





Abstract
While male song and its functions have been well studied, female song has often been overlooked. In this study, we provide a detailed description of the spontaneous female song in a well-studied northern temperate songbird, the European Starling (Sturnus vulgaris). We compared the song organization, complexity, and composition of female and male starlings housed in large outdoor aviaries. Overall, the general organization was similar in both sexes, and some females sang complex song bouts of more than 30 seconds of uninterrupted song. Although some females were capable of singing the four phrase-type categories typically found in male song bouts (whistle, variable, rattle, and high-frequency phrase types), a significantly lower proportion of females sang all four categories of phrase type as compared to males. Our results also revealed large individual variation in song characteristics among females: repertoire size varied between 11 and 36 phrase types, while average song bout length ranged between 10 and 26 seconds. All song parameters (total repertoire size, song bout length, and repertoire size within the four different phrase categories) were significantly lower in females than in males. Nevertheless, except for the number of rattle phrase types, song parameters overlapped between the sexes demonstrating that some females produced a more complex song than some males.
Canto de la Hembra en Sturnus vulgaris: Diferencias entre Sexos, Complejidad y Composición
Resumen. Mientras que el canto del macho y sus funciones han sido bien estudiados, el canto de las hembras ha sido menos estudiado. En este estudio, brindamos una descripción detallada del canto espontáneo de la hembra en una especie bien estudiada de ave canora del norte templado, Sturnus vulgaris. Comparamos la organización del canto, la complejidad y la composición entre machos y hembras en aviarios de gran tamaño emplazados al aire libre. En total, la organización general fue similar en ambos sexos, y algunas hembras cantaron unidades de canto complejas durante más de 30 segundos de modo ininterrumpido. Aunque algunas hembras fueron capaces de cantar las cuatro categorías de tipos de frases típicamente halladas en las unidades de canto de los machos (silbido, variable, matraqueo y tipos de frases de alta frecuencia), una porción significativamente menor de las hembras cantó todas las cuatro categorías de tipos de frases en comparación con los machos. Nuestros resultados también revelaron gran variación individual entre hembras en las características del canto: el tamaño del repertorio varió entre 11 y 36 tipos de frases, mientras que la duración promedio de la unidad de canto fluctuó entre 10 y 26 segundos. Todos los parámetros examinados del canto (tamaño total del repertorio, duración de la unidad de canto y tamaño del repertorio dentro de las cuatro categorías diferentes de frases) fueron significativamente menores en las hembras que en los machos. A pesar de ello, a excepción del número del tipo de frases de matraqueo, los pará metros del canto se superpusieron entre los sexos, implicando que algunas hembras produjeron un canto más complejo que algunos machos.
Introduction
Songbirds' ability to learn and produce complex vocalizations have made them an important model system in communication and sexual selection research (Catchpole and Slater 1995, Slater 2003); however, most of the studies have focused on temperate species in which song is restricted mainly to males (Langmore 1998, Price 1998, Riebel 2003, Slater and Mann 2004). Indeed, the study of song behavior in females has been a long-neglected field (Baptista and Kroodsma 2001). In recent years, however, there is increasing evidence that female song is not so uncommon as previously thought but instead may be an adaptive plastic trait in females (Langmore 1998, Eens and Pinxten 1998).
During the last 20 years, the European Starling (Sturnus vulgaris) has become an important model in bird song and song system research (Eens 1997, Riters and Ball 1999, Ball et al. 2002, Duffy and Ball 2002, Pinxten et al. 2002, Van der Linden et al. 2002, Riters et al. 2005). European Starlings are sociable birds that sing throughout most of the year (Feare 1984), feeding and roosting in flocks, and breeding in colonies. Although it has long been recognized that female starlings often sing (Freitag 1936, 1937, 1939, Witschi and Miller 1938, Bullough 1942, Feare 1984, Eens 1997), the great majority of studies have focused on male starlings. Male starling song has been well described by several researchers (Adret-Hausberger and Jenkins 1988, Eens et al. 1989, 1991a, 1991b, Chaiken et al. 1993, Mountjoy and Lemon 1995). Males produce a long and elaborate song that functions primarily to attract females in spring (Eens et al. 1990, 1993). Males also sing frequently outside the breeding season but the function of song at this time is less clear, although it seems to play a role in flock cohesion or in the maintenance of dominance hierarchies (Eens 1997, Riters et al. 2000, 2002, Pinxten et al. 2002). Males have large repertoires ranging from 14 to almost 70 phrase types (Eens 1997). There is also great deal of evidence both from the field and the laboratory suggesting that individual variation in males' song plays an important role in female mating decisions (Eens et al. 1991a, Eens 1997, Mountjoy and Lemon 1996, Gentner and Hulse 2000). Male starlings produce their song in structured episodes that have been referred to as song bouts. Song bouts vary in length both within and among individuals, but follow a general phonological pattern that appears to be species typical (Eens et al. 1991a, Chaiken et al. 1993, Gentner and Hulse 2000). Complete song bouts consist of four categories of phrase types (whistles, variable phrase types, rattle phrase types, and high-frequency phrase types) that are sung in a highly predictable sequence (Eens 1997).
Female starlings have been reported singing during the breeding and nonbreeding season, and both in the wild and in captivity (Hausberger and Black 1991, Hausberger et al. 1995b, Eens 1997, Henry and Hausberger 2001, De Ridder et al. 2002). The function of female song in starlings is unclear at present, although there are indications that during the breeding season it plays a role in intrasexual competition (Eens et al. 1996, Sandell and Smith 1997). To the best of our knowledge, there are no detailed descriptions of the structure and organization of spontaneous song in female starlings. All previous studies describing song have used testosterone implantation techniques to induce song behavior (Hausberger and Black 1991, Hausberger et al. 1995a, 1995b). Using such techniques, these authors have concluded that testosterone-implanted females produce a song very close in structure to male song. They also reported that the high-frequency song types that males typically sing at the end of a song bout were missing in female starlings. Given that testosterone not only affects the rate but also the structure of song (Fusani et al. 1994, Beani et al. 2000, Ball et al. 2002), it remains to be examined whether these results also hold true for naturally singing females.
Several recent comparative studies have shown that the evolution of sex differences in brain nuclei that control song have coevolved with the evolution of sex differences in singing behavior in songbird species (MacDougall-Shackleton and Ball 1999, Ball et al. 2001). Given that the volume of several song-control nuclei in female starlings are smaller than in males but also that the sexual dimorphism in the song system is much smaller than in species where females do not sing (Bernard et al. 1993), we expected to find a relatively complex song in female starlings. The aim of this study was three-fold. First, we wanted to provide the first detailed and quantitative descriptions of spontaneous song in (captive) female starlings. Second, we describe interindividual variation in song characteristics among females focusing on repertoire size, song bout length, and the presence of each of the four song categories making up song in males. Third, we provide a detailed comparison of song between female and male starlings recorded in similar captive conditions. Our study will hopefully stimulate further research into the causes and consequences of song variation in female starlings and other songbirds, and also provide insight into the function of female song.
Methods
Study Subjects and Song Recordings
We compared the song complexity and song organization between 12 female and 24 male European Starlings. All males and females were at least in their third calendar year when their song was recorded. All song recordings were made in captivity in large outdoor aviaries on the campus of the University of Antwerp, Belgium. Previous studies have shown that male starlings exhibit normal singing behavior in captivity (Eens et al. 1991a, 1991b, Eens 1997). All aviaries were provided with plenty of nest boxes and branches to perch on, and a 30-cm wooden perch was attached to the bottom of each nest box, so that starlings could sit and sing in front of it. Food and water were available ad libitum. All birds were individually marked with a numbered metal ring and color rings.
Given that free-living male and female starlings live in separate flocks throughout most of the year (Feare 1984, Eens 1997), and that males usually arrive in the breeding area before the females (Eens 1997), we decided to observe and record starlings in single-sex groups. We believe that this is a biologically relevant approach, and furthermore such a recording strategy should avoid male behavior (aggression, nest-box occupation, singing) that affects or suppresses female singing behavior.
Twenty-nine adult female starlings were housed together in a large outdoor aviary (12.10 × 8.10 × 2.12 m; length × width × height, respectively) in a single-sex group in June 2000. Spontaneous song of females was recorded between 20 February and 2 April 2001 from a permanent hide situated about 3 m away from the aviary. Recordings were obtained in the morning between 09:00 and 12:00 using clip microphones, (Philips SBC ME 600, Europe), attached inside the nest boxes and connected to a UHER 4400 Report Stereo IC tape recorder (Munich, Germany) via a long cable and recorded on BASF tapes (type DP 26 FE LH, Ludwigshafen, Germany). Although all 29 females were observed singing, and despite the fact that prior to song recording 20 females occupied and defended a nest box, we were only able to obtain a sufficient number of good-quality recordings from 12 females. This was largely due to the fact that often females were singing together in a group, rather than singing close to their nest boxes (i.e., in the close proximity of the microphones).
To compare the song between females and males, we used song recordings from 24 adult captive males, recorded between 1987 and 1994. Most data on male song have already been published elsewhere (see Eens et al. 1991a, 1991b, 1992a, 1992b, Eens 1997). Male starlings were housed in similar conditions as the females, and the methodology applied for recording and analyzing song, was the same as for females.
Male and female starlings were all wild-caught mostly as adults and in a few cases as juveniles, but never as nestlings. Although we do not have information on the birds' developmental history before capture, they were kept in large outdoor aviaries from the moment of capture and they were visually and acoustically exposed to other female and male captive and wild starlings. Therefore, both males and females had access to many different tutors while held in captivity.
Song Organization and Description
The terminology of describing and classifying male starling song varies among researchers. Here, we use the terminology employed by Eens (1997). Starlings produce most of their song in song bouts. A song bout is defined as a period of at least 5 sec of song with pauses no longer than 1.5 sec. A song bout has a complex syntactical and temporal organization and it is composed of complex sequences of discrete units (phrases). A “phrase” is a fixed combination of acoustic elements with a duration of about 0.5– 1.5 sec. Often one phrase is repeated once or several times before the next one is introduced. We refer to one or more successive repetitions of a particular phrase as a “phrase type”. Phrases are organized in four distinct categories of phrase types: whistle, variable, rattle, and high-frequency phrase types. In male starlings, a song bout usually begins with whistle phrase, which are relatively pure tone-like sounds separated by each other with pauses of 1 sec or longer. Whistles are rather simple but they may also contain more structurally complex elements as well as heterospecific imitations. Each male can possess up to 12 whistle phrase types. After the introductory whistles, a song bout progresses into a series of complex phrases (variable phrase types) sung with no pauses or very short pauses between them. Heterospecific imitations are a common component of this part of the song. Males can possess between 10 and 35 variable phrase types in their repertoire. This section of the song is usually followed by phrases containing a rapid succession of clicks (rattles), produced in a low-frequency range (maximum 4 kHz) and delivered at a rate of about 10–15 per second (rattle phrase types). They are softer in volume than the preceding variable phrase types, rarely include heterospecific imitations, and often have no distinct temporal gaps between them. Males can have between 2 and 14 rattle phrase types in their repertoire. Finally, song bouts were usually completed with high-frequency phrase types, the loudest part of the song. High-frequency phrase types are a series of phrases containing high-frequency sounds mainly in the range of 6–10 kHz. Males can possess up to six high-frequency phrase types.
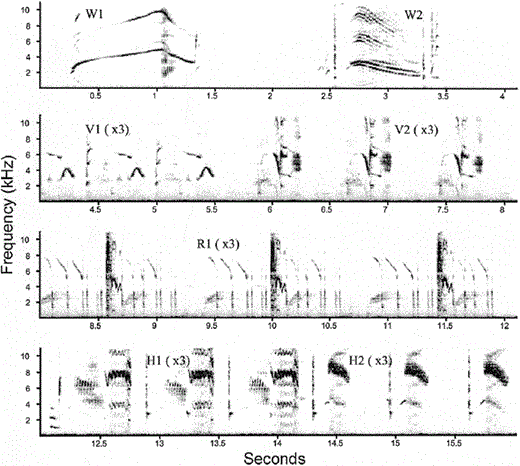
Sonogram of a complete song bout of a female starling (W is a whistle, V is a variable, R is a rattle, and H is a high-frequency phrase type). The illustrated song bout contains 7 different phrase types (two whistle, two variable, one rattle and two high-frequency phrase types) and 17 phrases
A complete song bout consists of all four categories of phrase types that are sung in a highly predictable sequence. However a song bout may end at any point of this sequence and sometimes males may revert to phrase types that have already been sung, thus whistle, variable, or rattle phrase types can be produced after these high-frequency phrase types. Average song bout length varies from 12–40 sec in males, with some song bouts lasting more than 60 sec. Often, phrase types in a song bout contain “double voicing” where two tones are being produced at the same time. Some phrase types, mostly whistles, can be sung apart from the song bout, but in most cases they appear in the song bouts as well. The total number of different phrase types that a bird can produce, is defined as its repertoire size. Overall repertoire size ranges between 14 and 68 phrase types in male starlings (Eens 1997).
To interpret the song bouts from the sonograms, we converted song bouts into a sequence of letters (corresponding to the phrase-type category) and numbers (corresponding to the novel phrase types appearing throughout the song bout; Fig. 1). We determined the repertoire size of each recorded female by counting novel phrase types as they appeared throughout the song bouts. To be certain that the whole repertoire of each female was obtained, we plotted the cumulative number of new phrase types appearing throughout the song bouts against the total number of analyzed phrases (Kroodsma and Miller 1982, Adret-Hausberger and Jenkins 1988, Eens 1997). As can be seen from the cumulative plots (Fig. 2), after a certain number of analyzed phrases, no new phrase types occur; asymptotic curves were reached in all cases.
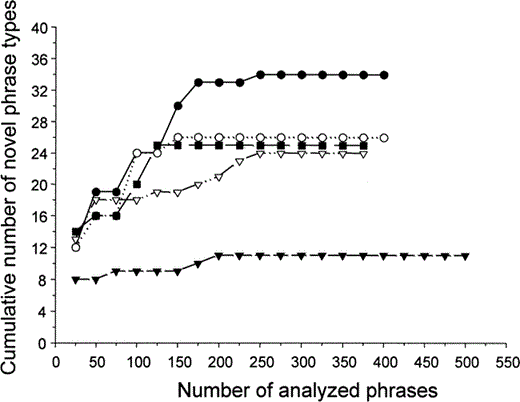
Cumulative plots of the number of new phrase types versus the number of phrases analyzed for five female starlings, corresponding to females 4, 5, 6, 7, and 10 from Tables 1 and 2. The complete repertoire size was obtained only after a minimum of 150 phrase types were analyzed, and in one case after 250 phrase types. The plots represent a subset of females with low, intermediate, and large repertoire sizes
In females, we measured the presence of phrase-type category, the number of phrase types sang per song bout, the duration of the song bout, and the total repertoire size (including repertoire size within each phrase category and phrase types sang apart from the song bout). To compare song characteristics between sexes, in males we measured the presence of phrase-type category, the total repertoire size, and the song bout length.
Statistical Analyses
Data were analyzed using the statistical software program Statistica ver. 5.0 (StatSoft 1995). Song was analyzed using Avisoft SASlab Pro software, version 3.5b (Specht 1993). Data were normally distributed, except in one case, when values had to be log transformed to meet the criteria required for parametric statistics. Relationships among different song parameters were analyzed using Pearson correlations. In one case a two-tailed post hoc power test (GPower 1998) was applied to calculate the statistical power. To test whether song-bout lengths and the number of phrase types sang in a song bout differ among females, a one-way ANOVA was used. To compare other song characteristics between females and males, we applied t-tests for independent samples and Mann-Whitney U-tests. A Fisher Exact test was applied to examine whether females and males differ in the proportion of having all four phrase-type categories in their song bout. The probability level for significance was set at α = 0.05. Values are presented as means ± SE, unless noted otherwise.
Results
Females
As in males, females produced their song in song bouts and each phrase type was repeated one or several times before the next one was introduced. Song bout lengths varied significantly among the females (one-way ANOVA: F11,195 = 6.8, P < 0.001) ranging between 10.2 and 25.9 sec with an overall average of 16.7 ± 1.2 sec (Table 1). There was also a significant difference among females in the number of phrase types sang in a song bout (one-way ANOVA: F11,189 = 7.9, P < 0.001) with a minimum of 5.6 ± 0.3, a maximum of 14.3 ± 1.5 and an overall mean of 8.4 ± 0.6 (Table 1).
Total number of analyzed phrases, aver age number of phrase types sang in a song bout, av erage length of the song bout, and total repertoire size for each female starling. Values are presented as means ± SE. A song bout in European Starlings is defined as a period of at least 5 sec of song with pauses no longer than 1.5 sec (Eens 1997)
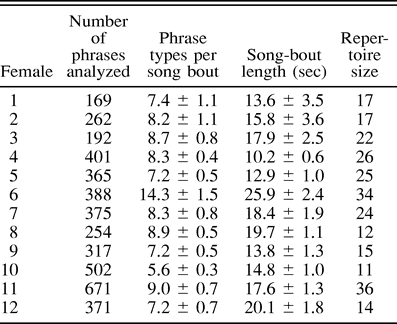
Total number of analyzed phrases, aver age number of phrase types sang in a song bout, av erage length of the song bout, and total repertoire size for each female starling. Values are presented as means ± SE. A song bout in European Starlings is defined as a period of at least 5 sec of song with pauses no longer than 1.5 sec (Eens 1997)
No females sang their entire repertoire in only one song bout. In total, we analyzed 4267 phrases (with a minimum of 169 phrases per female and a maximum of 671 phrases per female; Table 1). Total repertoire size of females ranged between 11 and 36 phrase types (21.1 ± 2.4, Table 1) and was not dependent on the number of phrases that we analyzed (Pearson correlation test: r = 0.48, P = 0.11, n = 12). Repertoire size was significantly positively correlated with the average number of phrase types sang per song bout (Pearson correlation test: r = 0.68, P = 0.01, n = 12). There was no significant relationship between the repertoire size and the average song bout length of a female (r = 0.25, P > 0.05, n = 12), however, the power of the test was 0.3 (effect size conventions: small = 0.1, medium = 0.3, large = 0.5). Further, there was no significant positive relationship between the repertoire size and the number of phrase-type categories that a female was observed to sing (r = 0.47, P = 0.13, n = 12).
Presence of phrase-type categories (whis tle [W], variable [V], rattle [R] and high-frequency [H] phrase types) in the song bouts of female starlings, and repertoire size within each category for each female. A complete song bout in males consists of all four categories of phrase types, while rattle or high-fre quency phrase types could be absent in the song bouts of some females
![Presence of phrase-type categories (whis tle [W], variable [V], rattle [R] and high-frequency [H] phrase types) in the song bouts of female starlings, and repertoire size within each category for each female. A complete song bout in males consists of all four categories of phrase types, while rattle or high-fre quency phrase types could be absent in the song bouts of some females](https://oup.silverchair-cdn.com/oup/backfile/Content_public/Journal/condor/107/3/10.1093_condor_107.3.559/1/m_i0010-5422-107-3-559-t02.gif?Expires=1750241049&Signature=iFgmYf1woX~-vlhTl2g-XvN1NKhc26i7U5UFPdVRsQZHpCqAggMchT5jVPuF95TTCX4zSlPDXSlUrfAlIf5~WR9T1okRMLFFpuhhpEWG3Ih3BORYkgGlgfJIEYWQWMVfec7vgYfwyemG~5BoYbicsl8YuhtIOn2W4YHxP8XA5v7SOSIW-GSuMgz33nGO1D~WxziYL-munXECU9echuODlu6qSRu0dQfUIZ2k6ZI~CzQrYzv49TpxK1lit6Uj81~uAawCKLsuDrbZgTQ9kGm4RM1ie7ZXBgZlsuodE7CbxYieTkXj4p8GnU37USKGic9ShEUvQMaQ-qAG~B-QCUozsw__&Key-Pair-Id=APKAIE5G5CRDK6RD3PGA)
Presence of phrase-type categories (whis tle [W], variable [V], rattle [R] and high-frequency [H] phrase types) in the song bouts of female starlings, and repertoire size within each category for each female. A complete song bout in males consists of all four categories of phrase types, while rattle or high-fre quency phrase types could be absent in the song bouts of some females
In general, female song contained all four categories of phrase types typically presented in male song. However, only 42% of females produced all four categories (Table 2). All females possessed whistle and variable phrase types in their repertoire while rattle and high-frequency phrase types were present in only 58% and 67% of the females, respectively. Most of the song bouts started with a whistle, although whistle phrase types could be sang not only as an initial element, but also at other parts within the song bout. If the song bout did not begin with a whistle, it always started with a variable phrase type; we never observed a song bout starting with a rattle or with a high-frequency phrase type. Rattle phrase types were produced at a rate of 7–16 clicks per second and in a low-frequency range (0.5–4 kHz), similar to those of males (Eens 1997). High-frequency phrase types were sang mostly after the rattle phrase types at the end of the song bout, but sometimes rattle or variable phrase types could be produced after them.
Often song bouts contained heterospecific imitations incorporated in the different phrase categories. Although we did not measure the amount of heterospecific imitations produced per female, all 12 females incorporated such imitations in their song. In Figure 3 we show examples of a heterospecific imitation of a blue tit and of an ambulance (the aviary was situated nearby a hospital). This ambulance imitation was observed in 5 out of 12 (42%) females. “Double voicing” was also frequently observed, occurring in all four categories of phrase types.
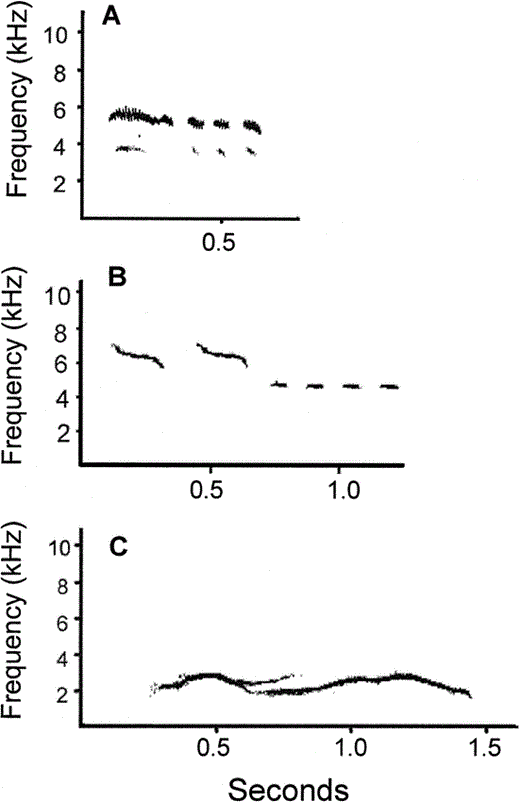
Examples of heterospecific imitations sung by female starlings: A) Blue Tit (Parus caeruleus) imitation by a female starling, B) Sonogram of an actual Blue Tit song, and C) Imitation of an ambulance by a female starling
Comparison Between Females and Males
Twenty-three out of 24 (96%) males produced all four categories of phrase types in their song bouts (high-frequency phrase types were missing in the song of one male), compared to only 5 out of 12 (42%) females (Table 3). This difference was highly significant (Fisher Exact test: P < 0.001). For all song categories except the rattle phrase types, there was an overlap between males and females, indicating that some females produced a more complex song than some males (Table 3). Nevertheless, on average, all song parameters (total repertoire size, song-bout length, and repertoire size within the four phrase-type categories) were significantly higher in males than in females (Table 3). Males sang on average seven times more rattle phrase types than females. The number of phrase types that were sung outside the song bout did not differ between the sexes (Table 3).
Discussion
Here we described in detail the organization of spontaneous song, and demonstrated sex differences in song complexity and song composition in starlings. In many previous studies exploring female song in passerines, song was induced by testosterone administration (Kriner and Schwabl 1991, Hausberger and Black 1991, Hausberger et al. 1995a, Vallet et al. 1996), thus data on spontaneous song in female birds are often incomplete. Although the spontaneous song of female starlings has been previously investigated (Hausberger et al. 1995b, Henry and Hausberger 2001), until now it has not been described in detail.
Sex differences in the song bout length (SBL), total repertoire size (RS), and repertoire size within each phrase category for male and female starlings: whistle (W), variable (V), rattle (R), and high-frequency (H) phrase types. Phrase types sung separately from a song bout involved only whistles (W*). Sex differences were calculated using t-tests for independent samples, unless otherwise indicated
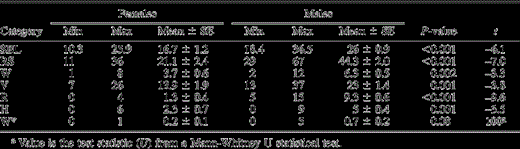
Sex differences in the song bout length (SBL), total repertoire size (RS), and repertoire size within each phrase category for male and female starlings: whistle (W), variable (V), rattle (R), and high-frequency (H) phrase types. Phrase types sung separately from a song bout involved only whistles (W*). Sex differences were calculated using t-tests for independent samples, unless otherwise indicated
As established previously for male starlings, females also produce a complex song. With some females singing song bouts of more than 30 seconds of uninterrupted song, female European Starlings probably have of one of the most complex singing behaviors in temperate-zone songbirds. In general, the organization of the song was similar in both sexes. All four categories of phrase types (whistle, variable, rattle, and high-frequency phrase types) typically presented in a male starling song bout were also found in the song of females. Hausberger et al. (1995a, 1995b) reported that the clicks and the high-pitched trills (corresponding to rattle and high-frequency phrase types) were mostly absent from females' repertoire, whereas in our study only two females produced neither rattles nor high-frequency phrase types and most possessed either one or the other song category. A possible explanation for this difference might be the difference in the methodology of recording female song: females were always recorded in individual cages and separate anechoic rooms in the studies of Hausberger et al. (1995a, 1995b) while in our study females were housed together in large social groups, which might better replicate their natural environment. This might suggest that rattles and high-frequency phrase types are important in a social context. In agreement with this, Chaiken et al. (1993) reported that 50% of tape-tutored male starlings produced abnormal rattle and high-frequency phrase types. Further studies should examine whether the observed differences in the occurrence of rattle and high-frequency phrase types in female starling song are due to population differences or social housing conditions. Finally, compared to males, a significantly lower proportion of females sang all four categories of phrase types.
Similar to males (Eens et al. 1992a), all 12 females incorporated heterospecific imitations in their song. Although we did not measure the occurrence of vocal mimicry quantitatively, it was clear that vocal mimicry is a common feature of the female starling song. In one case, almost half of the females shared a heterospecific imitation (ambulance siren). At this time, it is unclear whether this shared heterospecific imitation was learned from the heterospecific sound source, from conspecifics, or from both.
At present, we can only speculate as to why some females do not sing all four categories of song while others do. Individual variation could arise from differences in song-learning abilities or perhaps due to motor constrains on vocal performance (Podos 1996, Nowicki et al. 2002). Song learning has rarely been studied systematically and experimentally in female songbirds (Riebel 2003, but see Yamaguchi 1998, 2001), thus detailed data are lacking about when, from whom, and how long it takes for female starlings to learn their songs. As song requires complex learned motor skills involving coordination of the syringeal respiratory and musculature, producing sounds such as rattles and high-frequency phrases, might be costly. Suthers and Goller (1997) stated that rapidly trilled phrases with minibreaths (as the rattles) should be particularly costly to produce, since switching between expiration and inspiration involves increased muscular effort. Although it is possible that rattle and high-frequency phrases are more difficult or costly to produce than the two other categories, this requires further study. Finally, the observation that even after testosterone administration not all females were observed to sing high-frequency phrase types (see Hausberger and Black 1991, Hausberger et al. 1995a) may suggest that organizational rather than activational effects are responsible for this constraint.
Females that possessed all phrase categories were not necessarily those with the largest repertoire size. Given that song complexity is often measured in terms of the repertoire size, our results may suggest that the presence (or absence) of phrase-type category conveys other information than complexity of the song. The idea that different song features might hold specific information and serve a special function was first proposed by Marler (1960). Since this time, evidence has accumulated suggesting that there are different functions for different songs in male birds (Vallet et al. 1998, Leboucher et al. 1998), whereas, no data are as yet available showing whether this might hold true for females.
For both male and female starlings, it has been shown that they can acquire new phrase types in adulthood (Eens et al. 1992a, Eens 1997, Hausberger et al. 1995a, Mountjoy and Lemon 1995). Whether females that were observed singing only two out of four phrase type categories (whistles, variable phrase types) can produce the other two categories of phrase type (rattles, high-frequency phrase types) later in life remains to be investigated. The fact that all females were at least in their third calendar year when recorded, makes it highly unlikely that the observed individual differences in song composition are due to age differences.
All song measurements that we compared between sexes (total and phrase-type category repertoire size, song-bout length, and the presence of the four phrase-type categories in a song bout) were significantly higher in males than in females demonstrating that overall song complexity in males is higher than in females. In this respect, our results differ strongly from the results of Hausberger and Black (1991) and Hausberger et al. (1995a). They reported that the repertoire size of females is only slightly different from males, consisting of up to 16 whistles and between 8 and 34 warbling motif types (“motif” is the equivalent for “phrases” according to the terminology of Hausberger et al. 1995a; whistles and warbling motifs are considered separately). However, since their studies were carried out after testosterone treatment of females, we cannot determine whether this is due to population differences or to testosterone manipulation. Conversely, for the repertoire size of all phrase categories except for the rattle phrase types, we found an overlap between sexes indicating that some females have a more complex song than some males. Overlapping repertoire sizes have recently also been found in other temperate species (Yamaguchi 1998). Our findings of sex differences in song production are in line with previous findings that the volume of several song control nuclei in female starlings is smaller than in males, but also compared to other species, sex dimorphism in the song system is much smaller (Bernard et al. 1993).
For male starlings it has been shown previously that there is a highly significant positive relationship between repertoire size and average song bout length (Eens et al. 1991a). In females, however, no significant positive correlation was found. But since the power of the statistical test was not large, we also cannot rule out that these two traits were positively related. Our results also showed a substantial amount of variation among females in both repertoire size and song bout length. A large number of studies on male song production have emphasized the significance of individual variation for assessing male quality. At present, there is a little doubt that large repertoires in males have arisen through sexual selection by female choice (MacDougall-Shackleton 1997), whereas it is still unknown whether song complexity functions as a potential indicator trait in females (but see Langmore et al. 1996). Given that there is accumulating evidence that mutual mate choice may be much more common than previously thought (Johnstone et al. 1996, Amundsen 2000, Bergstrom and Real 2000, Jones et al. 2001), future studies should address whether female song signals quality in starlings and other songbird species.
Acknowledgments
DP was supported by a doctoral grant of the Fund for Scientific Research Flanders, Belgium (FWO-Flanders). This study was also made possible through financial support from the Research Council of the University of Antwerp (NOI-BOF UA 2002) and by a research project of the FWO-Flanders (G. 0420.02).
Literature Cited

{kind=link}
{kind=link}
{kind=link}