





Abstract
The importance of flooding to birds that nest in bottomland hardwood forests is poorly understood, in part because floods are difficult to predict. A large flood occurred in 2002, during our study of cavity-nesting Wood Ducks (Aix sponsa) in the Mississippi River floodplain. Nests were depredated prior to and after, but not during, the four-week flood. This observation prompted a retrospective analysis of flooding, nest predation, and nest success for our three-year study period of 2000–2002 and with a dataset extending back to 1995. Nest predation was lower and nest success was higher in the floodplain during floods. Flood conditions did not significantly affect nest success or nest predation rates in the adjacent upland, but nest success in upland forest was higher than in the floodplain during dry periods. Historically, spring flooding of riparian areas may to some extent have mitigated nest predation in bottomland forests.
La importancia de las inundaciones para las aves que nidifican en los bosques ribereños de maderas duras está poco entendida, en parte porque las inundaciones son difíciles de predecir. Una gran inundación ocurrió en 2002, durante nuestro estudio de nidos en cavidades de Aix sponsa en la planicie de inundación del Río Mississippi. Los nidos fueron depredados antes y después de la inundación, pero no durante las cuatro semanas de inundación. Esta observación originó un análisis retrospectivo de la inundación, la depredación de nidos y el éxito de nidificación en el período de estudio de 2000 a 2002, combinado con una base de datos que se extendía hasta 1995. La depredación de nidos fue menor y el éxito de los nidos fue mayor en la planicie de inundación durante las inundaciones. Las condiciones de las inundaciones no afectaron significativamente el éxito de nidificación o las tasas de depredación de los nidos en las tierras altas adyacentes, pero el éxito de nidificación en los bosques de tierras altas fue mayor que en las planicies de inundación durante los períodos secos. Históricamente, las inundaciones de primavera de las áreas ribereñas pueden haber mitigado en alguna medida la depredación de los nidos en los bosques ribereños.
Many studies have demonstrated the influence of environmental variables on avian nest success. Physical and landscape variables, such as nest height (Wilcove 1985, Yahner and Scott 1988) and distance to edge, strongly influence predation rates Gates and Gysel (1978). Such variables are generally constant throughout a nesting attempt and are easily measured. However, climatic variables typically vary unpredictably throughout nesting attempts and also affect nest fates. Such variables include local precipitation and flooding events, which affect water levels surrounding nests of birds in wetland habitats.
Most studies of water levels and nest success have focused on marsh-nesting birds (Picman et al. 1993, Jobin and Picman 1997, Sánchez-Lafuente et al. 1998). Water depth was inversely related to nest predation in artificial over-water passerine and waterfowl nests (Jobin and Picman 1997, Sánchez-Lafuente et al. 1998), and the proportion of seasonal and semipermanent wetlands holding water was inversely related to predation of ground-nesting ducks (Johnson et al. 1989). Although water in marshes was thought to have inhibited predation in these studies, rising water levels also inundate the nests of species that nest on or near the ground (Ohlendorf et al. 1989, Marshall and Reinert 1990).
By comparison, studies of the effects of water levels on nest predation of birds that nest in floodplain forests are limited, despite the historical and ecological importance of flooding in these ecosystems. Kennamer (2001) documented greater production during wet years in a population of Wood Ducks (Aix sponsa) using nest boxes, in part due to less predation. Hoover (2006) found that high water levels also deterred nest predation in Prothonotary Warblers (Protonotaria citrea).
Much of our understanding of the impact of floods on animals is anecdotal or based on correlative data that are opportunistically obtained when flood events occur during ongoing studies (Hunter et al. 1987, Flaspohler 1996, Knutson and Klaas 1997). Data collected before, during, and after such events is valuable for examining relationships between water levels and predation. We observed a large flood event while studying the nesting biology of Wood Ducks in a bottomland hardwood forest near the Mississippi River in 2002. Our objectives here are to: 1) compare predation rates and nest success between floodplain and upland habitats with and without flooding, 2) to determine whether predation is random with respect to flood events, and 3) to retrospectively examine the relationship between annual Wood Duck nest predation and flooding during 1995–2002. This study adds to current data on the effects of flooding on birds, as well as furthering our limited understanding of nest predation on Wood Ducks that nest in natural cavities.
Methods
Our study was conducted adjacent to the Mississippi River in Union County, southwestern Illinois. The study area included 3217 ha of floodplain forest and cropland in the Union County Conservation Area and 2289 ha of scrub-shrub and forest swampland in LaRue Swamp Research Natural Area. Forested areas of the floodplain were located in low-lying areas and were the first to flood. Higher elevation portions of the floodplain have been converted to agriculture. Upland habitats adjacent to each of these areas included the Shawnee National Forest. Ryan et al. (1998) provide a detailed description of the study area.
Wood Ducks were trapped with swim-in traps. Radio-collars were attached to females using methods described by Montgomery (1985). We located nest trees by following 130 radio-marked birds to nest sites (134 trees; Ryan et al. 1998, Zwicker 1999, Roy Nielsen et al. 2006a), through searching random plots established for another study (six trees; Zwicker 1999), and through observations of females and feathers at cavity entrances (three trees). We used standard rock-climbing gear to ascend nest trees and inspect cavities. Cavities were inspected with camera equipment on a 50-foot pole in 2000 (Sandpiper Technologies, Inc., formerly Christensen Designs, Manteca, California) whenever logistically possible. Nest trees initially identified by following radio-marked birds were visited in subsequent years to find additional nests. Nests were monitored by tracking radio-marked nesting females 2–3 times per week. Nests were revisited when females were located off their nests more than once during the same week, or after their expected hatching date as determined by egg candling Hanson (1954) or onset of incubation. Nests were found at various stages of laying and incubation in 1995–1998, but eggs were not always candled, especially during earlier years. We were less successful at identifying nest predators during 1995–1998 because we did not immediately revisit nests after abandonment or on expected hatching dates (Ryan et al. 1998, Zwicker 1999). Nevertheless, we typically revisited nests or relocated radio-marked females within one week after termination of nesting so nest success was readily determined. During 2000–2002, we candled ≥1 egg from all clutches (except for a few cases in which cavity depth in excess of 0.6 m made safe return of eggs to nests a concern) and focused on recovering egg remains for genetic analyses immediately after nesting (Roy Nielsen et al. 2006a). Consequently, we were more successful at identifying nest predators and used data from 2000–2002 to conduct more detailed analyses of timing and sources of nest predation and nest success. No data were collected in 1999.
Timing of nest predation was estimated as the date predators were observed at previously active nests or as the midpoint of the interval between successive nest checks Mayfield (1961). Predation was distinguished from nest abandonment by the following criteria: abandoned nests were not missing eggs, did not contain partially or completely eaten eggs in the nest or around the bottom of the tree, and snakes were not present in the nest cavity (we never encountered other predators at the nest). Predators were identified from egg remains according to methods described in Bellrose and Holm (1994). Briefly, woodpeckers puncture eggs with their bills, snakes consume eggs whole, squirrels cut eggs on one end, and raccoons compress eggs, creating an inward fold of the eggshell membranes. Although Larivière (1999) expressed compelling skepticism about the validity of using egg remains to identify nest predators, the number of predators able to access cavity nests up to 21 m above the ground is considerably fewer than predators of ground nests. This simplified identification of nest predators considerably. Furthermore, egg membranes regurgitated by snakes, raccoon hair, and the presence of predators at nests were often used in conjunction with egg remains to identify predators.
Determination of Flood Events
Selle (1999) measured water depths in the floodplain during 1996–1997 and found that depths were significantly associated with gauge heights of the Mississippi River and local precipitation. We examined the timing of predation relative to flood events based on information provided by the Clear Creek Drainage District Office. Our study area begins to flood when the river is above flood stage (9.7 m) at Cape Girardeau, Missouri (23 km from the study site) and significant rainfall occurs (M. McLane, Clear Creek Drainage District Office, pers. comm.). Groundwater seepage contributes to flooding if the river exceeds flood stage for more than two weeks. We obtained gauge heights for the Mississippi River at Cape Girardeau, Missouri, from the U.S. Army Corps of Engineers. Precipitation data for Anna, Illinois (13 km from the study site), was obtained from the National Weather Service. We defined flooding events as periods following ≥2.5 cm of rainfall when the river was at flood stage, and also as periods when the river exceeded flood stage for >2 weeks. This method produced estimates of flooding that agreed with our observations of flooding in the study area.
Statistical Analyses
We randomly simulated predation events 5000 times using nests that were under observation in the floodplain each week to determine whether predation events were random with respect to flooding during 2000–2002. We compared the number of times that predation occurred during flood events to the randomized distribution and rejected the null hypothesis if the observed value was in the lowest 5% of the null distribution Manly (1997). Because randomization tests determine whether the data are likely to have resulted from chance but cannot be easily generalized to a population Manly (1997), we used a chi-square test to examine differences in nest survival (1 − (predation events)*(nest exposure weeks)−1) for flooded and nonflooded periods and for upland and floodplain habitats for six weeks of laying and incubation (Mayfield 1961, Johnson 1979). Similar randomization and chi-square analyses were conducted for upland nests. We used logistic regression to test the association between number of nest visits and incidence of nest predation, to determine whether our activity at the nest attracted predators.
We used least squares regression to examine whether annual predation rates could be predicted from river gauge heights at crest and rainfall received when the river was above flood stage during 1995–2002. We examined the correlation between independent variables using a Pearson correlation. Floodplain and upland habitats were considered separately and the slope and elevation of the regression lines were compared using t-tests Zar (1996). Randomizations were done with PopTools for Microsoft Office Excel. Mayfield nest success and comparisons were done with MICROMORT Heisey and Fuller (1985) and CONTRAST (Hines and Sauer 1989, Sauer and Williams 1989). All other statistics were done with SPSS 10.0.1 SPSS (1999). Means are reported with standard errors, and the significance level was set at α = 0.05.
Results
During 2000–2002, we located nests of radio-marked females after nests had been incubated for 6.2 ± 1.6 days (median = 5 days, range = −4 to 19, n = 16). We located nests of females without radios during day 4.7 ± 1.9 of incubation (median = 1.5 days, range = −8 to 30, n = 24). Nests monitored with telemetry were checked visually by climbing trees every 14.6 ± 1.7 days (median = 17 days, range = 1–28, n = 31). We climbed nest trees of females without radios every 8.6 ± 0.8 days (median = 5 days, range = 1–32, n = 87). The number of nest visits was not associated with nest fate (x ¯ = 1.4 ± 0.4 visits to depredated nests, x ¯ = 1.8 ± 0.3 visits to nests that were not depredated, odds ratio = 1.1, χ21 = 0.8, P = 0.37).
Nest fates were determined for 223 nests (97 floodplain and 126 upland) during 1995–2002; 63 nests (28%) were depredated. During 1995–1998, 35 of 134 nests were depredated, although few nest predators were identified during these early years of the study when nest monitoring was less intense (only two black rat snakes [Elaphe obsoleta obsolete], three raccoons [Procyon lotor], and two avian predators were recorded). In contrast, predators were identified for all 28 depredated nests of the 89 total nests during 2000–2002. Predators included black rat snakes (11), raccoons (13), squirrels (2), and avian predators (2). Snakes (12; two found in one nest) and snake excrement (1) were observed at nests, although we could not exclude the possibility that predators arrived at nests following nest abandonment or previous predation attempts. Raccoon hairs on three cavity entrances and two females killed on nests corroborated evidence based on egg remains. Three nests were depredated but still successful. Four females renested after failing in an earlier nest attempt.
Ten flood events occurred while nests were under observation during 1995–2002. The average duration of flood events was 17.4 ± 4.0 days. Gauge height at crest and precipitation while the river was at flood stage were correlated (r2 = 0.71, P = 0.04), so only gauge height was included in the regression. The height of the Mississippi River at crest was inversely related to annual nest predation for Wood Ducks in the floodplain (r2 = 0.78, y = −0.07x + 0.63, df = 6, P = 0.009; Fig. 1), but was not strongly related to predation rates in the upland (r2 = 0.45, y = −0.03x + 0.24, df = 6, P = 0.10). Crest heights were highest in 1995 and 2002, and lowest in 2000. The slope of the regression line was greater for the floodplain than the upland for 1995–2002 (t1,10 = −1.9, P < 0.05), as was the elevation (t1,11 = 8.2, P < 0.001). Flood waters did not reach the entrances to any cavities during any flood event. However, we did observe two instances in which cavities filled with water during floods due to water movement through the tree. Therefore, nest abandonment in response to flooding was negligible (2 of 223 nests, <1%). Other reasons for abandonment (e.g., female died while off nest, squirrels moved into cavity) are reported by Roy Nielsen et al. (2006b).
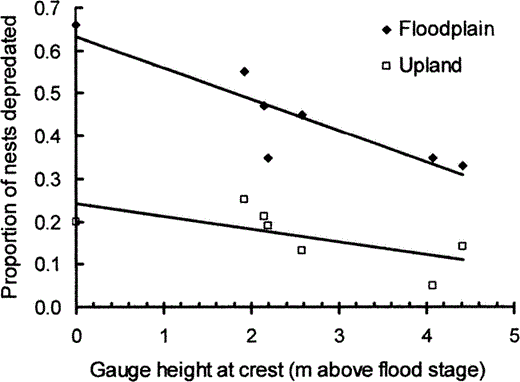
The proportion of Wood Duck nests depredated annually decreased as river height at crest increased above flood stage at Cape Girardeau, a station near the study site in southern Illinois, 1995–2002.
Data were not collected in 1999. Analyses are based on 97 floodplain and 126 upland nests.
During 2000–2002, nest success was higher in the upland (0.54 ± 0.04) than the floodplain (0.26 ± 0.03) during dry periods (χ21 = 21.9, P < 0.001), but nest success was higher in the floodplain (0.65 ± 0.06 vs. 0.50 ± 0.48) during floods (χ21 = 3.7, P = 0.05). Weekly predation in the floodplain during flooded periods (0 events in 29 nest exposure weeks, or 0%) was much lower than during nonflooded periods (17 events in 85 nest exposure weeks, or 20%, including two partial predation events), which increased nest success during floods (χ21 = 33.5, P < 0.001). Predation on upland nests during the same periods of flooding (0 events in 46 nest exposure weeks, or 0%) was similar to that during nonflooded periods (3 events in 82 nest exposure weeks, or 4%, including one partial predation event), as was nest success (χ21 = 0.4, P = 0.51). Predation events in the floodplain were less likely during floods than expected by chance (P = 0.007, 0 of 17 predation events occurred during floods; Fig. 2). Predation on upland nests during the same period was random with respect to flooding (P = 0.69), with predation during floods (n = 0) similar to that during nonflooded periods (n = 3).
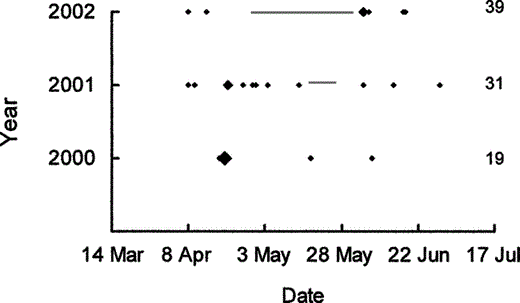
Wood Duck nest predation did not occur during floods in a floodplain in southern Illinois during 2000–2002.
Gray lines represent periods of flooding and diamonds represent predation events. Larger diamonds indicate two or three predation events. The total number of nests each year is given to the right of the chronology. Nests that were being depredated at the time of initial discovery are included (n = 6) but nests depredated before discovery (n = 2) are excluded.
Discussion
Predation ceased during a major flooding event in 2002 and prompted our retrospective analysis of flooding and predation during 1995–2002. Overall, predation on Wood Duck nests was lower in upland areas than in the floodplain, but predation decreased in the floodplain as flood stage increased. A possible trend toward lower predation in upland areas during flooding was also detected, but was not significant. Higher nest predation in the floodplain than the upland is consistent with the results of Robb and Bookhout (1995), who found that Wood Duck nests located farther from water had higher nest success. Raccoons forage primarily in wetlands Fritzell (1978), which may contribute to higher densities or predator activity in the floodplain. Black rat snakes, on the other hand, utilize edge habitats (Weatherhead and Charland 1985, Blouin-Demers and Weatherhead 2001), which were also more common in the floodplain than in the more contiguous, upland forest. Therefore, predator densities were likely higher in the floodplain in the absence of floods.
Our findings are also consistent with those of Kennamer (2001), who found that Wood Duck production was higher in swamp forest during wet years than dry years, in part due to less predation in wet years. Ducklings produced per female and the number of second broods was also higher in wet years. Although Kennamer (2001) worked with a population that used artificial nest boxes, the consistency between our two studies indicates that these patterns may be able to be generalized to both nest boxes and natural cavities. This conclusion is further supported by Hoover (2006), who found that nest predation was lower for box-nesting Prothonotary Warblers when water levels were high. Similar patterns have also been reported during flood events in studies of marsh-nesting birds (Picman 1988, Arnold et al. 1993, Picman et al. 1993).
There are at least three possible ways in which flooding might deter nest predators: 1) flooding might reduce predator movements and hence reduce encounters with nests, 2) predators may leave flooded areas or die, or 3) predators might switch prey due to changes in prey availability during flooding. Although predator behavior and responses to flooding are poorly understood for snakes Weatherhead and Blouin-Demers (2004), deep water may be a barrier to mammalian predators Jobin and Picman (1997). Cagle (1949) reported that raccoons moved from heavily used marsh areas to levees and trees during floods. In contrast, Gehrt et al. (1993) reported that raccoons did not abandon flooded areas and that flooding did not cause mortality. Variability in predator responses among studies might result from differences in the magnitude of flooding and availability of high ground. Other studies have found bird productivity and nest predation to be related to alternative prey availability (Pehrsson 1985, Summers 1986). Availability of alternative foods might increase during floods if prey becomes concentrated in flood refugia, increasing their densities in nonflooded areas. This mechanism might explain the apparent declining trend in nest predation in upland areas during flooding. Some upland nests were located very near the floodplain and may have been affected by prey concentration or changes in availability of alternative prey.
Avian ecologists need to understand how predators locate nests and how predator communities are affected by flooding before the impact of flooding on bottomland forest birds can be fully appreciated. Birds that nest in bottomland hardwood forests may historically have experienced lower nest predation during spring flood pulses. Water control structures have since altered the natural hydrology of many riparian systems to prevent flooding. Furthermore, raccoon densities were lower historically before large carnivores were extirpated Zeveloff (2002). Because nest predation is the single greatest factor affecting passerine Ricklefs (1969) and waterfowl populations (Klett et al. 1988), hydrological alteration of bottomland hardwood forests may have greatly influenced floodplain-nesting birds.
Acknowledgments
This research was conducted under U.S. Fish and Wildlife Service permit MB022884-0 and state permits W01.0060, W02.0005, and W03.0005 issued to CLRN. This work was also performed in accordance with Animal Care Protocols at University of Missouri–St. Louis (W01-3) and Ohio State University (00A0001). St. Louis Zoo, Delta Waterfowl and Wetlands Research Station, and Sigma Xi Grants in Aid of Research funded this work. Sandpiper Technologies provided an equipment grant (TreeTop Peeper II) that also facilitated this study. C. Nielsen, E. Zwicker, R. G. Anderson, R. Geboy, B. Geboy, S. Price, D. Kruze, and J. Steckel contributed assistance in the field. C. Nielsen made constructive comments on an early version of the manuscript. Special thanks to Alan Woolf and the Cooperative Wildlife Research Laboratory at Southern Illinois University in Carbondale for office space, computer, and clerical support, which greatly facilitated progress on this manuscript. We are grateful to R. Kennamer and anonymous reviewers for their helpful comments on this manuscript.
Literature Cited
Author notes
Present address: Cooperative Wildlife Research Laboratory, Southern Illinois University Carbondale, Carbondale, IL 62901. E-mail: [email protected]

{kind=link}
{kind=link}