





Abstract
Timing of breeding activities by birds hasimportant fitness consequences, as deciding when tonest can affect nest success. For three breedingseasons, we provided supplemental food to Wrentits(Chamaeafasciata) inhabitingsemiarid shrublands. We tested the effects of foodsupplementation on timing of initial egg laying,number of nests attempted, timing of fledging, andlength of the breeding season. We also evaluatedwhether Wrentits timed nest initiation to avoidperiods of greatest predation risk. Our study wasconducted during a period of high interannualvariation in precipitation. Supplemental food didnot advance laying date in Wrentits. During adrought, even Wrentits given supplemental fooddelayed nesting, initiated fewer nesting attempts,fledged young earlier, and reduced the length ofthe breeding season. In a year of average rainfall,supplemental food allowed pairs to continuerenesting late into the summer, after repeated nestlosses from predation. Wrentits did not timenesting to avoid periods of greatest predationrisk. In timing initial breeding, Wrentits appearedto be responding to climatic conditions and toindirect cues that predicted environmentalconditions later in the breeding season rather thanto food availability at the time of egg laying.
El Papel del Alimento, de la Depredación de Nidos Y del Clima en la Temporalidad de las Actividades Reproductivas de Chamaea fasciata
La temporalidad de las actividades reproductivas delas aves tiene importantes consecuencias para laadecuación biológica, ya que ladecisión de cuando anidar puede afectar eléxito de nidificación. Durante trestemporadas reproductivas proveímos conalimento suplementario a individuos deChamaeafasciata que habitanáreas arbustivas semiáridas.Evaluamos los efectos del alimento suplementario enla temporalidad del inicio de la puesta de huevos,en el número de intentos deanidación, en el tiempo de emplumamiento yen el tiempo de duración de la temporadareproductiva. También evaluamos siC.fasciata ajusta el tiempo denidificación para evitar periodos de altadepredación. Nuestro estudio fue conducidodurante un periodo de alta variacióninteranual en la precipitación. El alimentosuplementario no adelantó las fechas depuesta en C.fasciata. Durante unasequía, incluso los individuos deC.fasciata que recibieron alimentosuplementario retrasaron sus tiempos de puesta,iniciaron menos intentos de nidificación,los polluelos salieron del nido más tempranoy se acortó la temporada reproductiva. En unaño con precipitación promedio, elalimento suplementario permitió a lasparejas continuar re-anidando más tardedurante el verano, después de repetidaspérdidas de nidos a causa de ladepredación. C.fasciata no ajustótemporalmente la nidificación para evitarperiodos de alto riesgo de depredación. Encuanto al ajuste temporal del inicio del periodoreproductivo, C.fasciata parece respondermás a las condiciones climáticas y alas señales indirectas que predicen lascondiciones ambientales durante la temporadareproductiva que a la disponibilidad de alimentodurante el tiempo de la puesta de huevos.
Introduction
In birds, the timing of annual breeding activities has important consequences for individual fitness. Individuals must decide when to start nesting and whether and when to renest after a failed nest attempt or successfully fledging a brood (Smith and Marquiss 1995, Verhulst et al. 1997, Christman 2002). Long-term cues such as photoperiod, temperature, and endogenous rhythms can stimulate birds to come into and maintain physiological condition for breeding (Wingfield and Kenagy 1991, Wingfield et al. 1992). Within-season decisions about accelerating, delaying, or terminating nesting attempts, and whether to renest after a previous attempt, can be influenced by factors operating at a smaller scale, such as local food availability, nest predation risk, parental age and experience, local temperature and precipitation, plant phenology, or other environmental cues (Perrins 1970, Morton 1971, Geupel and DeSante 1990, Nager and van Noordwijk 1995, Lloyd 1999).
Many bird species breed in strongly seasonal environments with large fluctuations in available food resources, which can affect individual decisions about timing of reproductive activities. Precipitation and temperature are important drivers of these seasonal patterns in resource availability and can influence the timing and success of nesting activities (George 1987, Rotenberry and Wiens 1991, Lloyd 1999, Chase et al. 2005). In northern temperate zones, food resources tend to be lowest early in the season as winter transitions to spring, and again late in the breeding season (Perrins 1970). In more southerly arid ecosystems there are often sharp peaks in resource availability linked to rainfall (Gibbs and Grant 1987, Jaksic et al. 1997, Polis et al. 1997). Environmental conditions may vary considerably among years, leading to annual variability in the timing of availability of food resources for breeding (Nager and van Noordwijk 1995, Pasinelli 2001, Cresswell and McCleery 2003). Traditionally, food availability has been considered the primary factor driving the timing of breeding activities in birds (Lack 1954, Perrins 1970). Individuals should time nest initiation so that the most energetically demanding part of the nesting cycle, when pairs have older nestlings and fledglings, coincides with periods of greatest food abundance (Lack 1954). Nesting early may ensure that the seasonal peak in food resources occurs during the nestling and fledgling stages. Under this food limitation hypothesis, nesting early may be constrained by low resource availability for egg-laying females (Perrins 1970). In support of this hypothesis, many food supplementation studies conducted on a wide range of bird species have documented earlier laying dates for individuals provided with supplemental food prior to breeding (Källander 1974, Davies and Lundberg 1985, Hörnfeldt and Eklund 1990, Svensson and Nilsson 1995, Aparicio and Bonal 2002).
Fewer studies have considered the role of nest predation in influencing the timing of reproduction (Morton 1971, Wiklund 1984, Young 1994), although nest predation is the leading cause of nest failure in most passerine species (Nice 1957, Ricklefs 1969, Martin 1993). The threat of nest predation can vary over time, making some periods of the breeding season riskier for nesting (Wiklund 1984, Armstrong and Pyke 1991, Brown and Roth 2002). Under this nest predation hypothesis, individuals should time breeding activities to avoid the period of greatest predation risk.
Individuals may face conflicting demands in timing nesting to avoid periods of highest nest predation risk and greatest food limitation. If nest predation declines as the breeding season progresses, early nests that are favored under the food limitation hypothesis have a higher chance of failing from nest predation. In contrast, nests initiated later in the season may face reduced nest predation risk although there may be limited food available for nestlings and fledglings.
We used a combination of extensive observations and experimental food manipulation under variable environmental conditions to assess the effects of food availability and, secondarily, nest predation on timing of nesting in the Wrentit (Chamaea fasciata). Our semiarid study site experiences a strongly seasonal environment and substantial variation in annual precipitation. The breeding season typically begins by mid-March after most annual precipitation has occurred in the form of winter rains. Early nesting may be restricted by a lack of food resources available for laying eggs when weather is cold and rainy or when there is a lack of rainfall to stimulate primary productivity. By June, conditions for breeding at the study site have deteriorated due to a lack of precipitation and high daytime temperatures up to 40°C. Many shrub species become drought-deciduous and the relative abundance of insects, particularly caterpillars, decreases as the summer drought progresses.
Wrentits suffer relatively high rates of nest predation and may persistently renest over their long breeding season (Erickson 1938, Geupel and Ballard 2002, Preston 2004). The Western Scrub-Jay (Aphelocoma californica) is an important predator of Wrentit nests at our study area. Jays depredate nests with both eggs and nestlings, and predation is highest early in the breeding season when jays are nest-building and incubating (Preston 2004, Preston and Rotenberry 2006). We experimentally reduced Wrentit nest predation rates by providing supplemental food to jays in a subset of Wrentit territories. This manipulation increased Wrentit reproductive success and annual fecundity (Preston and Rotenberry 2006). However, we were unable to test the effects of predation on timing of reproduction by Wrentits as individuals could not accurately assess predation risk, since jays remained present and active within all predation treatments. Instead, we relied on observational data to evaluate the timing of Wrentit reproduction in relation to predation risk.
Under the food limitation hypothesis we expected that Wrentits given supplemental food would initiate breeding earlier, attempt more nests, and continue breeding later in the season than control pairs. Under the nest predation risk hypothesis, we expected Wrentits to delay reproduction to avoid the period of highest predation risk. We undertook an experimental manipulation of food and an observational study of predation to examine the relative contribution of these alternatives to reproductive timing.
Methods
Study Species
Wrentits are small (15 g) year-round residents in shrubland habitats along the Pacific Coast of the United States, and are primarily insectivorous during the breeding season (Geupel and Ballard 2002). They are socially monogamous and relatively long-lived. Both males and females participate in all aspects of parental care, which may be considerably extended through the postfledging period (Erickson 1938, Geupel and Ballard 2002, Preston 2004). Wrentits may renest multiple times after repeated nest failures due primarily to predation (Erickson 1938, Geupel and Ballard 2002), with some pairs producing two broods within a breeding season (Geupel and DeSante 1990). At our study site, annual nest success under unmanipulated predation conditions ranged from 29% to 79% during the three years of our study (Preston 2004). In general, nest predation risk decreased as the breeding season progressed. However, Wrentit nest predation risk was also related to stage of the Western Scrub-Jay nesting cycle when the Wrentit nest was initiated. Nests begun when jays were nest-building or preparing to lay eggs were more likely to fail than nests initiated later in the jay nesting cycle (Preston 2004).
Study Site
Our study was conducted at a 40 ha site in the 700 ha Hellhole Canyon Open Space Preserve in northern San Diego County, California (33°13′N, 116°55′W). The site ranges in elevation from 600 m to 690 m, and supports dense Southern Mixed Chaparral vegetation with some interspersed patches of smaller, drought-deciduous coastal sage scrub shrubs (Holland 1986). Climate at the site is characterized as Mediterranean, with cool, moist winters and hot, dry summers. Almost 90% of the annual average precipitation in the region falls from November through March (Western Regional Climate Center 2006).
Experimental Design
Wrentit food supplementation experiments were conducted during the latter half of the 2000 breeding season and over the entire 2001 and 2002 breeding seasons. During the 2000 and 2001 breeding seasons, we provided supplemental food to Wrentits without manipulating nest predation risk. During 2002, we reduced nest predation risk in selected Wrentit territories by providing supplemental food to Western Scrub-Jays that were resident in overlapping territories (Preston 2004, Preston and Rotenberry 2006). As part of our broader research program, in 2003 we also surveyed for banded Wrentits throughout the study area. Incidental to these surveys, we found several active nests and documented breeding behavior. These observations were used to assess timing of initial reproduction in 2003.
Since our research in 2000 did not extend over the entire breeding season, we collected only last nest initiation dates and fledging data for that year. Wrentits were assigned to food supplementation and control territories using a stratified random design, and both treatments were interspersed throughout the study area. Wrentit food supplementation (mealworms [Tenebrio molitor]) began on 1 February in 2001 and 2002, about six weeks before nest building typically begins and sufficiently early to potentially influence timing of first nests. We terminated supplemental feeding for all pairs at the same time on 25 July 2001 and 31 July 2002, well after nestlings had fledged. Food supplementation began on 13 May in the 2000 breeding season, after nesting had begun, and was terminated for each pair as soon as young fledged (30 May–8 July). We filled feeders with mealworms every 1–2 days. We housed mealworms on a mixture of bran and chicken starter or hen laying mash to provide additional calcium to Wrentits (Allen 1989). We monitored feeders (using video cameras and direct observations) to confirm their use by resident Wrentit pairs and to document use by other Wrentits. Food was plentiful for all Wrentits that we supplemented, with more than 1.5 × 106 mealworms used during the two and a half breeding seasons. Details of feeder design and documentation of their use by Wrentits may be found in Preston and Rotenberry (2006).
Data Collection
We monitored the nesting status of 39 Wrentit pairs during the 2000 breeding season, 46 pairs in 2001, and 82 pairs in 2002. Between 1 March and 31 July, each pair was observed an average of 18.5 ± 1.6 (SE) times in 2000, 35.3 ± 2.8 times in 2001, and 25.1 ± 1.2 times in 2002. In 2003 we visited the study site 15 times between 15 February and 26 June to survey for banded birds, with two-thirds of the surveys conducted before 1 April. We mist-netted and banded individual Wrentits with unique color-band combinations from late summer through winter in 2000 and 2001.
Individual nesting chronologies were constructed based on a combination of finding nests and observing diagnostic stage-specific breeding activities and behavior (Erickson 1938, Geupel and Ballard 2002). Because of concerns about increased nest disturbance and predation risks, we minimized the number of visits to a nest. Wrentit nest stages were divided into five categories: nest building and pre-egg laying, egg laying, incubation, nestling, and fledgling. The nest building and pre-egg laying stage started with nest construction and extended until the day before the first egg was laid, which defined the onset of the egg laying period. The nest initiation date was the observed or estimated date that the first egg was laid for a nesting attempt. We defined incubation as beginning when a pair displayed incubation behavior, and this stage often began before the clutch was completed. The nestling stage began with hatching of the first egg and continued until the first nestling left the nest, which marked the onset of the fledgling period. At times it was necessary to estimate the timing of transition from one stage to another and in these cases we used the average length of different nesting stages (Erickson 1938, Geupel and Ballard 2002). Incubation and nestling stages were considered to last 15 days each. We defined the length of the breeding season for each pair as the elapsed time (days) between when the pair began construction of the first nest and when their last nest fledged young or the pair terminated breeding activities.
We recorded the location and number of all potential nest predators, including Western Scrub-Jays, observed while conducting nest searches and behavioral observations of Wrentits. We monitored jay breeding behavior to construct nesting chronologies for pairs of jays within Wrentit territories. Using jay nest initiation date we evaluated whether Wrentits delayed their first nests to avoid the period of highest predation risk when jays within their territory were nest-building or egg-laying. As a second measure of nest predation risk, we used the average number of Western Scrub-Jays observed in a Wrentit territory prior to initial egg-laying. Wrentit nest success was negatively associated with the average number of jays per Wrentit territory at our study site (Preston 2004). We predicted a positive correlation between the average number of jays per territory and Wrentit nest initiation date.
Environmental Conditions
Freezing temperatures are relatively rare at our semiarid study site and summer highs can exceed 40°C. The four years of our study encompassed substantial variation in precipitation for the region (Western Regional Climate Center 2006), and included the driest year recorded (2002) for San Diego in 150 years of record keeping (Bolger et al. 2005). With the exception of 2001, annual rainfall totals between 1999 and 2003 were well below average for two weather stations within 18 km of our study site (Western Regional Climate Center 2006). Annual precipitation totals were 23%–43% of the 18-year average in 2002, an unusually low rainfall year.
At our site, bioyear precipitation (i.e., that falling between 1 August and 31 July; Rotenberry and Wiens 1991, Morrison and Bolger 2002) averaged 47.5 ± 33.4 (SE) cm during the most recent six years (1999 to 2005). During our study, bioyear rainfall ranged from 20 cm (2002 breeding season) to 57 cm (2003), with intermediate rainfall totals in 2000 (26 cm) and 2001 (41 cm). The extremely dry conditions of 2002 resulted in limited primary production at the study site. Based on qualitative observations, vegetation was dessicated, many shrubs and trees died or lost leaves, and few plants flowered or produced seeds and fruits during the 2002 breeding season. While 2000 was also a year of unusually low rainfall, the plant community was not as severely affected as in 2002. It is likely that there was a cumulative effect of two preceding dry years (1999 and 2000) that combined with the extremely low rainfall in 2002 to adversely affect plant growth, reproduction, and survival. Insect abundance also appeared to be reduced in 2002 compared with the other years, especially 2001 and 2003. This is consistent with the findings of a study of shrubland birds at sites located 40–65 km south of our study site during 2001 and 2002 that documented declines in insect abundance and primary productivity in 2002 (Bolger et al. 2005).
Temperatures were variable within and among breeding seasons at our study site. While overall average annual temperatures were within 1°C of long-term averages, there was substantial monthly variation. Generally, temperatures were cool or near average for the first half of all four breeding seasons (January–April), whereas the latter half of the 2000 and 2001 breeding seasons were warmer than the long-term averages and the 2002 and 2003 breeding seasons.
Statistical Analyses
We first tested whether there were differences in treatment effects between manipulated and control predation treatments in 2002. If there were no differences, we pooled these treatments in our food supplementation analyses. We included data from 2000 in fledging analyses only, since food supplementation in that year began after Wrentit pairs had started nesting. Observations from 2003 were only used in a descriptive context to compare first nest initiation dates.
Using data from the food supplementation experiments, we compared the effects of year and food supplementation on variables of interest with mixed model analysis of variance (ANOVA). Wrentit food treatment was classified as a fixed effect and year as a random effect to compare their influence on first nest initiation date, fledging date, breeding season length, and number of renesting attempts. The interaction term between treatment and year was included in the original model; however, if this term was not significant at α = 0.25, we omitted it from the ANOVA model and pooled the interaction and error terms to test the main effects (Howell 1997).
We used multiple regressions to analyze Wrentit nest initiation dates in relation to jay nest initiation dates and year. We pooled Wrentit food treatments in this analysis if food had been shown to have no effect on first nest initiation date. We repeated this analysis with the average number of jays observed within a Wrentit territory prior to first egg laying.
All modeling and statistical analyses were performed with SAS System for Windows software (SAS Institute 1999). Statistical significance was assessed at α = 0.05. Means are presented ± SE.
Results
There were no effects of nest predation risk treatment on any aspect of timing of Wrentit reproductive activities. Therefore, we pooled data across predation treatments for the following analyses.
Timing of First Nests
Nest building began on 15 March in 2001 and on 10 March in 2002, with egg-laying beginning on 30 and 31 March, respectively (Fig. 1). In 2003, Wrentits began building nests in mid-February and one pair initiated egg-laying by 28 February, a full month earlier than in the previous two years. Despite starting at almost the same time in 2001 and 2002, however, the breeding season was otherwise retarded in the drought year of 2002. In 2001, 100% of first nests by Wrentits with and without supplemental food had been initiated on or before 16 April, whereas by this date in 2002 only 54% of pairs had begun laying eggs. On average, Wrentits in both food supplementation and control treatments initiated first nests significantly later in 2002 than in 2001 (mixed model ANOVA: F1,78 = 36.6, P < 0.001; Fig. 1, Table 1). Food supplementation in 2001 and 2002 had no significant effect on first nest initiation date (mixed model ANOVA: F1,78 = 1.0, P = 0.31; Fig. 1, Table 1).
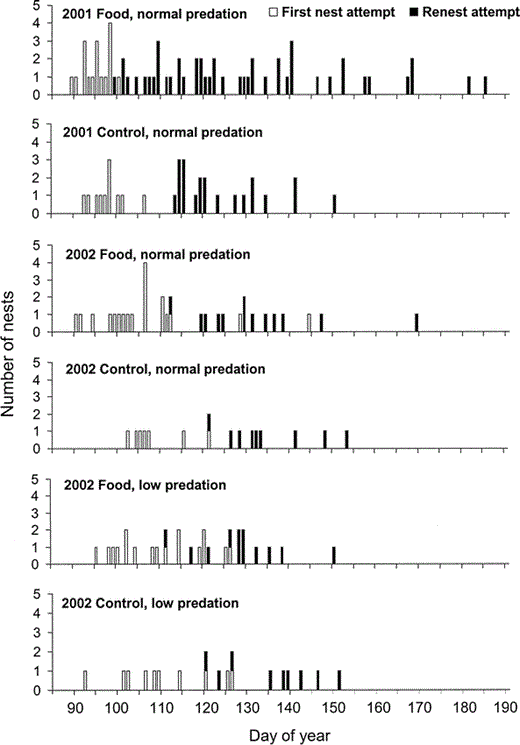
Day of year for first nests and renesting attempts initiated by Wrentits in food-supplemented and control treatments during the 2001 and 2002 breeding seasons in San Diego County, California.
Wrentits initiated first nests earlier and attempted to nest more times over an extended breeding season when given supplemental food (“Food”) in a year of average rainfall (2001) compared with Wrentits without supplemental food (“Control”) and in a year of drought (2002). Experimental reduction of predation risk via supplemental feeding of Western Scrub-Jays in 2002 (“low predation”; Preston and Rotenberry 2006) did not affect timing of Wrentit reproduction compared to unmanipulated controls (“normal predation”). Day 90 = 31 March, day 190 = 9 July.
Mean ± SE values of reproductive parameters describing the nesting chronology of food-supplemented and control Wrentit pairs during the 2000–2002 breeding seasons in San Deigo County, Californa. Wrentits nested earlier, attempted more nests, and had an extended breeding season when given supplemental food (“food”) in a year of average rainfall (2001) compared to Wrentits without supplemental food (“control”) and in a drought year (2002). Wrentits were not monitored in the first half of 2000, so data are unavailable (ND) for most reproductive parameters. Sample sizes are given in parentheses. All dates are given as day of year, where day 90 = 31 March and day 170 = 19 June.

Mean ± SE values of reproductive parameters describing the nesting chronology of food-supplemented and control Wrentit pairs during the 2000–2002 breeding seasons in San Deigo County, Californa. Wrentits nested earlier, attempted more nests, and had an extended breeding season when given supplemental food (“food”) in a year of average rainfall (2001) compared to Wrentits without supplemental food (“control”) and in a drought year (2002). Wrentits were not monitored in the first half of 2000, so data are unavailable (ND) for most reproductive parameters. Sample sizes are given in parentheses. All dates are given as day of year, where day 90 = 31 March and day 170 = 19 June.
Wrentits did not delay nesting to avoid the period of highest predation risk when jays were building nests and laying eggs early in the breeding season. Wrentit nest initiation date was unrelated to Western Scrub-Jay nest initiation date in both years (linear regression, 2001: F1,21 = 0.02, P = 0.89, R2 = 0.001; 2002: F1,47 = 0.02, P = 0.89, R2 = 0.001). The average number of jays observed on a Wrentit territory prior to egg-laying was also unrelated to Wrentit first nest initiation date (linear regression, 2001: F1,17 = 2.3, P = 0.15, R2 = 0.12; 2002: F1,44 = 0.03, P = 0.86, R2 = 0.001).
Breeding Season Length and Number of Nest Attempts
The average length of the breeding season, from nest-building to fledging young or terminating breeding activities, was significantly longer for all Wrentit pairs in 2001 than 2002 (80.3 ± 4.0 vs. 65.5 ± 3.1 days; mixed model ANOVA: F1,79 = 11.6, P = 0.001). In 2002, 98% of nests had been initiated by 31 May, whereas the same proportion was not reached in 2001 until 17 June, 17 days later.
Overall, food supplementation did not affect breeding season length (mixed model ANOVA: F1,79 = 3.5, P = 0.07; Table 1). The interaction between food and year was not significant (mixed model ANOVA: F1,79 = 3.1, P = 0.08), although it could not be excluded from the model. There was no overall effect of food supplementation over the entire study period, because Wrentits provided supplemental food extended breeding in 2001 (ANOVA: F1,32 = 7.3, P = 0.01), but not in 2002 (ANOVA: F1,47 = 2.0, P = 0.17).
Typically, early nests failed and pairs renested several times until they successfully produced fledglings or discontinued attempting to breed. Only one Wrentit pair, a pair provided supplemental food, produced two broods in the same season during our three-year study. Over the 2001–2002 period, supplementing Wrentit food did not affect the number of nesting attempts (mixed model ANOVA: F1,64 = 1.9, P = 0.17; Table 1), although there was a significant difference between years (mixed model ANOVA: F1,64 = 8.0, P = 0.006). Following depredation, all pairs attempted more nests in 2001 than in 2002 (1.9 ± 0.2 versus 1.1 ± 0.1 renesting attempts, respectively). The interaction term between food and year was not significant (mixed model ANOVA: F1,64 = 2.7, P = 0.10), but it could not be excluded from the model. Despite the overall lack of a significant effect of supplemental food on the number of nesting attempts, pairs with supplemental food attempted more nests in 2001 than control pairs (ANOVA: F1,33 = 4.6, P = 0.04). Control pairs attempted to nest up to three times before successfully breeding or discontinuing nesting. In contrast, six pairs given supplemental food attempted to nest four to seven times. In 2002, food supplementation had no effect on number of Wrentit renesting attempts (ANOVA: F1,31 = 0.8, P = 0.37). Pairs in both treatment groups gave up breeding in 2002 if they were not successful after two nesting attempts, with only one pair (with supplemental food) initiating three nesting attempts. The date the last nest of the season was initiated by a Wrentit pair varied among years, with pairs receiving supplemental food nesting later in all three years (Table 1).
Fledging Date
Wrentits fledged young from early May through the first week of August, with most broods fledging in May and June. There were annual differences in Wrentit fledging dates (mixed model ANOVA: F2,127 = 3.8, P = 0.03; Table 1), with earlier fledging in 2002 than in 2000 or 2001. Over the three years, food supplementation did not affect fledging date (mixed model ANOVA: F1,127 = 0.6, P = 0.43).
Discussion
Precipitation and Annual Variation in Timing of Breeding
Annual variation in the timing of Wrentit breeding activities coincided with substantial variability in climatic conditions at our study site. During 2002, an extremely dry year, many pairs delayed nesting compared with 2001, a near average rainfall year. There was early, above average rainfall in the winter of 2002–2003 (57 cm), leading to substantial qualitative changes in vegetation compared with the preceding three years. Early rainfall combined with an exceptionally warm January may have stimulated plant growth and contributed to early production of insects in the 2003 breeding season. Egg laying by Wrentits began a month earlier than in either 2001 or 2002. Other bird species at the study site (including Western Scrub-Jays) were also observed initiating nests earlier in 2003 than in previous years (KLP, unpubl. data).
Numerous studies of birds in arid or semiarid environments, including several conducted within our general region (Ellison 1999, Patten and Rotenberry 1999, Morrison and Bolger 2002, Chase et al. 2005), have documented similar plasticity in timing of first nests and productivity that are correlated with rainfall patterns at both long (months in advance) and short (weeks) timescales (Brown and Li 1996, Lloyd 1999, Grant et al. 2000). In arid environments, precipitation, primary productivity, and the abundance of insect populations are closely linked (Polis et al. 1997, Bolger et al. 2000, Morgan Ernest et al. 2000). Rainfall, and cues associated with rainfall, such as fresh green plant growth, humidity, and barometric pressure, can stimulate breeding, and may predict future conditions for rearing young (Wingfield and Kenagy 1991). The fact that Wrentits with supplemental food did not significantly alter timing of initial nests compared to control pairs implies that the onset of Wrentit reproductive activity is not directly influenced by food supply at the time of egg-laying. Since insect abundance is lowest near the end of the breeding season, when demand for food for nestlings and fledglings is highest, other cues predicting future environmental conditions, such as precipitation and its correlates, may be more important for timing successful reproduction (Eeva et al. 2000).
Opposing Effects of Food Limitation and Nest Predation in Timing of Reproduction
Nest predation risk varied temporally for Wrentits at our site and was highest early in the breeding season (Preston 2004). As a result, food limitation and nest predation exert their strongest effects on Wrentit reproductive success at different times during the nesting cycle. To reduce nest predation, Wrentits at this study site should delay nesting, since early nests are more likely to fail from predation. Opposing this selection gradient to delay breeding is an increasing deterioration of the environment as the season progresses into the period of summer drought. In semiarid shrublands in our region, insect diversity and abundance are highest during spring, lowest during the summer and fall drought, and intermediate in winter after the rains have begun (Bolger et al. 2000, 2005, Burger et al. 2003). Food supplementation did not cause females to breed earlier; rather, it allowed them to lengthen the breeding season during a year of average rainfall. In contrast, in a year of extreme drought, environmental conditions were so severe that both food-supplemented and control females only renested once, and if the second nest was unsuccessful they gave up breeding altogether.
Under the nest predation risk hypothesis, birds should time their breeding to avoid peak predation if nest predation varies in intensity during the breeding season (Morton 1971, Armstrong and Pyke 1991, Sieving 1992, Young 1994). Under this hypothesis, Wrentits should have delayed nesting while Western Scrub-Jays were nest-building and egg-laying. Wrentits in territories with higher densities of jays should also have delayed reproduction relative to pairs with fewer jays in their territory. We did not observe either of these responses. In 2001, Wrentits followed a strategy of initiating nests when predation risk was high and nests were likely to fail, but they continued renesting until successful. In contrast, although nesting was delayed during the drought in 2002, this was not associated with risk of predation within a Wrentit territory as jays also delayed nesting in 2002.
Given the opposing selective pressures of food limitation (greatest late in the season) and nest predation risk (greatest at the beginning of the season), the strategy Wrentits follow in timing their reproductive activity appears to involve flexibility and persistence in nesting in a highly variable environment. An ability to assess environmental cues that accurately predict future conditions for raising offspring appears important for determining when and how much Wrentits allocate to nesting attempts.
Acknowledgments
This work was supported by a Department of Education Graduate Assistance in Areas of National Need fellowship and funding from the University of California, Riverside, Graduate Division and Biology Department. We thank M. Zuk, L. Nunney, R. Redak, and G. Geupel for thoughtful comments on an earlier version of our manuscript. We thank Veronique Rorive for translating the abstract into Spanish. The County of San Diego Parks Department granted permission to work at Hellhole Canyon Open Space Preserve. We dedicate this paper to the Wrentits and Western Scrub-Jays of Hellhole Canyon, which were lost in the southern California wildfires of 2003.
Literature Cited
Author notes
E-mail: [email protected]

{kind=link}