





Abstract
Larger birds are generally more strongly affected by habitat loss and fragmentation than are smaller ones because they require more resources and thus larger habitat patches. Consequently, conservation actions often favor the creation or protection of larger over smaller patches. However, in grassland systems the boundaries between a patch and the surrounding landscape, and thus the perceived size of a patch, can be indistinct. We investigated whether eight grassland bird species with different body sizes perceived variation in patch size and landscape structure in a consistent manner. Data were collected from surveys conducted in 44 patches of northern tallgrass prairie during 1998–2001. The response to patch size was very similar among species regardless of body size (density was little affected by patch size), except in the Greater Prairie-Chicken (Tympanuchus cupido), which showed a threshold effect and was not found in patches smaller than 140 ha. In landscapes containing 0%–30% woody vegetation, smaller species responded more negatively to increases in the percentage of woody vegetation than larger species, but above an apparent threshold of 30%, larger species were not detected. Further analyses revealed that the observed variation in responses to patch size and landscape structure among species was not solely due to body size per se, but to other differences among species. These results indicate that a stringent application of concepts requiring larger habitat patches for larger species appears to limit the number of grassland habitats that can be protected and may not always be the most effective conservation strategy.
¿Afecta el Tamaño Corporal la Sensibilidad al Tamaño del Parche y a la Estructura del Paisaje?
Las aves de gran tamaño generalmente se ven más afectadas por la pérdida del hábitat y la fragmentación que las aves más pequeñas debido a que las aves más grandes requieren una mayor cantidad de recursos y por lo tanto parches de hábitat de mayor tamaño. Consecuentemente, las acciones de conservación generalmente han favorecido la creación o protección de parches de gran tamaño ante la protección de parches más pequeños. Sin embargo, en sistemas de pastizales los bordes entre un parche y el paisaje que lo rodea, y por lo tanto el área percibida del parche, pueden ser difíciles de distinguir. Investigamos si ocho especies de aves de pastizal que difieren en tamaño corporal percibieron una variación en el tamaño de los parches y la estructura del paisaje de una manera constante. Los datos fueron recolectados a partir de muestreos realizados en 44 parches de pradera de pastizal alto del norte entre 1998 y 2001. La respuesta al tamaño del parche fue muy similar entre especies independientemente de su tamaño corporal (la densidad fue afectada muy poco por el tamaño del parche), excepto para Tympanuchus cupido, que mostró un efecto umbral: esta especie no fue encontrada en parches de áreas menores a 140 ha. En paisajes que contenían entre un 0% y un 30% de vegetación leñosa, las especies de menor tamaño respondieron de manera más negativa al aumento del porcentaje de la vegetación leñosa que las especies de mayor tamaño, pero aparentemente por encima de un umbral del 30%, las especies de mayor tamaño no fueron detectadas. Otros análisis revelaron que la variación observada entre especies en las respuestas al tamaño del parche y la estructura del paisaje no se debió solamente al tamaño corporal per se, sino que también a otras diferencias entre especies. Estos resultados indican que la aplicación rigurosa de los conceptos que requieren parches de gran tamaño para las especies más grandes parece limitar el número de hábitats de pastizal que pueden ser protegidos, y en muchos casos ésta puede no ser la estrategia de conservación más efectiva.
Introduction
Larger-bodied animals of many taxa are known to have lower densities and larger home ranges than smaller-bodied animals (Schmid et al. 2000, Haskell et al. 2002) because of greater energy needs and lower density of food resources (Jetz et al. 2004). Thus, it is not surprising that larger species are generally more sensitive to habitat loss and fragmentation than are smaller species (Haskell et al. 2002), and creating or maintaining larger habitat patches is of high priority in many conservation plans. Because conservation of larger species can also protect other species inhabiting the same habitat, larger species are often used as umbrella species (Poiani et al. 2001, Suter et al. 2002). However, using larger-bodied species to determine optimal patch size and landscape structure is complicated by several factors:
The patch-size requirement of a species can vary with resource availability within a habitat patch (Haskell et al. 2002) and with the structure of the surrounding landscape (Bakker et al. 2002, Fletcher and Koford 2002, Winter et al. 2006; Cunningham and Johnson 2006). It is therefore not possible to generalize patch-size requirements across the breeding range of even a single species.
It is more difficult to collect reliable distribution data on rare species than on abundant species (Seoane et al. 2005). Data on the distributions of smaller species—if they are more abundant—might therefore be a more reliable indicator of the habitat needs of a community. However, rare species often have more restricted habitat requirements than more common generalist species. Therefore, the use of more common species as indicator species will only be applicable in those cases in which the needs of a more common species are closely aligned with the needs of the target species.
The size of a habitat patch as defined by humans might not reflect an animal's perception of a patch. This is especially true for species that inhabit open habitats such as native grasslands within agricultural areas, where the edge of a patch is less clearly defined than in patches of other habitat types.
Grassland birds have been the focus of intensive research and conservation because of their consistent and steep population declines in many parts of the world (Peterjohn and Sauer 1999, Donald et al. 2000, Newton 2004). In North America, the Bird Conservation Area concept (Henderson and Sample 1995) provides guidelines for grassland bird conservation, including the establishment of large core areas (>700 ha) of high-quality grassland habitat surrounded by larger expanses of nonforested habitats, such as smaller-grain cropland or hay fields. This habitat configuration is based on the requirements of the Greater Prairie-Chicken (Tympanuchus cupido), a species that is mostly absent from smaller grassland patches and from patches that are surrounded by forested habitat (Ryan et al. 1998, Merrill et al. 1999, Winter and Faaborg 1999) and therefore serves as an umbrella species for the protection of many other grassland birds. However, even though it is desirable to protect larger habitat patches in the form of Bird Conservation Areas, such protection might not be the most cost-effective way to protect grassland birds throughout their range (Davis 2004, Winter et al. 2006).
Recently, we demonstrated that two passerine species nesting in northern tallgrass prairie did not respond consistently to the size of grassland patches, whereas an increasing percentage of woody vegetation surrounding a grassland patch consistently resulted in lower species densities (Winter et al. 2006, also see Grant et al. 2004). The lack of a consistent patch-size effect may have been caused by the small territory sizes of the focal species, because even our smallest grassland patches could easily have contained several territories of either species. In contrast, larger-bodied, nonpasserine species might respond more strongly to patch size as well as to the percentage of trees and tall shrubs in the surrounding landscape, because their territories might extend across a given habitat patch. If larger species are more sensitive to patch size and the percentage of woody vegetation, then a conservation focus on larger prairie patches is justified. However, if such a pattern is not generally true, and larger species also use smaller habitat patches, then the conservation of smaller prairie patches could benefit both smaller and larger species. If smaller and larger species respond similarly to variation in patch size in certain settings, then it would be more efficient to monitor smaller species (found at higher densities) to make management decisions regarding larger-bodied species. In this paper we determined whether eight species of tallgrass prairie birds, four passerines and four nonpasserines, showed similar responses to variation in patch size and the percentage of woody vegetation in the landscape, and whether differences among species were largely the result of variation in body size.
Methods
Between 1998 and 2001 we conducted bird surveys in 44 study plots located in three regions: within Sheyenne National Grassland in southeastern North Dakota and in two regions in northwestern Minnesota close to the cities of Crookston and Glyndon (for a detailed description of study sites and methods, see Winter et al. 2005a, 2005b). Study plots were located within prairie patches that ranged from 2.4 to 1245.6 ha (x ¯ = 226 ha, SE = 52 ha), and the percentage of woody vegetation within a 200 m buffer of the study plots ranged between 1% and 46% (x ¯ = 13%, SE = 2%). The sizes of prairie patches were not strongly correlated with the percentage of woody vegetation within a 200 m buffer (r = −0.20, P = 0.18, n = 44). The size of study plots within the prairie patches varied from 1.5 to 20 ha (x ¯ = 11 ha, SE = 1 ha), depending on the size of the patch within which the study plot was embedded. Only the largest prairie patch contained >1 study plot; the three plots here were >500 m apart. All but two study plots were located within predominantly native tallgrass prairie. Exclusion of those two study plots did not change the results of our analyses (MW, unpubl. data). Study plots were marked with flags or wooden stakes at 50 m intervals along transects that were 100 m apart.
Bird censuses were conducted twice a year from the end of May to early July, between 05:00 and 10:00 CDT, and under conditions of low-to-moderate wind speeds (≤35 kph), no precipitation, and good visibility. During censuses, all singing males were recorded on a map of the study plot, and flight paths were noted to decrease the likelihood of double-counting. If two birds of the same species were not singing, but were in close vicinity to each other and chipping at the observer, we assumed that one of the two birds was a male. The annual maximum count of a species on a plot was used to determine its annual density (number of males per 100 ha). A single observer conducted all counts within a year, with two observers (MW and JAS) alternating in successive years. For analyses we used only individual birds that were detected within the boundaries of our study plots.
As an index of body size, we used male mass (g) derived either from our own study or from studies closest to our sites (Table 1). Our focal species were grassland obligates that occurred regularly in at least some of the study plots: Le Conte's Sparrow (Ammodramus leconteii), Savannah Sparrow (Passerculus sandwichensis), Bobolink (Dolichonyx oryzivorus), Western Meadowlark (Sturnella neglecta), Northern Harrier (Circus cyaneus), Upland Sandpiper (Bartramia longicauda), Marbled Godwit (Limosa fedoa), and Greater Prairie-Chicken.
Species and associated body sizes used to determine if body size influences the effect of patch size and landscape structure on the density of a species in northern tallgrass prairie, 1998–2001. Sample size (n) is the number of birds used to determine average body size.
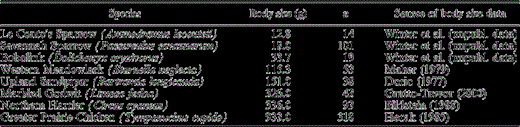
Species and associated body sizes used to determine if body size influences the effect of patch size and landscape structure on the density of a species in northern tallgrass prairie, 1998–2001. Sample size (n) is the number of birds used to determine average body size.
We defined a grassland patch as a continuous stretch of grassland that was not interrupted by four-lane or two-lane roads with disturbed roadsides, or by wooded areas or open-water wetlands that were >20 m wide and extended across >75% of a grassland patch. Grassland patches bordering each other at one corner were considered as two separate patches. Hay-meadows and CRP fields were not considered part of the prairie patch but as part of a treeless landscape.
The size of the grassland patch in which study plots were located was determined from digital orthophoto quadrangles (DOQs) that were imported into TNTmips® Map and Image Processing System, versions 6.6 and 6.7 (MicroImages 2005). The DOQs were from 1991 for Minnesota and from 1997 for North Dakota. We also used these images to determine the percentage of shrubs and trees within 200 m and 1 km buffer zones, where buffer zones included the study plot. The percentage of shrubs and trees within a 200 m buffer zone was highly correlated with the percentage of shrubs and trees within a 1 km buffer zone (r = 0.78, P < 0.001, n = 44 study plots). Assuming that the percentage of shrubs and trees closest to the study plots had a larger effect on the distribution of birds within the study plots than the percentage of shrubs and trees at farther distances, we used only the percentage of shrubs and trees within a 200 m buffer zone in all analyses and refer to this variable as “landscape.”
Statistical Analyses
We used general linear mixed models to determine the factors that best explained variations in bird density, because models included both fixed effects (body size, patch size, and landscape) and random effects (prairie; Littell et al. 1996, SAS 1999). Density data were log-transformed in all analyses, with logcount = ln(count + 1). By modeling prairie as a random effect, we are explicitly assuming that prairie-specific deviations from an overall effect are normally distributed on the log-transformed scale on which the data were analyzed. We examined this assumption by treating prairie as a fixed-effect categorical analysis, in a model that otherwise was identical to the final model (see below), and plotting the prairie-specific regression coefficients as a histogram. The histogram did not deviate from a symmetrical distribution. Thus, we feel confident that the scale-specific normality assumption for the random effect was justified.
We used an information-theoretic approach to fit and rank models (Burnham and Anderson 2002). Previous analyses indicated that grassland bird density varied greatly among prairies (Winter et al. 2005b). We therefore included prairie as random effect in all models. To reduce the number of models tested, we conducted our analyses in two steps (Winter et al. 2005a, 2005b). In a first step, we determined if body size (continuous variable) influenced the effect of patch size on bird density by comparing four models with the following explanatory variables: (1) patch size; (2) body size; (3) patch size and body size; and (4) patch size, body size, and their interaction. We refer to this set of models as “body-size models.” We then used the body-size model that best fit the data, based on Akaike's information criterion corrected for small sample sizes (AICc; Burnham and Anderson 2002), as the base model for a second set of analyses. If more than one model was supported (ΔAICc < 4), we used the most inclusive model, i.e., the model that included the most variables, as the base model. This procedure ensured that all potentially important variables were included in the final model. In the second step, we determined if the density response to the percentage of woody vegetation in the surrounding landscape (“landscape”) was influenced by body size—given the previously determined effect of patch size—by comparing three models: (1) base model (from step 1); (2) base model with landscape variable; and (3) base model with landscape and landscape*body size interaction. From this set, we determined the best-supported model as the final model.
Variation in the response to patch size and to the percentage of woody vegetation structure among birds with different body sizes might not be due to body size per se, but due to differences in ecology and behavior. We investigated this possibility using a similar two-step analysis as described above, but included species as a categorical variable instead of body size as a continuous variable. We compared AICc between the body size and species models, and graphically illustrated the outcome of the two sets of analyses (body size and species models) by plotting the estimated densities derived from each base and final model. The magnitude of the response to patch size and landscape structure appears much greater in species that have high densities than in rare species, because absolute changes in numbers and thus slopes are greater in abundant species. These absolute changes in density do not give a clear impression of the relative magnitude of patch size and landscape effects among species. Therefore, we also plotted the relative changes in density between maximum and minimum percentage woody cover by dividing the estimated maximum by the estimated minimum densities as derived from the final species model.
Results
Models that included species as a categorical variable fit the data better than models that included body size (ΔAICc > 700). Thus, changes in density with patch size and landscape composition are better predicted by a combination of differences among species than by a single type of interspecific variation, body size. For example, the final body-size model predicted that the three smallest species would respond similarly to changes in landscape structure (Fig. 1a); in reality, Savannah Sparrows and Bobolinks were more strongly affected by landscape structure than the smaller Le Conte's Sparrows (Fig. 1b, 2).
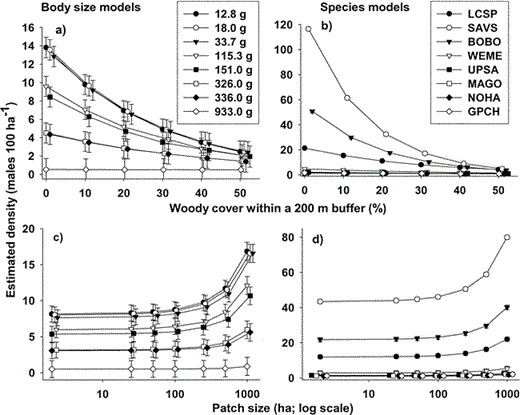
Estimated densities (males per 100 ha ± SE) of bird species nesting in northern tallgrass prairies between 1998 and 2001 decrease with increasing percentage of woody vegetation within a 200 m buffer zone (a and b), and increase with patch size (c and d).
Estimated densities were derived from models that include either body size (“body size models”) or species (“species models”; see Table 2). Error bars for the species models are smaller than the symbol size, and thus are not recognizable. Species include Le Conte's Sparrow (LCSP), Savannah Sparrow (SAVS), Bobolink (BOBO), Western Meadowlark (WEME), Upland Sandpiper (UPSA), Marbled Godwit (MAGO), Northern Harrier (NOHA), and Greater Prairie-Chicken (GRPC). Variations in densities in response to woody vegetation and patch size are better explained by a combination of inherent differences among species (b and d) than by body size alone (a and c).
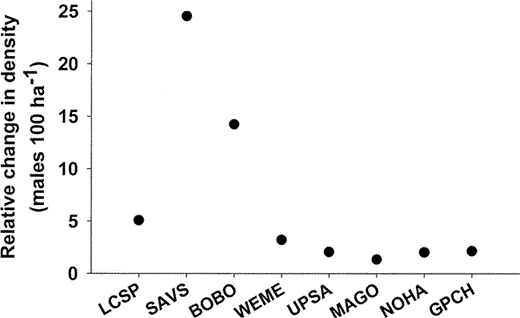
Relative change in densities of species (males per 100 ha) with percentage of woody vegetation (estimated density at lowest woody percentage / estimated density at highest woody percentage) in northern tallgrass prairies, 1998–2001.
Species include Le Conte's Sparrow (LCSP), Savannah Sparrow (SAVS), Bobolink (BOBO), Western Meadowlark (WEME), Upland Sandpiper (UPSA), Marbled Godwit (MAGO), Northern Harrier (NOHA), and Greater Prairie-Chicken (GRPC).
Candidate models used to investigate which factors best explain variation in density of eight bird species nesting in the northern tallgrass prairies of Minnesota and North Dakota, 1998–2001 (n of observations = 1280). Models are ranked from most (ΔAICc = 0) to least supported. Covariates include patch size (“patch”), the percentage of woody vegetation within a 200 m buffer (“land”), and either male body size (“body”: body size models) or species (species models). “Prairie” is included in each model as a random effect because previous analyses indicated that densities varied consistently among prairies (Winter et al. 2005b). The most inclusive patch-size model with a ΔAICc < 4 was used as the base model (Base) for the landscape model, and the most inclusive landscape model was designated as the final model (Final). K is the number of parameters in the model,  is twice the negative value of the maximized log-likelihood function, AICc is Akaike's information criterion corrected for small sample sizes and ΔAICc is the difference in AICc between the best fitting model and model i, and wi is the Akaike weight, which indicates the relative support for each model given the data.
is twice the negative value of the maximized log-likelihood function, AICc is Akaike's information criterion corrected for small sample sizes and ΔAICc is the difference in AICc between the best fitting model and model i, and wi is the Akaike weight, which indicates the relative support for each model given the data.
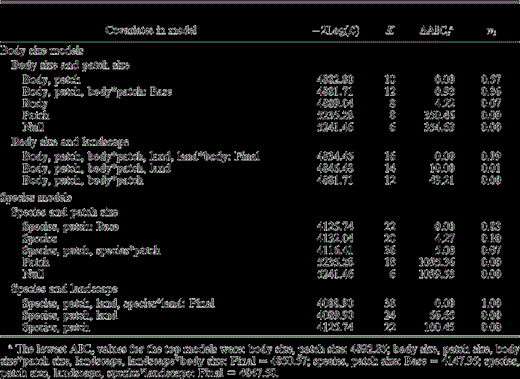
Candidate models used to investigate which factors best explain variation in density of eight bird species nesting in the northern tallgrass prairies of Minnesota and North Dakota, 1998–2001 (n of observations = 1280). Models are ranked from most (ΔAICc = 0) to least supported. Covariates include patch size (“patch”), the percentage of woody vegetation within a 200 m buffer (“land”), and either male body size (“body”: body size models) or species (species models). “Prairie” is included in each model as a random effect because previous analyses indicated that densities varied consistently among prairies (Winter et al. 2005b). The most inclusive patch-size model with a ΔAICc < 4 was used as the base model (Base) for the landscape model, and the most inclusive landscape model was designated as the final model (Final). K is the number of parameters in the model, is twice the negative value of the maximized log-likelihood function, AICc is Akaike's information criterion corrected for small sample sizes and ΔAICc is the difference in AICc between the best fitting model and model i, and wi is the Akaike weight, which indicates the relative support for each model given the data.
Patch size had a negligible effect on bird density, especially at patch sizes below 500 ha (Fig. 1c, 1d), even though patch size and the interaction between patch size and body size were in the most inclusive model with a ΔAICc < 4 (Table 2). The effect of patch size (slope: 0.0007 ± 0.0003 SE) and the interaction of patch size and body size (slope: <0.0001 ± <0.0001 SE) on density was minimal. A 1000 ha increase in patch size only roughly doubled the density of most species (Fig. 1c, 1d). Even larger-bodied species (Northern Harrier, Marbled Godwit, and Upland Sandpiper) occurred in relatively small patches (<20 ha), with the exception of the Greater Prairie-Chicken; this species was absent from prairie patches smaller than 140 ha.
The addition of landscape features to patch-size models greatly improved both the body size and species models, decreasing the AICc value by 42 and 100, respectively (Table 2). Density decreased with increasing percentage of woody vegetation within a 200 m buffer (Fig. 1a, 1b); this decrease varied with body size and among species (Table 2). The relative change in density with percentage of woody vegetation varied by species (Fig. 2), with smaller species showing a stronger response to changes in the percentage of woody vegetation: a 50% decrease in woody cover within a 200 m buffer caused an increase in density that varied by 3%–25% in smaller species, whereas the density of larger species increased by less than 2.2% (Fig. 2). However, if woody cover exceeded 30% within a 200 m buffer, larger species were not detected. Thus, an apparent threshold in woody cover exists for larger species, above which these species do not occur.
Discussion
Our results show that, at least in our study system, we cannot assume that larger species always require larger habitat patches and more open landscapes than smaller species, because models that included body size received less support than models that included species. In fact, within a certain range of woody cover, smaller species were more strongly affected by changes in landscape composition than larger species. Thus, providing suitable habitat for smaller-bodied species can also ensure the protection of larger-bodied species if both the small-bodied species and the species of concern have similar habitat requirements and the amount of wooded habitat is below any thresholds that may be present. A single, larger-bodied umbrella species, therefore, might not always be the ideal subject to guide community-wide conservation decisions (Andelman and Fagan 2000, Roberge and Angelstam 2004); instead, management recommendations need to be adapted to the specific area and species to be managed.
Our findings have several important implications for the conservation of grassland birds in northern tallgrass prairies, and might also be applicable to other systems:
In open landscapes, patch size variation is of relatively little importance even for larger-bodied species. All species but the Greater Prairie-Chicken were found in prairie patches smaller than 20 ha. Thus, the protection and management of small prairie patches can be worthwhile not only for passerines (Davis 2004, Winter et al. 2006; Davis et al., in press), but also for several larger-bodied, nonpasserine species. However, to ensure the occurrence of Greater Prairie-Chickens, some grassland patches need to be at least 140 ha in size even if the patch is located in a relatively open landscape.
Species of different body sizes can have similar responses to patch size and landscape variables. Thus, at least in our study area, large-bodied species seem to be of limited utility as umbrella species. Relying instead on the response of smaller species would have the benefit of more accurate density estimation, because densities of smaller animals are estimated with greater accuracy, if the species is relatively abundant (Seoane et al. 2005).
Even a seemingly small amount of woody vegetation can greatly reduce the number of individuals of a species present in a grassland patch. Any reduction of woody vegetation can thus be beneficial to grassland birds (but see Suedkamp Wells 2005). This observation seems to constitute a general pattern for grassland birds in the northern prairies, since studies elsewhere have reported similar results (Bakker et al. 2002, Fletcher and Koford 2002, Grant et al. 2004; Cunningham and Johnson 2006). In addition, nesting success may be lower for nests placed close to woody vegetation (Johnson and Temple 1990, Winter et al. 2000).
Effective grassland bird conservation requires a balance between the time and money needed and the degree to which high-quality refuges for grassland species can be obtained. In an open agricultural landscape such as our study area, we suggest that a mix of both smaller and larger (>140 ha) prairie patches will likely support all grassland-nesting species. Generally, the protection of larger, continuous areas is preferable. However, a stringent application of concepts that acknowledge the desirability of only larger habitat patches limits the grassland habitats that can be protected and may not always be the most effective conservation strategy during times of limited financial support for conservation.
Acknowledgments
We thank the more than 40 field assistants, whose efforts we greatly appreciate. W. D. Svedarsky of the University of Minnesota in Crookston, R. Julian of the U.S. Fish and Wildlife Service, B. M. and S. Winter and the late G. Yalch of The Nature Conservancy, and B. Stotts of the USDA Forest Service provided logistical help. The Nature Conservancy, U.S. Fish and Wildlife Service, USDA Forest Service, and Minnesota Department of Natural Resources allowed us access to the study sites. J. E. Austin, the late D. P. Fellows, P. J. Pietz, and M. A. Sovada of the Northern Prairie Wildlife Research Center loaned equipment and supplies. B. R. Euliss provided patch sizes and data on the percentage of woody vegetation. The Cornell Laboratory of Ornithology provided working facilities for MW. The manuscript was greatly improved by comments from Todd Grant, Wesley Hochachka, Steve Davis, David Dobkin, Thalia Sachtleben, and one anonymous reviewer. Funding was provided by the U.S. Geological Survey, the U.S. Fish and Wildlife Service, Regions 3 and 6, the American Bird Conservancy, and the Northwest Research and Outreach Center at the University of Minnesota, Crookston.
Literature Cited
Author notes
Present address: Laboratory of Ornithology, Cornell University, 159 Sapsucker Woods Rd., Ithaca, NY 14850. E-mail: [email protected]

{kind=link}
{kind=link}