





Abstract
Blue-black Grassquits (Volatinia jacarina) are small, granivorous, Neotropical birds that are abundant in central Brazil. During the reproductive season, the socially monogamous males acquire a blue-black plumage and defend very small, clustered territories that resemble leks. They execute a conspicuous courtship display that consists of a leap, revealing white under-wing patches, synchronized with a vocalization. We collected data on the morphology and behavior of banded males and characteristics of their territories to determine how these factors may influence acquisition of mates and nesting. For a second group of birds in the area, we used microsatellite genotyping to test the breeding synchrony hypothesis, which predicts that tropical species that breed synchronously should exhibit high rates of extra-pair fertilization (EPF). We found that males that successfully formed a pair bond differed from unsuccessful males in their behavior, but not in morphological attributes or territory features. Successful males spent more time in their territories, executed displays for longer periods and at greater rates, and their display leaps were higher. These results point to the greater importance of behavior relative to other factors in the successful reproduction of Blue-black Grassquit males. In the second group of birds, EPFs occurred in 63% of 11 nests and involved 50% of the 20 chicks sampled. This exceptionally high incidence of EPF in the small sample analyzed occurred in diverse contexts, including intraspecific parasitism and quasi-parasitism, a rare type of maternity loss where the resident female is parasitized by other females that were fertilized by the resident male. A short and highly synchronized breeding season, clustered breeding in small territories, and granivorous habits may contribute to the high rates of EPF in this tropical species.
Resumen
Estratégias Reprodutivas De Um Pássaro Neotropical Socialmente Monogâmico: Fertilizações Extra-par, Comportamento E Morfologia
O Tiziu (Volatinia jacarina) é um pequeno pássaro granívoro neotropical muito abundante na região central do Brasil. Durante a estação reprodutiva os machos socialmente monogâmicos adquirem uma plumagem negro-azulada e defendem pequenos territórios agrupados, em um padrão que lembra o sistema reprodutivo do tipo lek. Tais machos realizam ainda uma exibição comportamental bastante conspícua, que consiste de pequenos saltos seguidos de uma breve exposição das manchas brancas sob as asas juntamente com uma vocalização. De modo a estudar aspectos relacionados à aquisição de parceiros reprodutivos e nidificação, nós coletamos dados sobre a morfologia e comportamento de machos anilhados, assim como as características de seus territórios. Utilizamos, ainda, marcadores moleculares microsatélites para testar a hipótese da sincronia reprodutiva, que prevê altas taxas de cópula extra-par (CEP) em espécies tropicais que se reproduzem sincronicamente. Os resultados mostram que machos que se reproduziram localmente diferiram dos que não se reproduziram com relação às suas características comportamentais, mas não com relação aos diferentes aspectos de sua morfologia e características territoriais. Machos bem sucedidos reprodutivamente passaram mais tempo dentro de seus territórios, executaram suas exibições durante maiores períodos de tempo e, ainda, apresentaram taxas de execução e altura dos saltos mais elevadas. As CEP ocorreram em 63% dos 11 ninhos amostrados e envolveram 50% dos 20 filhotes encontrados. Esta elevada taxa de CEP ocorreu de diversos modos, incluindo o parasitismo intra-específico e o quasi-parasitismo, um tipo raro de perda de maternidade, onde a fêmea residente é parasitada por outras fêmeas que foram fertilizadas pelo macho residente. Estes resultados mostram a grande importância do comportamento na reprodução do tiziu e indicam que a curta e sincronizada estação reprodutiva, aliada à reprodução em pequenos territórios agrupados, além dos hábitos granívoros da espécie, podem contribuir para as altas taxas de CEP.
Introduction
Variation in reproductive success has been shown to lead to the selection of male characteristics (Andersson 1994), thus competition for reproductive partners is expected to occur (Kirkpatrick et al. 1990, Andersson 1994). In monogamous species, such competition may be a consequence of distortion in the sex ratio of the population, leading to the exclusion of some individuals from reproduction, and of differences in quality among individuals (Andersson 1994). To increase her reproductive success, a female may choose a male based on his morphological or behavioral characteristics, which may be used as indicators of physical condition, sexual maturity, or potential for paternal investment. Alternatively, a male may be chosen on the basis of characteristics of his territory, which may be related to greater availability of good nesting or foraging sites, and protection against predators (Birkhead and Møller 1992).
Monogamy is the predominant mating system of birds, occurring in 91% of species (Lack 1968). However, in some cases social pairing may not signify an exclusive reproductive relationship. Extra-pair paternity (EPP) is widespread, and has been found in at least 130 of the approximately 150 birds studied with genetic markers (Griffith et al. 2002). Among socially monogamous species, over 11% of offspring, on average, result from EPP (Griffith et al. 2002). The significance of this differs for the sexes. For males, EPP offers a way to increase reproductive success without the associated costs of paternal investment. Numerous hypothesized advantages of extra-pair copulation (EPC) have been proposed for females (Birkhead and Møller 1992, Wagner 1992, 1993, Griffith et al. 2002).
Over 90% of all parentage studies have been conducted on temperate, socially monogamous birds (Stutchbury and Morton 2001). Thus, generalizations concerning rates of extra-pair fertilization (EPF) for birds are likely to be biased, since the highest diversity of birds occurs in the tropics. The very few parentage studies on socially monogamous tropical birds have indicated few (<15%) or no extra-pair young, and Stutchbury and Morton (2001) proposed the breeding synchrony hypothesis to account for such low rates of EPF in tropical birds. The relatively short breeding season in temperate zones leads to highly synchronized breeding, and high availability of fertile females (13%–72%; Stutchbury 1998), resulting in high rates of EPF. In the tropics, females in the same population breed over extended periods, and as few as 8% may be fertile at a given time, resulting in low rates of EPF.
The Blue-black Grassquit (Volatinia jacarina) is a small, granivorous passerine that occurs from southern Mexico to northern Argentina and Chile, and is very abundant throughout Brazil (Sick 1997). The species inhabits open vegetation, such as savanna grasslands or areas of disturbed vegetation. Males exhibit a blue-black breeding plumage, with white underwing patches. Females and juveniles are brownish, with a white, streaked breast (Alderton 1963, Sick 1997, Doucet 2002).
During the breeding season (November–April) in central Brazil, males are found in aggregations, within which each individual defends a very small territory. Males exhibit a conspicuous display, composed of repeated vertical leaps from elevated perches, accompanied by a short vocalization and a rapid opening of the wings at the top of the leap that exposes the white underwing patches (Weathers 1986, Sick 1997). The small, aggregated territories and conspicuous displays resemble a lek breeding system. However, past studies have indicated that females nest within male territories, and that males invest in parental care (Alderton 1963, Marcondes-Machado 1988, Almeida and Macedo 2001). These characteristics are incompatible with the usual definition of a lekking system (Bradbury 1981), and social monogamy has been suggested as the mating system of the species (Almeida and Macedo 2001).
We conducted two parallel investigations within a broader study concerning the reproductive biology of the Blue-black Grassquit in central Brazil. Because of the lek-like characteristics of the clustered territories, it is possible that males vary greatly in morphological and behavioral attributes, and that female choice is actively exercised. Thus, the objective of the first part of the study was to evaluate which parameters have a greater impact on immediate measures of male fitness, such as successfully acquiring a breeding territory: behavioral, morphological, or territory features? To answer this, we asked whether males that successfully obtained territories and attracted a mate differed in the above parameters when compared to those that were unsuccessful, despite defending territories within the same area. In the second part of the study, we used microsatellite genotyping to analyze the genetic basis of the breeding system and test the prediction that tropical birds that breed synchronously have high rates of EPF (Stutchbury and Morton 2001).
Methods
Study Area
The study area is within the Fazenda Água Limpa (15°57′S, 47°56′W), a 2000 ha property of the Universidade de Brasília, approximately 18 km southeast of Brasília, Brazil, and composed mostly of native cerrado vegetation with some agricultural areas. The region has a marked seasonal climate, with a rainy season from October to March. For the parentage analyses, we collected data from November to March, 1999–2000 and 2000–2001, in two different areas within the Fazenda. The site used in 1999–2000 was an agricultural area, and encompassed 0.3 ha. In the second breeding season we used an area of about 0.4 ha composed of native cerrado sensu strictu, partially invaded by exotic grasses. We used this latter site to collect data on male breeding parameters from December 2000 to March 2001. Thus, the genetic and breeding biology data reported in this paper, though collected in the same area, refer to different individuals in most cases, and only a very small number of males were used in both parts of the study.
Field Methods
The following methodologies were common to both parts of the study. We captured individuals in the study areas with mist nets (2.5 m × 12 m, 10 mm mesh), and banded them with numbered metal bands (Brazilian bird-banding institution, CEMAVE) and unique combinations of three colored plastic bands (A. C. Hughes Ltd., Middlesex, UK). Individuals were weighed to the nearest 0.2 g with a Pesola scale, and measurements of the tarsus, wings, tail, exposed culmen, bill height, and bill width at the base were taken to the nearest 0.1 mm with digital calipers. We assumed the triangular area of the white underwing patch approached a right triangle, and estimated its size by measuring two adjacent sides of the patch and geometrically calculating the resulting area.
Male Behavior and Territory Measurements
We demarcated male territories by marking the places where aggressive interactions between neighboring males occurred and all perches used during displays, and used these points to define a maximum-sized polygon. We measured the sides of the polygon and the angles between them to geometrically calculate the area of each territory. To evaluate the vegetation structure of territories, we stretched a 50 m measuring tape across the two longest distances inside the territory, and marked out segments corresponding to different vegetation categories along the tape. Categories were: (1) exposed ground; (2) herb stratum; (3) low grass (≤40 cm); (4) high grass (>40 cm); and (5) mixed grass with shrubs. The percentage occupied by each category along the two measuring tapes was extrapolated for the territory. We also counted and measured the height of all possible perches (shrubs and trees) in each territory. Each territory was searched for nests throughout the season. We were able to distinguish three subgroups of males within the study site: (1) males that reproduced locally and had nests in their territories; (2) males that maintained territories in the study site but never had mates or nests; and (3) males that executed displays within the study area but were unable to maintain territories or nest there, and eventually left the area.
To characterize male activities, we observed each focal male for a minimum of 30 consecutive minutes every 15 days throughout the season. Observations were conducted between 07:00 and 12:00 (DST) to minimize variation due to time of day. Breeding activities appear to be tightly synchronized in this population because of the seasonal nature of the climate in this region of Brazil, although a formal measure (sensu Kempenaers 1993) of synchrony was not conducted. Thus, male activities probably varied due to individual differences and not differences in breeding stage. We registered the type of activity exhibited by males at 1-min intervals (totaling 30 observations in the half-hour periods) in the following categories: (1) absence from the territory; (2) on the ground below vegetation; (3) in active display; (4) perched between displays; (5) confronting another individual (chases, conflicts); and (6) performing parental activities (nest-building, feeding chicks or female, perching very close to the nest). Display activities were also considered in more detail, as described below. During these observations of males, we also quantified the time spent by females within a male territory, whether or not there was a nest present. Thus, each observation of male behavior was accompanied by a record of the presence or absence of one or more females within the territory of that male.
Two previous studies conducted with Blue-black Grassquits showed, first, that seasonal rates of display intensity for the whole population apparently do not vary and, second, that males do not change their individual display patterns over the course of the season (Almeida and Macedo 2001, Costa and Macedo 2005). Thus, the behavioral observations of each male included evaluations of his display. Each male was observed for five consecutive minutes to quantify: (1) display rate (i.e., number of displays per minute); and (2) height of individual leaps and of the perches used for displays. From a distance we estimated leap and perch heights through a translucent scale, and subsequently measured the real perch height with a tape measure. The same observer made all measures of leap height, and the translucent scale was always held at eye level and at arm's length. Using the estimates of leap height and perch height made through the translucent scale and the measured perch height, it was possible to arithmetically calculate an approximation of the actual leap height.
Genetic Parentage Data
We collected blood samples (about 50 µl) from the jugular vein of captured individuals, and stored samples in 3 ml of lysis buffer (Quinn et al. 1999) at 4°C until DNA extraction. We monitored banded individuals during each breeding season to determine mated pairs and to identify potential fathers of broods. We took approximately 20 µl of blood from the jugular vein of nestlings six or more days after hatching.
Dna Extraction and Development of Microsatellites
Approximately 30 µl of each sample (blood plus buffer) was incubated overnight with proteinase K at 55°C, and genomic DNA was obtained by phenol-chloroform extraction. Enrichment was done by hybridization subtraction (Edwards et al. 1996) as modified by Muniz et al. (2003). One µg of genomic DNA from each of nine unrelated individuals (four males and five females) was pooled and restricted with MboI (Promega, Madison, Wisconsin), 1× Buffer C, and 100 µg ml−1 BSA. The digested DNA was ligated to 500 ng of annealed SauLA and SauLB linkers (Armour et al. 1994) and 2 µl of the ligate was PCR amplified following the methods of Muniz et al. (2003). Enrichment was done with (CA)15 oligonucleotides. These were cross-linked to nylon membranes (Hybond-N+, Amersham, Little Chalfont, UK) and prehybridized; the amplified ligate was then hybridized to them and nonspecific hybridization products were removed. Bound DNA was eluted, precipitated, and subjected to a second round of PCR amplification. Amplification was followed by a second round of hybridization subtraction, elution, and precipitation, and 2 µl of this doubly enriched library was then used in a final PCR amplification. The enriched library was prepared for ligation by removal of the Saul linkers with MboI digestion and purification with QuickstepTM PCR purification columns (Edge Biosystems, Gaithersburg, Maryland) following the manufacturer's protocol. Purified, enriched library fragments were ligated into ‘Ready-to-go’ BamHI digested pUC18 vector and ligated plasmids were transformed and screened with end-labeled (CA)24 oligonucleotides following Muniz et al (2003).
We isolated and amplified 49 strongly positive clones with M13 primers to determine insert length. Twelve clones had inserts of 300–600 bases and were sequenced on an ABI 377 sequencer (GMI Inc., Ramsey, Minnesota). Of these, 11 had simple or broken repeats of more than 10 base pairs and sufficient flanking region for primers designed using Primer3 (Rozen and Skaletsky 1998). Only two (VjE5 and VjJ13) were highly polymorphic and were used in the analysis of parentage. Six other loci, originally developed for the closely related Medium Ground-Finch (Geospiza fortis, Petren 1998), were also optimized for the Blue-black Grassquit. Microsatellite sequences for the Blue-black Grassquit are deposited in GenBank® (accession numbers: AY730043, AY730045, and AY939817–AY939823).
Population Genetic Diversity and Parentage Analyses
Allele frequencies and Hardy-Weinberg equilibrium probabilities using the Markov chain method were calculated with GENEPOP version 3.2a (Raymond and Rousset 1995). We analyzed parentage with CERVUS version 2.0 (Marshall et al. 1998). Since neither parent was known with certainty, we followed the recommendations in the CERVUS user's manual (Marshall 2001) and first examined the sex with fewer parental candidates (females). We then used the assignments for females that were at the 90% level or greater to run the analysis for the males.
Statistical Analyses
Data were analyzed using SYSTAT 5.0 for Windows (SYSTAT Software 1992). Two-tailed Student's t-tests were used to assess differences in morphological characters between males that were and were not reproductively successful. We conducted Mann-Whitney U-tests to evaluate whether males with and without active nests in their territories varied in terms of time invested in different activities, sizes of their territories, number and height of display perches, and in vegetation characteristics of their territories. We used a Mann-Whitney U-test to compare behavioral measures between the two groups of males, and an ANOVA to examine these parameters among males within the group of successfully reproducing individuals. Finally, ANOVAS were used to evaluate variation in display behavior between the two groups of males and among males in the successful category. Data are presented as means ± SD.
Results
Territoriality and Nesting
Because there were no statistically significant differences in measurements of the left and right wings and white underwing patches of individuals, we used only the measurements of the right wings. We did not find any statistically significant differences in morphological measures between males that nested locally (n = 9) and those that were excluded from nesting in the area (n = 37), i.e., those that either left the area or had territories in the area but no nests (two-tailed t-test: all t44 < 1.5, all P > 0.05). However, individuals that were excluded from nesting in the study site may have nested elsewhere after leaving the area.
We demarcated 15 territories: 14 occupied by banded males and one by an unbanded male that we were unable to capture. Nesting occurred in 12 (80%) of these territories. We found a single nest in eight territories, and two nests in four territories, only one of which in each territory was active. The mean area of these very small territories was 72.4 ± 25.5 m2 (n = 15) with an average of 13.6 ± 10.2 possible display perches. We found no difference in the size of territories of males that obtained mates and nested (n = 12) and those of males that did not have nests (n = 3; Mann-Whitney U = 16, P = 0.77). We also found no difference in the number or average height of potential display perches of males in these two categories (U = 16, P = 0.83; U = 12, P = 0.39, respectively). None of the categories of vegetation structure measured differed between territories of males that did and did not nest in the study area (Mann-Whitney U-tests: all U < 25.5, all P > 0.05).
Male Behaviors and Presence of Females
To gauge differences in behavior, we compared the percentage of time spent on different types of activities by males that reproduced and males that occupied a territory but were unable to obtain a mate. We did not analyze behaviors exhibited after nesting occurred, since these parental behaviors were exclusive to only one class of males. This analysis indicated that males that reproduced spent more time in their territories and displayed for longer periods than males that held territories but did not obtain a mate (Table 1). However, there was no significant difference in the percentage of time spent by females in the territories of males that nested successfully (22.4 ± 24.4 min, n = 7 bouts; each bout refers to an observation period of 30 min conducted every 15 days), and in those of nonreproductive males (8.5 ± 6.7 min, n = 11 bouts), despite the almost three-fold difference in means (U = 55, P = 0.13).
Average percentage of time (min) spent on different activities by territorial Blue-black Grassquit males that nested in central Brazil and those that were unsuccessful in nesting, during 30-min observation bouts conducted every 15 days. Seven observation bouts were conducted for reproductive males and 11 for nonreproductive males. Absence means the male was absent from his territory, Ground means the male was on the ground below vegetation, and Confrontation means the male was chasing or fighting with another individual. The U and associated P-values reflect Mann-Whitney U-tests
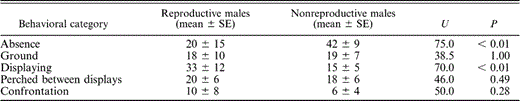
Average percentage of time (min) spent on different activities by territorial Blue-black Grassquit males that nested in central Brazil and those that were unsuccessful in nesting, during 30-min observation bouts conducted every 15 days. Seven observation bouts were conducted for reproductive males and 11 for nonreproductive males. Absence means the male was absent from his territory, Ground means the male was on the ground below vegetation, and Confrontation means the male was chasing or fighting with another individual. The U and associated P-values reflect Mann-Whitney U-tests
We also analyzed the behavior (before and after egg-laying pooled) of males that nested. Males differed in the percentage of time spent displaying (F11,28 = 4.3, P < 0.01) and on the ground (F11,28 = 2.7, P < 0.02), but not in other activities (all F11,28 < 0.9, all P > 0.50). The time spent by females in individual territories before and after mating was not significantly different (F11,28 = 1.38, P > 0.20).
Displays
Display characteristics (height of the leap and display rate) were compared between males that obtained a mate and those that did not. Again, we only included activities performed before nesting. Males that reproduced executed significantly higher display leaps (31.3 ± 9.5 cm, n = 72 leaps) than males that did not mate (20.9 ± 7.3 cm, n = 55 leaps; F1,125 = 45.8, P < 0.001; Fig. 1A). They also displayed at significantly higher rates than males that did not acquire mates (reproductive males: 13.3 ± 2.2 leaps per min, n = 10 bouts; nonreproductive males: 9.7 ± 3.0 leaps per min, n = 10 bouts; F1,18 = 9.2, P < 0.01; Fig. 1B). Analyses of reproductively successful males only showed that individuals differed with respect to the height of display leaps (F11,222 = 14.5, P < 0.001), but not display rate (F11,26 = 1.9, P > 0.05).
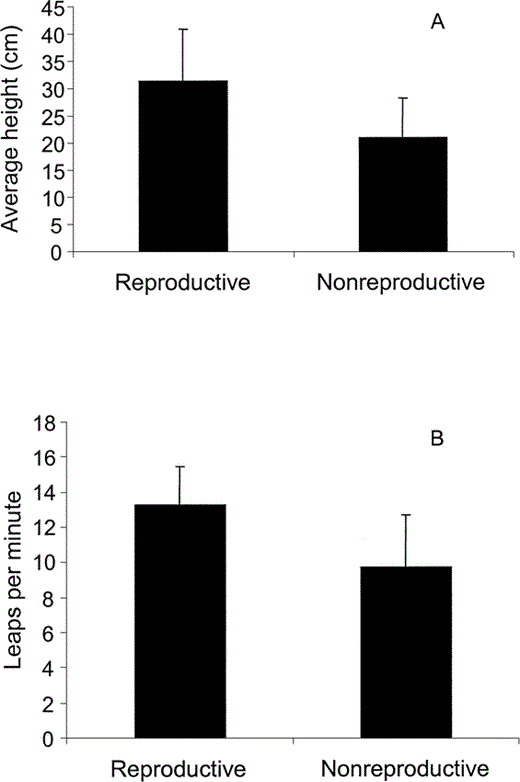
Figure 1. Characteristics of courtship displays of successfully reproducing Blue-black Grassquit males (reproductive) and males that did not have nests (nonreproductive) in Fazenda Água Limpa, Brazil: (A) mean height (± SE) of display leaps, and (B) mean display frequency (± SE)
Parentage
Potential parents (31 adults, social parents or seen in the vicinity of nests) were confirmed with a reasonable degree of certainty for 20 chicks from 11 different nests. Blood samples from both adults attending the nest were available for five of these nests (11 chicks). For the other six nests (9 chicks), only blood from the adult male attending the nest was available.
From the 51 individuals genotyped, there were 7–18 alleles per locus with heterozygosity ranging from 0.196–0.922 (Table 2). Gf01 and Gf05 were in genetic disequilibrium even after correcting for multiple comparisons (P < 0.001). Two loci were significantly out of Hardy-Weinberg equilibrium (Gf05 and VjE5; P < 0.001 for both). When using CERVUS it is recommended that any locus with >5% null alleles be dropped from the analysis. For Gf05, the estimate was 14% and for VjE5 it was 7%. The analysis was run both with and without these loci.
Test results of polymorphic microsatellite loci used for testing parentage of Blue-black Grassquits in central Brazil. Ta = annealing temperature; K = number of amplified alleles; n = number of individuals tested; HO = observed heterozygosity; HE = expected heterozygosity; and P = probability of deviation from Hardy-Weinberg equilibrium

Test results of polymorphic microsatellite loci used for testing parentage of Blue-black Grassquits in central Brazil. Ta = annealing temperature; K = number of amplified alleles; n = number of individuals tested; HO = observed heterozygosity; HE = expected heterozygosity; and P = probability of deviation from Hardy-Weinberg equilibrium
Including all eight loci in the analysis, the success rate in paternity evaluation was 100% in cases with a relaxed confidence level (80%) and 79% using a rigorous confidence level (95%). When loci Gf05 and VjE5 were excluded, the assignments of parentage were the same, but with reduced confidence levels; eight of the chicks were attributed to a female with 90% confidence and the other 12 with 70% confidence, while six of the chicks had a male parent identified at the 90% confidence level, and 14 at the 70% level. Only the results of the analysis using six loci are reported here.
Of the 20 chicks genotyped, 10 (50%) from seven nests (64%) were not genetically related to one or both of their social parents (Table 3). All of the genetic mothers were captured and genotyped, but not all of them nested in the study site in both years. We did not capture nine of the social mothers that nested in the study area. In seven cases, paternity could not be attributed to the male that attended the nest. In two other cases, the female providing care at the nest was not the genetic parent of the chick, and in one case the chick was unrelated to either of its social parents. We generated a simple reproductive cost-benefit matrix, using individuals attending nests and chicks in their care (Table 4). There were cases where paternity was attributed solely to the male attending the nest, including cases where different chicks in the same brood were related to more than one female; nests where the whole brood or some nestlings could not be genetically assigned to the male attending the nest; and one case (mentioned above) where one chick was unrelated to either social parent.
Identity of social parents (those providing parental care at the nest) and genetic parents (identified through genetic analysis using six loci) of Blue-black Grassquit nestlings in central Brazil. Letters preceding numbers identify all individuals (C = chick, M = male, F = female). Two asterisks (**) after the individual's identification code indicate cases where social parentage does not coincide with genetic parentage
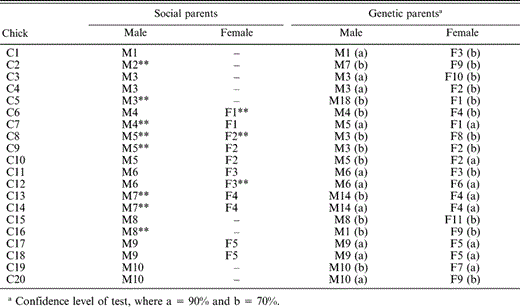
Identity of social parents (those providing parental care at the nest) and genetic parents (identified through genetic analysis using six loci) of Blue-black Grassquit nestlings in central Brazil. Letters preceding numbers identify all individuals (C = chick, M = male, F = female). Two asterisks (**) after the individual's identification code indicate cases where social parentage does not coincide with genetic parentage
Estimates of reproductive success for Blue-black Grassquits attending nests in central Brazil. Letters preceding numbers identify all individuals (C = chick, M = male, F = female). Individuals listed in the columns of within-pair and extra-pair chicks refer to offspring found in that adult's nest. Extra chicks refer to genetic offspring of that adult found in other nests
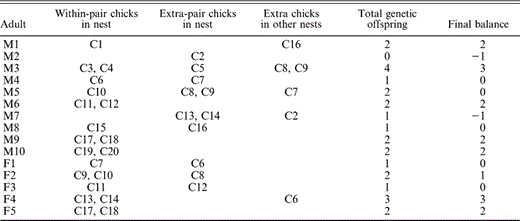
Estimates of reproductive success for Blue-black Grassquits attending nests in central Brazil. Letters preceding numbers identify all individuals (C = chick, M = male, F = female). Individuals listed in the columns of within-pair and extra-pair chicks refer to offspring found in that adult's nest. Extra chicks refer to genetic offspring of that adult found in other nests
Discussion
We did not find morphological variation between Blue-black Grassquit males that successfully nested and those that did not reproduce, which implies that morphology may not be a critical factor in the sexual selection process for this species. However, we cannot exclude the possibility that other morphological traits that we did not evaluate may be important (e.g., structural color of plumage; Doucet 2002). In addition, the absence of size, composition, and structural vegetation differences among occupied male territories suggests that females may not choose partners based on territory characteristics, confirming the findings of Almeida and Macedo (2001). However, it is important to note that because we had small sample sizes, the failure to find differences among features of territories and morphological traits of reproductive versus nonreproductive males may be due to low statistical power.
The displays of male Blue-black Grassquits result in greater conspicuousness, both to possible rivals and to females (Weathers 1986, Wilczynski et al. 1989). We found that males that spent more time displaying and within their territories were more successful in reproducing, and these may be characteristics favored by females. Likewise, female Bobolinks (Delichonyx oryzivorus) preferred males that spent more time in aerial displays (Mather and Robertson 1991). The difference that we found in the rate and height of display leaps between males that were successful in nesting and those that were not, as well as among males that nested, indicate that the traits that compose displays are also critical. Because males continued displaying during nesting, displays may have the double function of signaling male presence while mate-guarding, and also indicating the male's physical condition or status to other females that may be searching for high-quality males.
The extra-pair fertilization (EPF) rate of 50% found for the Blue-black Grassquit is much higher than the average of 11% found in socially monogamous birds (Griffith et al. 2002), and only three temperate species studied to date have comparable levels of EPF: the Red-eyed Vireo (Vireo olivaceus, Morton et al. 1998), Tree Swallow (Tachycineta bicolor, Barber et al. 1996, Dunn et al. 1994), and Reed Bunting (Emberiza schoeniclus, Dixon et al. 1994). Some authors (Stutchbury and Morton 1995, Stutchbury et al. 1998, Petren et al. 1999) have suggested that high rates of EPF should not be common in tropical species because of their reproductive asynchrony, which would thwart the possibility of females comparing males for mating, and decrease the possibility of extra-pair copulation (EPC). However, tropical species that breed synchronously would be expected to exhibit high rates of EPF. In central Brazil the rainy season is extremely seasonal, thus birds may breed relatively synchronously. Because the Blue-black Grassquit is granivorous, and seeds are only abundant toward the end of the rainy season, breeding is restricted to an even shorter time period. In 35% of cases, the paternity of chicks could not be attributed to the male attending the nest. It is hypothesized that females that seek EPCs may specifically select males of better quality than their social mates (Kempenaers et al. 1997). Benefits obtained by females through EPC may include the acquisition of “good genes” or genes associated with attraction (Houtman 1992, Hasselquist et al. 1996, Kempenaers et al. 1997), which would increase the fitness of the female's brood. In our study, males differed in behavior and display rates, and a recent study (Costa and Macedo 2005) revealed that display rate, wing area, and body mass are negatively associated with levels of infestation by coccidian parasites in Blue-black Grassquits. Thus, female mate choice in this species could be based on behavioral differences among males that indicate superior genetic quality, health conditions, or elevated status.
An alternative hypothesis is that the aggregated territories and possible reproductive synchrony in Blue-black Grassquits foster conditions conducive to EPCs in the absence of any male or female reproductive strategies. High levels of reproductive hormones may be implicated in copulatory behavior directed toward neighbors, whether or not they are of better quality than social mates. Further work is needed to determine whether extra-pair partners are chosen selectively or indiscriminately to differentiate between these two hypotheses.
In this study, we found two chicks in different nests that were not genetically related to the female providing parental care, but were related to the male associated with the nest. This type of nest parasitism is dubbed quasi-parasitism and is quite rare (Birkhead et al. 1990, Petter et al. 1990, Petrie and Møller 1991, Alves and Bryant 1998). We also detected one case in which the chick was not related to either social parent, which may be a case of intraspecific parasitism. In addition to being a possible female strategy for increasing reproductive success without the cost of parental investment, nest parasitism, when found in only a small proportion of nests, may result from opportunistic laying due to other reasons. Whether or not intraspecific parasitism or quasi-parasitism are common in the Blue-black Grassquit needs to be confirmed with larger samples.
The lack of mate fidelity in this species is not surprising given male aggregations and their conspicuous displays. The “hidden lek hypothesis” proposes an explanation for the evolution of territorial aggregation of males in monogamous species (Wagner 1997). This hypothesis suggests that territorial aggregation occurs around males of superior genetic quality as a consequence of female preference for territories closest to these males, to increase chances of obtaining EPCs from that male. Our study was not designed to test the “hidden lek hypothesis” and our limited genetic sampling precludes conclusions on this point. Nonetheless, our results are intriguing and future studies could test this hypothesis. In the genetic component of the study, six of ten males were socially and genetically monogamous. However, of these, only three were able to avoid rearing extra-pair chicks. Some males suffered a reproductive loss even when siring extra-pair chicks, while other males that produced extra-pair chicks also cared for the chicks of other males. The average values of the final reproductive balance for different males indicated that the most successful male reared an extra-pair chick in his own nest, in addition to two of his own, and sired an additional two chicks that were cared for by other males. On average, however, the highest reproductive success was obtained by males whose behavior resulted in no paternity loss, whether or not they were able to produce chicks through EPCs. It is possible that all males attempt to obtain EPCs, and only some are successful. In addition, though males may attempt to defend their mates and territories, they may be unsuccessful because of the close proximity of reproducing neighbors.
In conclusion, behavioral characteristics such as occupation of the territory and persistence in displaying, in addition to higher and more frequent display leaps, appear to be important attributes leading to male acquisition of mates. Despite the small sample for analysis of parentage, we found one of the highest incidences of EPF among birds reported to date, indicating that Blue-black Grassquits lack genetic monogamy despite being socially monogamous. Furthermore, our results indicate that high rates of EPF do occur in some tropical birds. In addition to the highly synchronous breeding that may be a prerequisite for high rates of EPF in tropical birds (Stutchbury and Morton 2001), we suggest that granivory in areas of high seasonality will lead to rates of EPF comparable to temperate zone species.
Acknowledgments
We thank Verônica Borges, Laura Muniz, Fabio Costa, and Valéria Saracura for their field assistance, and we are grateful for the help provided by Laura Muniz and Brian Powell in the laboratory. Helpful reviews of the original manuscript were provided by Maria Alice Alves, Mariana Cariello, Guarino Colli, Miguel Marini, Helena Morais, Richard Wagner, and two anonymous referees. Research was funded by the Conselho Nacional de Desenvolvimento Científico e Tecnológico through a graduate scholarship awarded to CBVC, and a fellowship for RHM. We thank the Center for the Study of Tropical Birds (Clark Franzier Memorial Grant) for providing research funds, and Universidade de Brasília and the University of St. Andrews for logistic support.
Literature Cited
Author notes
Present address: Conselho Nacional de Desenvolvimento Científico e Tecnológico, SEPN 509 Bloco A, Sala 304, 70750-501 Brasília, D.F. Brazil. [email protected]

{kind=link}