





Abstract
We report the wing bones of a small passeriform bird from the early Oligocene of France, which are among the earliest fossil remains of Passeriformes in Europe. The specimen is clearly distinguished from oscines, to which all extant European passeriforms belong, and closely resembles suboscine passerines in presumably derived features. If future, more complete specimens support its assignment to the suboscines, it would be the earliest fossil record of this passeriform taxon. Together with other fossils from the early Oligocene of Germany, it indicates that nonoscine passerines colonized Europe before the arrival of oscines.
Resumen
Una Pequeña Ave Paseriforme Tipo Suboscine del Oligoceno Temprano de Francia
Anunciamos el hallazgo de huesos del ala de una pequeña ave paseriforme del Oligoceno temprano de Francia, que está entre los restos fósiles más tempranos de paseriformes en Europa. El espécimen se distingue claramente de los oscines, al cual pertenecen todos los paseriformes europeos vivientes, y se asemeja marcadamente a los paseriformes suboscines en rasgos presumiblemente derivados. Si hallazgos futuros de especimenes más completos apoyan su asignación a los suboscines, sería el registro fósil más temprano de este taxón paseriforme. Junto con otros fósiles del Oligoceno temprano de Alemania, este hallazgo indica que los paserinos no oscines colonizaron Europa antes de la llegada de los oscines.
Passeriformes (passerines or perching birds) is today the most species-rich avian order and comprises more than half of all extant avian species. According to molecular studies, the New Zealand wrens (Acanthisittidae) are the sister taxon of all other passerines, the suboscines and oscines (Barker et al. 2002, Ericson et al. 2002, 2003). All extant European Passeriformes are oscines and the sole Old World representatives of suboscines are Eurylaimidae (broadbills and asities) and Pittidae (pittas).
The hitherto earliest European fossil passerine was described by Mayr and Manegold (2004, 2006) from the early Oligocene of Germany, and is known from a disarticulated skeleton and an additional incomplete skull. The exact phylogenetic affinities of this species are uncertain, and although it most closely resembles extant suboscines it may be outside the crown group of Eupasseres, the clade that includes suboscines and oscines (Mayr and Manegold 2004, 2006). The only older fossil record of a passeriform bird, from the early Eocene of Australia (Boles 1995, 1997), is based on fragmentary remains of two bones and needs to be confirmed by additional material. All other fossil passerines are of Upper Oligocene or younger age (Mayr 2005).
The presence of a passeriform bird in the early Oligocene of the Luberon area of Southern France was previously mentioned by Roux (2002), but that specimen is still undescribed. Here, we describe the distal portion of the wing skeleton of a small passerine from this locality, which closely resembles the corresponding bones of extant suboscines.
Methods
The fossil specimen is deposited in Forschungsinstitut Senckenberg (SMF), Frankfurt am Main, Germany. Osteological terminology follows Baumel and Witmer (1993) and Ballmann (1969).
Results
Systematic Paleontology
Passeriformes (Linnaeus, 1758)
cf. Suboscines (sensu Ericson et al. 2003)
Family, genus, and species indeterminate
Referred Specimen
SMF Av 504, right manus (Fig. 1).
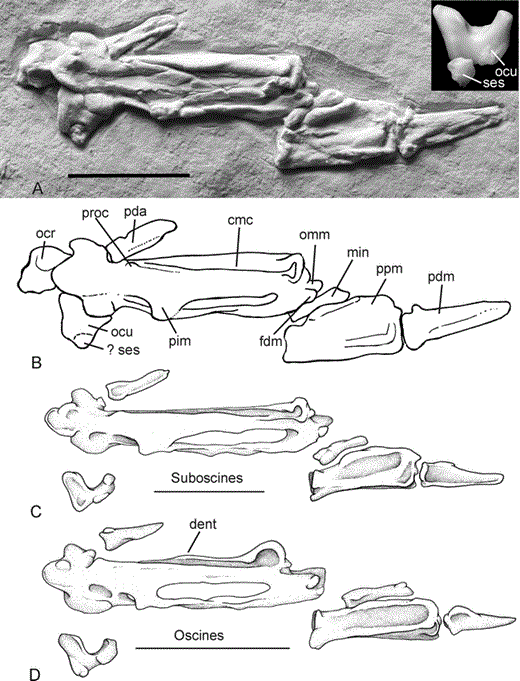
Figure 1. Distal part of right wings for comparison; the carpometacarpus is seen in dorsal view, the distal phalanges in ventral view. A: the fossil passerine SMF Av 504; inset: os carpi ulnare of the Superb Lyrebird (Menura novaehollandiae, oscines, Menuridae); B: schematic labeled drawing; C: African Broadbill (Smithornis capensis, suboscines, Eurylaimidae); D: Yellow-faced Honeyeater (Lichenostomus chrysops, oscines, Meliphagidae). Abbreviations: cmc = carpometacarpus, dent = processus dentiformis, fdm = facies articularis digitalis minor, min = phalanx digiti minoris, ocr = os carpi radiale, ocu = os carpi ulnare, omm = distal end of os metacarpale minus, pda = phalanx digiti alulae, pdm = phalanx distalis digiti majoris, pim = processus intermetacarpalis, ppm = phalanx proximalis digiti majoris, proc = process at proximal end of sulcus tendinosus, ses = sesamoid bone. Fossil coated with ammonium chloride to enhance contrast; note that, for diagenetic reasons, the bones of the fossil appear wider than they probably were originally. Scale bars equal 5 mm
Locality and Horizon
Luberon area, Southern France; Lower Oligocene (Mourer-Chauviré 1996).
Measurements
Carpometacarpus: maximum length, 10.4 mm; length from proximal end of trochlea carpalis to distal end of os metacarpale majus, 9.9 mm. Phalanx proximalis digiti majoris: length, 4.8 mm. Phalanx distalis digiti majoris: length, 4.0 mm.
Description and Comparison
The carpometacarpus exhibits a large processus intermetacarpalis, which overlaps the os metacarpale minus, and a small cranially protruding process at the proximal end of the sulcus tendinosus (Fig. 1). These derived features are characteristic for passerines; a similar carpometacarpus occurs only in piciform birds (woodpeckers and allies) in which the process at the sulcus tendinosus is, however, situated farther distally and the phalanx proximalis digiti majoris exhibits a derived morphology (Mayr et al. 2003, Manegold 2005; see also Ballmann 1969 for other differences between the carpometacarpi of passeriform and piciform birds). There is no processus dentiformis on the cranial margin of the bone, which is present in Acanthisittidae and very prominent in many oscines and suboscines (Pocock 1966, Mourer-Chauviré et al. 1989, Manegold 2005). As in extant suboscines, the distal end of the fossil's os metacarpale minus is only moderately protruding and its cranial portion forms a projection that reaches farther distally than the facies articularis digitalis minor (Fig. 1; Mourer-Chauviré et al. 1989, Mayr and Manegold 2006:fig. 3). In extant suboscines, this projection fits into a recess on the proximal end of the cleaver-shaped phalanx proximalis digiti majoris (Manegold 2005). In contrast, the distal end of the os metacarpale minus of oscines is not only much more protruding than in extant suboscines and SMF Av 504, but further forms a marked square projection very unlike the condition found in the fossil (Fig. 1). In Acanthisittidae, the os metacarpale minus is also more strongly protruding but, as in suboscines and SMF Av 504, its cranial portion forms a projection that is especially marked in some fossil species (Millener and Worthy 1991:fig. 12). In the somewhat larger early Oligocene passerine described by Mayr and Manegold (2006), the distal end of the os metacarpale minus is either still hidden in the matrix or did not protrude distally.
Whereas the carpometacarpus is seen in dorsal view, the detached distal wing elements expose their ventral surfaces. As in other passerines, the phalanx digiti alulae is short. The phalanx proximalis digiti majoris differs from the oscine crown group and agrees with suboscines in that it is cleaver-shaped and considerably widens cranio-caudally toward its distal end (this phalanx is of roughly equal width in Acanthisittidae and oscines; Fig. 1). There is a marked furrow along the cranial margin of its ventral surface, cranial to the fossa ventralis. The phalanx distalis digiti majoris is fairly long and measures almost the length of the phalanx proximalis. It bears a distinct caudally protruding tubercle on the caudal margin of its proximal end, as in extant Cotingidae (Naked-throated Bellbird [Procnias nudicollis], Andean Cock-of-the-Rock [Rupicola peruviana], Pompadour-Cotinga [Xipholena punicea], and Red-ruffed Fruitcrow [Pyroderus scutatus]). The phalanx digiti minoris is slender. On first sight, the os carpi ulnare appears to be cranio-caudally wider than the corresponding ossicle of the crown group Passeriformes (Fig. 1), but we assume that this impression is because there is a sesamoid ossicle attached to its caudal surface, as, for example, in pittas (suboscines, Pittidae), the Superb Lyrebird (Menura novaehollandiae, oscines, Menuridae; Fig. 1), and many other passerines. In the fossil, presence of this sesamoid bone, which to the best of our knowledge has not been previously mentioned in the literature, appears to be indicated by a very fine fissure.
Discussion
The fossil specimen described in the present study is from a species of similar size to the Malagasy sunbird-asities (Neodrepanis spp.), the smallest extant Old World suboscines. The specimen exhibits a projection of the os metacarpale minus which fits into a recess of the cleaver-shaped phalanx proximalis digiti majoris, a character complex considered to be an apomorphy of suboscines (Manegold 2005). If future, more complete specimens support its assignment to suboscines, it would be the earliest fossil record of this passeriform taxon. Owing to its fragmentary preservation, assignment to a particular taxon within the suboscines is not possible.
Only two fossil records of suboscines have previously been reported from the Old World: specimens of the Eurylaimidae from the early Miocene of Germany (Ballmann 1969) and an undetermined, and as yet undescribed, suboscine from the late Oligocene of France (Mourer-Chauviré 1995). Cheneval (2000) assigned a distal tarsometatarsus from the early Miocene of France to the Dendrocolaptidae (woodcreepers), a taxon now restricted to South America, but we do not consider this assignment to be justified and even doubt its identification as a passeriform bird. There is no unambiguous pre-Pleistocene record of suboscines in the New World; although Noriega and Chiappe (1993) tentatively identified a fragmentary distal humerus from the early Miocene of Argentina as being from a suboscine passerine, the specimen is so fragmentary that it cannot be reliably assigned to any passeriform subclade.
To date, all European passerines described from the early Oligocene have been nonoscine taxa. Since the earliest oscine Passeriformes occur in the late Oligocene of France (Mourer-Chauviré et al. 1989, Mourer-Chauviré 1996) and Germany (GM and AM, pers. obs.), the fossil record indicates that nonoscine passerines colonized Europe before the arrival of oscines, which originated on the Australian continental plate (Barker et al. 2002).
Acknowledgments
We thank S. Tränkner for taking the photograph, and C. Mourer-Chauviré and an anonymous referee for comments on the manuscript. This study is part of a project supported by Deutsche Forschungsgemeinschaft (MA 2328/3-1).
Literature Cited

{kind=link}