evolutionary perspective, with no information on its evolutionary history to inform our understanding of its tremendous variation. In this study, we examine the population history of the Island Thrush within the Philippine Islands, sampling 88 individuals from six named Philippine subspecies and three populations currently lacking formal subspecific names, as well as several outgroups. Many subspecies are not reciprocally monophyletic, yet most island populations are, and show up to 2.6% sequence divergence (uncorrected). Philippine populations are not monophyletic; samples from Borneo, Sulawesi, Vanuatu, and the Solomon Islands are nested within the Philippines. Previously hypothesized routes of the colonization of the Philippines assume a single initial colonization event and cannot account for the current distribution of haplotypes, which implies at least two colonization events. Morphologically similar taxa in the Philippines are not each other's closest relatives, thus demonstrating plumage convergence. We emphasize the need for further study of this taxon throughout its range, particularly in light of conservation priorities.
Resumen
Con más de 50 subespecies descritas que se distribuyen en todo el sudeste de Asia y Micronesia, y con una amplia variación morfológica y ecológica, Turdus poliocephalus es una de las especies de ave con más variabilidad conocida. Sin embargo, esta especie ha sido estudiada poco desde una perspectiva evolutiva y no existe información sobre su historia evolutiva para entender su enorme variación. En este estudio, examinamos la historia poblacional de T. poliocephalus en las islas Filipinas, muestreando 88 individuos pertenecientes a seis subespecies filipinas descritas y a tres poblaciones que actualmente carecen de una descripción formal como subespecies, así como también de varios grupos externos. Muchas subespecies no son recíprocamente monofiléticas, aunque la mayoría de las poblaciones insulares lo son, con hasta un 2.6% de divergencia (no corregida) en las secuencias. Las poblaciones filipinas no son monofiléticas; muestras de Borneo, Sulawesi, Vanuatu y de las islas Solomon se encuentran anidadas dentro de las poblaciones filipinas. Las hipótesis existentes hasta ahora sobre las rutas de colonización de las Filipinas asumen un único evento de colonización inicial y no explican la distribución actual de haplotipos, la cual implica por lo menos dos eventos de colonización. Los taxones morfológicamente similares de las Filipinas no son los más emparentados entre sí, lo que demuestra existencia de convergencia de plumaje. Enfatizamos la necesidad de más estudios de este taxón a través de toda su área de distribución, particularmente debido a la prioridad que existe para su conservación.
Introduction
The Island Thrush (Turdus poliocephalus) is one of the most variable passerines in the world, with 52 (Clement 2000) or 53 (Mayr and Diamond 2001) described subspecies. The Song Sparrow (Melospiza melodia) and Blackbird (T. merula) are among the few species with comparable numbers of subspecies, but they display only a fraction of the plumage and distributional variation seen in the Island Thrush. As its name suggests, the species is restricted to islands, ranging from western Indonesia east to Samoa but is absent from Australia and New Zealand. Named subspecies are often single-island endemics, but larger, mountainous islands often contain multiple subspecies. These subspecies display tremendous plumage variation: head plumage may be black, white, or various shades of brown; the crissum may be plain or streaked; the belly can be uniform or may have up to three colors on it (Clement 2000). Populations of these subspecies exhibit classic examples of ecological character displacement, with the ranges of elevations that they inhabit on each island correlated to the number of Turdus species present on the island in suitable habitat. For example, Rennell Island (Solomon Islands) birds (T. p. rennellianus; Fig. 1) do not encounter other Turdus and are thus found down to sea level, whereas birds in New Guinea (T. p. carbonarius and others) share their habitat with more species and are restricted to highlands over 3000 m (Diamond 1975).
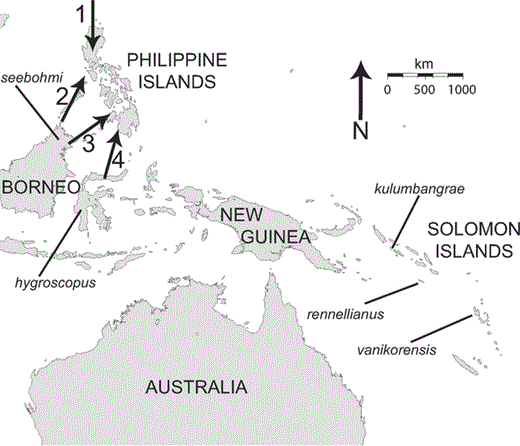
The landmasses of the southwestern Pacific. The five labeled subspecies of Island Thrush (Turdus poliocephalus) indicate the locations of individuals sampled to test the monophyly of Philippine populations (Fig. 2). Arrows labeled 1–4 represent colonization routes of Philippine organisms proposed by Dickerson (1928).

Map of the Philippine Islands, showing the names of major islands and the subspecies of Island Thrush that inhabit them. Dark gray areas represent modern landmasses in the Philippines, and light gray areas represent submerged land up to 100 m below sea level, approximating the distribution of land during the Pleistocene glaciation cycles. Subspecies names are taken from Dickinson et al. (1991); the Panay, Sibuyan, and Mt. Busa populations lack taxonomic names. Asterisks denote the approximate locations of origin of tissue samples. All individuals are represented by museum vouchers ( Appendix).
Throughout the range of the Island Thrush, the plumages of its populations are distinctive from subspecies to subspecies (typically from island to island), with adjacent populations dissimilar in plumage. For example, populations characterized by white-headed individuals are found only in Taiwan, Seram (eastern Indonesia), and in Vanuatu. Birds from one Luzon, Philippines population most closely resemble those in part of New Guinea. Within the Philippine archipelago, the populations show considerable plumage variation, and divergently plumaged populations separate similarly plumaged populations.
Conservation status varies geographically. Extinct birds were and very rare birds are generally restricted to smaller islands (such as Vanuatu, Lord Howe, Mare, and Norfolk Islands), yet other populations on small islands are common (including the Christmas Island population). Within the Philippines, no populations are considered rare (Kennedy et al. 2000), but the entire archipelago has experienced a tremendous rate of deforestation (Dickinson et al. 1991), and the loss of highland habitat on any of the six islands the birds inhabit could lead to endangerment or extinction of these populations or subspecies.
The Philippine islands represent a fringing archipelago—a series of islands adjacent to a continental shelf. Their component islands have never been directly attached to the mainland of Southeast Asia, even during sea level changes in the Pleistocene. Thus, overwater dispersal must account for the presence of the fauna here (Heaney 1986). Dickerson (1928) proposed four colonization routes for the Philippines based on knowledge of underwater shallow ridges (plate tectonics were not known at this time). The four routes, depicted in Figure 1, are: 1) north-to-south from Taiwan to Luzon, 2) south-to-north from Sulawesi to Mindanao, 3) from Borneo to Mindoro across Palawan, and 4) from Borneo to Mindanao across the Sulu Archipelago. A study of genus-level taxonomy by Diamond and Gilpin (1983) suggested the two routes from Borneo (i.e., routes 2 and 3) were of greatest importance. Dickinson et al. (1991) reviewed each of these four routes with a species-level distributional approach and found some evidence for each route, but did not specifically point to one route as being most important. Few published studies focus on intraspecific molecular phylogeography of any organism in the Philippines (Peterson and Heaney 1993, Miranda et al. 1998, Brown and Guttman 2002, Evans et al. 2003, Steppan et al. 2003, Gamauf et al. 2005, Heaney et al. 2005, Jansa et al. 2006), and of those that do, none presents results in a biogeographic context for any Philippine bird.
The Island Thrush is an ideal taxon for testing hypotheses of colonization patterns within the Philippines. It inhabits six Philippine islands, with seven named subspecies and three unnamed populations (Kennedy et al. 2000). The named subspecies (Fig. 2) are thomassoni and mayonensis in Luzon; mindorensis on Mindoro; nigrorum on Negros; and malindangensis, kelleri, and katanglad, on Mindanao. Unnamed populations are present on Sibuyan (Goodman et al. 1995), Panay (Kennedy et al. 2000), and on Mt. Busa in Mindanao (Kennedy et al. 2000). In this paper, we focus on the evolutionary history of Island Thrush populations within the Philippines, with sampling outside of this archipelago to infer colonization routes. Phylogenetic analyses are generated using mitochondrial DNA, the most appropriate type of molecular marker for discovering phylogeographic patterns at this taxonomic resolution (Zink and Barrowclough, in press).
Methods
Sampling
We obtained tissue samples for 84 individuals from throughout the Philippines from general collecting efforts of the National Museum of the Philippines (NMP), Cincinnati Museum of Natural History (CMNH), and the Field Museum of Natural History (FMNH). All birds are also represented by voucher specimens in these museums; both authors have examined all specimens from CMNH and NMP. Samples include representatives from six of the seven named subspecies in the Philippines (tissues for malindangensis from Mindanao are not available) as well as birds from three recently discovered populations (on the islands of Panay and Sibuyan and on Mt. Busa in Mindanao) currently lacking subspecific names; the sampling localities are designated in Figure 2.
A phylogenetic study of the genus Turdus found high support for monophyly of Island Thrush sampled from four islands (Voelker et al. 2007). We revisited the issue of monophyly by including that study's samples from the eastern portion of the Island Thrush range: T. p. rennellianus and T. p. kulumbangrae from the Solomon Islands and T. p. vanikorensis from Vanuatu (Genbank accession number DQ911111-DQ911113). We also sample three Island Thrush collected in Sulawesi (T. p. hygroscopus) by the American Museum of Natural History (AMNH) and one in Borneo (T. p. seebohmi) by the Louisiana State University Museum of Natural Sciences (LSUMZ). Detailed information on all samples is included the Appendix.
To orient the relationships of these sampled populations, two outgroups were selected based on the topology presented by Voelker et al. (2007). Their four Island Thrush samples are monophyletic with high support and are sister taxa to a clade of four species. We include two representatives of that clade in our analysis: the Eyebrowed Thrush (Turdusobscurus; AY049511) and Brown-headed Thrush (T. chrysolaus; DQ911070).
Dna Protocols
We isolated and purified DNA from tissues of specimens either via phenol-chloroform extractions (Hillis et al. 1996) or using a QIAamp Tissue Kit (Qiagen Inc., Valencia, California) following the manufacturer's protocols. The entire mitochondrial gene encoding the second subunit of NADH dehydrogenase (ND2) was then amplified from these extracts in 25 or 50 μl polymerase chain reactions (PCR). These reactions included extracted DNA and the primers L5215 (Hackett 1996) and H1064 (Drovetski et al. 2004). The PCR products were then cleaned with a QIAquick PCR Purification Kit (Qiagen Inc., Valencia, California) and then sequenced using the amplification primers as well as L347 (Drovetski et al. 2004) on an ABI 3700 sequencer (Applied Biosystems, Foster City, California) at the Advanced Genetic Analysis Center (University of Minnesota, St. Paul, Minnesota). The resulting trace files were aligned and inspected for indels and unexpected stop codons in Sequencher 3.0 (Gene Codes Corporation, Ann Arbor, Michigan), and the verified sequences were deposited in Genbank (DQ272663–DQ272691; DQ310371).
Phylogenetic and Statistical Analyses
A hypothesis of phylogenetic relationships among haplotypes was constructed using Bayesian methods as implemented in MRBAYES 3.1 (Ronquist and Huelsenbeck 2003). The appropriate model of molecular evolution was determined using Akaike's information criterion as implemented in MODELTEST 3.7 (Posada and Crandall 1998). Four heated chains of the Metropolis-coupled Monte Carlo simulation ran for 2 000 000 generations, with trees sampled every 100 generations and the first 1000 trees discarded (as the burn-in, when the chain was searching suboptimal tree space). Two independent runs were carried out in order to identify if the search was carried out in suboptimal local maxima. Sequence-based analyses were also used to examine population history within each of the island populations that had sufficient samples available. Historical patterns of population size were estimated in mismatch distributions using Arlequin 2.0 (Schneider et al. 2000), with results compared to a model of exponential growth in population size. Significance was judged using FS (Fu 1997; test of selective neutrality). DnaSP Version 4.10.2 (Rozas et al. 2003) was also used to calculate the metrics FST (population differentiation) and Nm (migration rate).
Result
We obtained a dataset of 33 haplotypes from the 91 ingroup taxa (plus two outgroup haplotypes). It contained 124 variable sites, and of those, 70 were parsimony informative. MODELTEST specified a GTR + 1 + G model of molecular evolution (i.e., gamma distributed rate variation, a specified proportion of invariant sites, and a general time reversible Markov model with six nucleotide substitution rates). The phylogenetic analyses produced two sets of 19 001 Bayesian trees. The two independent runs produced the same topology, with posterior probabilities nearly identical; the runs were combined to generate Figure 3.
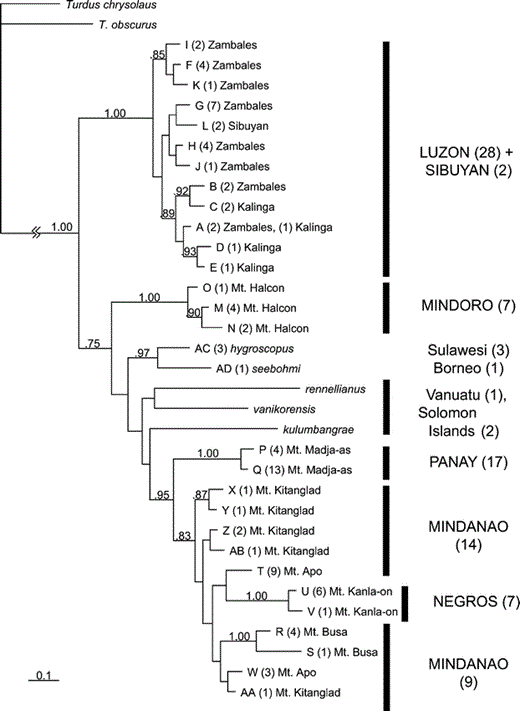
A phylogenetic hypothesis of relationships among haplotypes of Island Thrush, based on 1041 base pairs of the mitochondrial ND2 gene. Letters at the tip of each branch refer to haplotype designations, which are referenced in the Appendix; numbers in parentheses refer to the number of individuals with that haplotype. This phylogeny is a consensus of 19 001 Bayesian trees (from a run of 2 000 000 generations, sampled every 100 steps, with the first 1000 trees discarded as a burn-in) using the GTR + I + G (general time reversible Markov model, with a specified proportion of invariant sites, and gamma distributed rate variation across sites) model of molecular evolution. Numbers beside each node represent posterior probabilities; values less than 0.70 are not shown. Capitalized island names on the right are Philippine islands; other names are islands outside of the Philippines. Neither the Philippine populations as a whole nor Mindanao's populations themselves are monophyletic.
Nodes occurring toward the root of the tree have fairly strong posterior probabilities. Monophyly of the Island Thrush is supported with 1.00 posterior probability, and values greater than 0.95 support the three major clades of Philippine populations (Luzon + Sibuyan, Mindoro, and Panay + Negros + Mindanao). Nodes involving the placement of the Mindoro clade and subspecies outside of the Philippines receive much weaker support, often below 0.50, with the exception of the relationship between hygroscopus and seebohmi (0.97 posterior probability). Weak support for placement of Solomon Islands and Vanuatu samples may be an artifact of long-branch attraction (Bergsten 2005). A topology corresponding to monophyly of all Philippine populations is not present in the set of 99% credible Bayesian trees.
Subspecific taxonomy is only partially correlated with evolutionary history. Subspecies mindorensis and nigrorum are monophyletic; the remaining sampled subspecies (katanglad, kelleri, mayonensis, and thomassoni) are polyphyletic. Polyphyly is related to taxonomic diversity within each Philippine island. The two monophyletic subspecies occur on the two islands that house only a single subspecies (Mindoro and Negros). The two larger islands, Mindanao and Luzon, contain multiple subspecies. Both islands' populations are polyphyletic, as are the subspecies on each. Sibuyan's two sampled individuals from its unnamed population have identical haplotypes and are nested within the Luzon clade. The samples from the unnamed population on Mt. Busa, Mindanao are monophyletic and embedded within other Mindanao and Negros populations. Panay is home to a single, unnamed population that is monophyletic. Sequences from Philippine clades differ from each other by up to 2.6% (uncorrected).
Mismatch distributions for all populations but Mt. Apo, Mindanao conform to expectations under a model of exponential population growth for all populations with adequate sample sizes (i.e., ≥7). Pairwise FST values are presented in Table 1. Additional FST comparisons of areas of biogeographic interest were performed and are presented in Table 2.
The degree of differentiation among populations, presented as pairwise FST values, for Island Thrush populations. “East” refers to two samples from the Solomon Islands and one from Vanuatu. Kalinga is the locality sampled in northern Luzon, and Zambales is the locality sampled in western Luzon; Apo, Busa, and Kitanglad are mountains sampled in Mindanao.
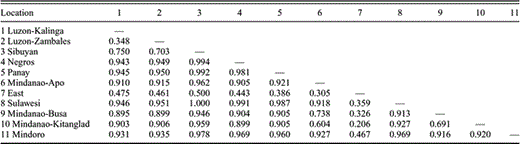
The degree of differentiation among populations, presented as pairwise FST values, for Island Thrush populations. “East” refers to two samples from the Solomon Islands and one from Vanuatu. Kalinga is the locality sampled in northern Luzon, and Zambales is the locality sampled in western Luzon; Apo, Busa, and Kitanglad are mountains sampled in Mindanao.
Island Thrush FST (population differentiation) and Nm (migration rate) values of several distinct biogeographic regions.
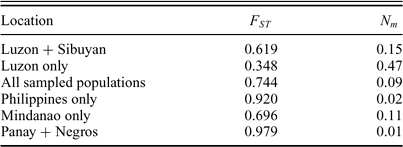
Island Thrush FST (population differentiation) and Nm (migration rate) values of several distinct biogeographic regions.
Discussion
Biogeographic History
Each of the four colonization routes of Dickerson (1928) has testable phylogenetic predictions, all assuming a single colonization of the Philippines and subsequent spread throughout the rest of the archipelago. The phylogenetic hypothesis presented for the Island Thrush based on mitochondrial DNA sequence clearly violates this assumption. There is a division between northern (Mindoro, Luzon, and Sibuyan) and southern (Negros, Panay, and Mindanao) Philippine islands, with southern populations more closely related to birds from Borneo, Sulawesi, Vanuatu, and the Solomon Islands than they are to the northern Philippine populations. Without further sampling from the remainder of the range of this species complex, the timing and direction of colonization events to the Philippines are uncertain, but this phylogeny requires at least two colonizations of the Philippines. At a minimum, this would involve one colonization through route 1 or 2, and a second colonization by route 3 or 4 (Fig. 1). Other studies in this region have reached similar conclusions. There is evidence that both fanged frogs (Evans et al. 2003) and Spizaetus hawk-eagles (Gamauf et al. 2005) colonized the Philippines through multiple events. The assumption of monophyly for Philippine populations is rejected in these studies and indicates that the four proposed colonization routes should not be considered mutually exclusive.
Monophyly of Philippine populations has not been rejected in all published studies. Steppan et al. (2003) suggest a Luzon origin and subsequent southward dispersal for the mouse genus Apomys. Jansa et al. (2006) studied a subfamily of rodents (Murinae) and recovered two clades of Philippine “old endemics” that originated within the Philippines. These studies may differ due to varying taxonomic levels, timescales of diversification, and biases in dispersal abilities. Additional studies of various taxonomic groups and at different taxonomic levels will be needed before a general synthesis of the evolutionary history of Philippine organisms is possible.
Pleistocene sea level changes further complicate hypotheses on the origins of Philippine avifauna. As sea levels dropped, Panay and Negros merged into a single island as new land was exposed (Heaney 1986), but the effect of Pleistocene climate change on highland habitats is unknown (Flenley 1979). Island Thrush populations on Negros and Panay are reciprocally monophyletic nonsister taxa, rejecting a hypothesis of persistent gene flow during the Pleistocene glaciation cycles. Our phylogeny also rejects vicariance as a mode of diversification for Panay and Negros populations, as vicariance assumes sister status for these two populations, yet we find Negros populations are more closely related to Mindanao than Panay populations.
Goodman et al. (1995) discovered a population of Island Thrush on Sibuyan, a small island approximately halfway between Luzon and Panay. They note that Sibuyan birds resemble Luzon populations (T. p. mayonensis and thomassoni) in plumage. Our data explain this resemblance through shared ancestry; the two samples from Sibuyan are embedded within the Luzon clade, and differ by only 0.32%.
Mismatch distributions for all but one island population conform to expectations of exponential population growth. Exponential population growth is observed in many continental populations of birds, particularly those in recently glaciated regions (Pavlova et al. 2003, Zink et al. 2003). The distribution of habitats in the Philippines during Pleistocene glaciations is unknown, but our data suggest that the major islands experienced drastic habitat loss in the recent past.
Values for FST across populations are generally quite high, particularly when compared to those of mainland passerines. Zink (2004) predicted that tropical bird populations should show high FST compared to temperate populations. Within Luzon, the FST value is 0.348 (0.619 if Sibuyan is included), a value that is lower than expected for tropical regions and similar to those from temperate regions. These FST values are dramatically higher than those of the bats of genera Cynopterus and Haplonycterus studied by Peterson and Heaney (1993), which reflect very low gene flow among islands, regardless of Pleistocene affinities. Similarly, the bat results demonstrate isolation of Sibuyan populations, whereas our analysis finds the Sibuyan population of Island Thrush to be the most recently derived of the island populations. The gene flow results of Peterson and Heaney (2003) were explained by relative vagility, but this explanation does not apply to Island Thrush. Our focal species is clearly adept at overwater dispersal, as it has established itself throughout an enormous geographic range including many remote oceanic islands. However, Island Thrush populations no longer experience gene flow among most islands after colonization, leading to island-by-island differentiation.
Plumage Patterns
With strikingly similar plumage patterns found repeatedly in disparate parts of its range, the distribution of plumages of Island Thrush is a challenge to explain in a historical context without a phylogenetic framework. A simplified version of our phylogenetic hypothesis is shown in Figure 4, demonstrating that convergence of plumage types has occurred several times in Philippine populations of Island Thrush. Individuals of subspecies mindorensis and katanglad each have rufous bellies with white centers, brown heads and breasts, and striped crissa. These subspecies are not in close geographic proximity to each other, and our results demonstrate that they are not each other's closest living relatives. Similarly, the thomassoni subspecies most closely resembles nigrorum and kelleri; individuals of all of these subspecies have dark brown backs and wings and lighter brown heads and breasts. Their belly feathers are variably shaded in brown, but all lack white central feathers as well as striped crissa. Once again, the taxa with similar appearances are not each other's closest living relatives. Regardless of which plumage states we might treat as the primitive condition, it is clear that there is convergence of plumage patterns within Philippine populations.
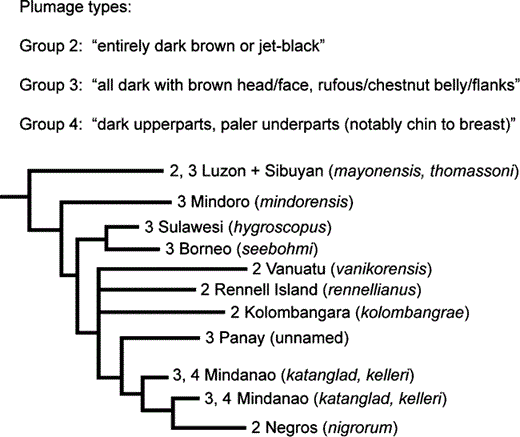
Convergence of plumage types in the Island Thrush. This simplified phylogeny represents branch lengths and topology from Figure 3, collapsing structure among haplotypes within a clade as well as nodes with less than 0.50 posterior probabilities. Plumage type descriptions are quoted from Clement (2000).
Conclusions about convergence are sensitive to weakly supported nodes. However, nigrorum, thomassoni, and the three taxa from the Solomon Islands and Vanuatu (all with plumage type 2; Fig. 4) cannot be sister taxa even when all nodes with less than 0.95 posterior probabilities are collapsed. The same is true for populations with plumage type 3, which is widespread throughout many Philippine islands as well as in Borneo (hygroscopus) and Sulawesi (seebohmi). Furthermore, plumage types 3 and 4 co-occur within two nonsister clades on Mindanao, and plumage types 2 and 3 co-occur within the Luzon + Sibuyan clade. Plumage converge is widespread in birds, with nearly identical plumages in distantly related lineages documented in disparate avian lineages including woodpeckers (Weibel and Moore 2005), wagtails (Pavlova et al. 2003), and orioles (Omland and Lanyon 2000). Plumage convergence in island situations has been documented in Micronesian monarchs (Pomarea spp.; Cibois et al. 2004) and Indian Ocean bulbuls (Hypsipetes spp.; Warren et al. 2005). The Golden Whistler (Pachycephala pectoralis) may represent a comparable case of plumage diversity and convergence in the southwestern Pacific. However, its plumage convergence patterns are obscured by taxonomic confusion, with Mayr and Diamond (2001) considering the species to comprise approximately 66 subspecies, whereas the complex is split into many allopatric species by Dickinson (2003), with P. pectoralis containing only seven subspecies. As with the Island Thrush, a well-sampled phylogenetic approach is needed to understand the origin and maintenance of plumage convergence. The mechanism for plumage convergence may be as simple as a single amino acid replacement (e.g., MC1R in three avian orders [Mundy 2005]), but a robust understanding of the selection regime producing these convergences will require more study integrating genetic and ecological data.
Taxonomic and Conservation Implications
The unnamed Panay population of Island Thrush is a well-supported clade in our analyses and merits taxonomic recognition. It is, however, unclear whether this population and other clades recovered in our phylogenetic hypothesis (including the unnamed population from Mt. Busa) are best treated as biological species or subspecies and whether they constitute phylogenetic species. With a combination of plumage convergence, polyphyletic subspecies, polyphyly of some island populations, and several weakly supported nodes in our phylogeny, it is premature to reassess species limits within this complex. A more thoroughly sampled study of this complex is needed to revisit species limits. Peterson (2007) has initiated this range-wide approach using morphometrics and plumage patterns and discovered 38 diagnosable units (not including Panay, which he did not sample). This approach is admittedly preliminary and should be augmented with other morphological, vocal, ecological, and molecular characters to refine the taxonomy of this species complex.
Within the Philippines, the Island Thrush is commonly found above 1000 m in forest and edges (Kennedy et al. 2000). However, conservation importance changes throughout its range, with some subspecies thought extinct (e.g., pritzbueri in Vanuatu, mareensis on the Mare Islands, vinitinctus on Lord Howe Island) and others rare or not recently recorded (e.g., xanthopus in New Caledonia and poliocephalus on Norfolk Island; Clement 2000). Most biodiversity prioritization efforts ignore subspecies, and the Island Thrush is not even mentioned in the important global assessment by Stattersfield et al. (1998). In general, subspecies names are often misleading as taxonomic categories with predictive values, especially in light of conservation priorities (Zink 2004). Those of the Island Thrush are no exception, and until taxonomic issues are revised for this complex, we suggest following the preliminary data of Peterson (2007) for prioritization of evolutionarily significant units (Moritz 1994) worthy of conservation attention.
Acknowledgments
We thank the field collectors responsible for these tissues, and C. Blake, Z. Cheviron, J. Feinstein, R. Genheimer, S. Hackett, S. Matter, F. Sheldon, P. Sweet, and D. Willard for tissue loans and specimen information from AMNH, FMNH, CMNH, and LSUMZ. Sequences shared by G. Voelker were crucial to this project. R. C. Rago-Blackwell, T. V. Line, S. Drovetski, and M. Westberg provided lab assistance. This manuscript benefited from discussions with F. K. Barker and A. Pavlova and comments by E. Burtt Jr., D. S. Dobkin, S. Lanyon, S. Sealy, A. Simons, R. Zink, and four anonymous reviewers. A. Peterson contributed information on references. This project was supported by Dayton and Wilkie Natural History Funds awarded to AWJ. The majority of the tissues from Philippine populations of Island Thrushes were obtained by the National Museum of the Philippines/Cincinnati Museum of Natural History Philippine Biodiversity Inventory with support from the John D. and Catherine T. MacArthur Foundation and the generosity of numerous friends of RSK and CMNH.
Literature Cited
Appendix
Sampling localities and museum information for the 82 Island Thrush individuals used in this study. All tissue samples are associated with voucher specimens. Birds deposited in the National Museum of the Philippines or in Cincinnati Museum of Natural History are listed with RSK's field numbers, as some of these birds are on permanent loan to CMNH; others have not yet been catalogued. FMNH refers to the Field Museum of Natural History; AMNH is the American Museum of Natural History (DOT is their tissue catalog number, and RWD is the field collector); LSUMZ is the Louisiana State University Museum of Natural Science. Haplotype names correspond to those in Figure 3.
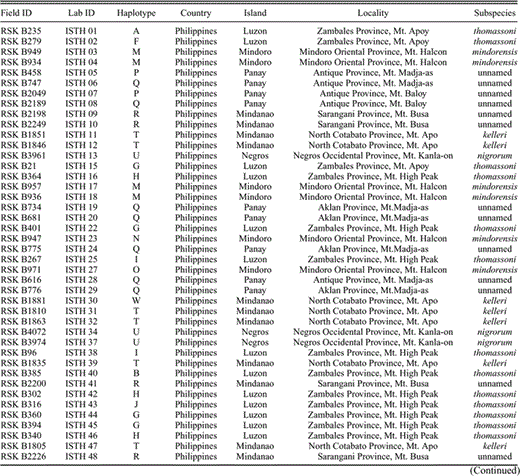
Sampling localities and museum information for the 82 Island Thrush individuals used in this study. All tissue samples are associated with voucher specimens. Birds deposited in the National Museum of the Philippines or in Cincinnati Museum of Natural History are listed with RSK's field numbers, as some of these birds are on permanent loan to CMNH; others have not yet been catalogued. FMNH refers to the Field Museum of Natural History; AMNH is the American Museum of Natural History (DOT is their tissue catalog number, and RWD is the field collector); LSUMZ is the Louisiana State University Museum of Natural Science. Haplotype names correspond to those in Figure 3.
Conitnued.

Conitnued.

{kind=link}
{kind=link}
{kind=link}
{kind=link}