





Abstract
From 1999–2003, we tracked movements of adult female Prairie Falcons (Falco mexicanus) using satellite telemetry to characterize long-range movement patterns and breeding dispersal. We radio marked 40 falcons from April–May on their nesting grounds in the Snake River Birds of Prey National Conservation Area in southwest Idaho. All falcons with functioning transmitters left the Snake River Canyon from late June through mid-July. Most headed northeast across the Continental Divide to summering areas in Montana, Alberta, Saskatchewan, and the Dakotas. Prairie Falcons stayed at their northern summer areas for 1–4 months before heading to the southern Great Plains or to southwest Idaho. The Great Plains was a key migration pathway. Important wintering areas included the Texas Panhandle and southwest Idaho. Most falcons completed their seasonal migrations within 2 weeks. Use of widely separated nesting, summering, and wintering areas appears to be a strategy to exploit seasonally abundant prey resources. Most falcons had three or fewer use areas during the nonbreeding season, and falcons showed a high degree of fidelity to their use areas during each season. At least 21 falcons returned to nest within 2.5 km of where they nested in the previous year, but one falcon moved to a new nesting area 124 km south of her previous breeding area. Prairie Falcon movements suggest large-scale connectivity of grassland and shrubsteppe landscapes throughout western North America. Conservation of Prairie Falcons must be an international effort that considers habitats used during both nesting and non-nesting seasons.
Movimientos de Largo Alcance y Dispersión Reproductiva de Falco mexicanus del Sudoeste de Idaho
Resumen. Entre los años 1999 y 2003, seguimos los movimientos de hembras adultas de Falco mexicanus utilizando telemetría satelital para caracterizar los patrones de movimiento de largo alcance y la dispersión reproductiva. Marcamos con radios a 40 halcones desde abril hasta mayo en sus sitios de nidificación en el Área de Conservación Nacional de Aves de Presa Snake River en el sudoeste de Idaho. Todos los halcones con transmisores funcionando dejaron el cañón de Snake River desde fines de junio hasta mediados de julio. La mayoría se dirigió hacia el noreste a lo largo de la línea de división continental rumbo a las áreas de veraneo en Montana, Alberta, Saskatchewan y Dakotas. F. mexicanus permaneció en sus áreas de veraneo del norte durante 1 a 4 meses antes de dirigirse hacia las Great Plains del sur o hacia el sudoeste de Idaho. Las Great Plains representaron una ruta migratoria clave. Las áreas de invernada importantes incluyeron al Texas Panhandle y el sudoeste de Idaho. La mayoría de los halcones completaron su migración estacional en menos de dos semanas. El uso de áreas de nidificación, de veraneo e de invernada ampliamente separadas entre sí parece ser una estrategia para explotar presas estacionalmente abundantes. La mayoría de los halcones tuvo tres o menos áreas de uso durante la estación no reproductiva y los halcones mostraron un alto nivel de fidelidad a sus áreas de uso durante cada estación. Al menos 21 halcones regresaron a nidificar a menos de 2.5 km de donde nidificaron el año previo, pero un halcón se desplazó a una nueva área de nidificación ubicada a 124 km al sur de su área previa de cría. Los movimientos de F. mexicanus sugieren la existencia de una conectividad a gran escala de los paisajes de pastizal y la estepa arbustiva a través del oeste de América del Norte. La conservación de F. mexicanus debe ser un esfuerzo internacional que considere los ambientes usados durante las estaciones reproductivas y no reproductivas.
Introduction
Although much is known about the ecology of Prairie Falcons (Falco mexicanus) during the nesting season, little is known about movement patterns between breeding and non-breeding areas, and even less is known about the extent of interchange among nesting populations (Steenhof 1998). The migration period is a critical time for many birds, and data from both breeding and non-breeding periods are needed to manage populations of birds that migrate (Sillett and Holmes 2002).
The Snake River Birds of Prey National Conservation Area (NCA) in southwest Idaho contains the largest nesting population of Prairie Falcons in North America (Marti 1992, Kochert and Collopy 1998). Prairie Falcons use the NCA for only part of the year (Steenhof et al. 1984). To understand more about the ecology of Prairie Falcons during the non-nesting season, we used satellite-received transmitters (Platform Transmitter Terminals [PTTs]; Microwave Telemetry Inc., Columbia, Maryland) to track their movements throughout the year. Specific objectives of this study were to: (1) characterize annual long-range movement patterns of female Prairie Falcons that nest in the NCA, (2) identify areas used by Prairie Falcons during the non-nesting season, and (3) assess breeding dispersal patterns of female Prairie Falcons.
Methods
Each year we randomly selected trapping sites within the NCA (42°50′N, 115°50′W) from a list of historical territories occupied by Prairie Falcon pairs in previous years. We defined a historical nesting territory as any area of cliff where a Prairie Falcon pair was found in one or more years but where no more than one Prairie Falcon pair nested in the same year (Newton and Marquiss 1982). We allocated our sample size among four geographic areas within the NCA (west, west-central, east-central, and east; Steenhof et al. 1999), based on the number of pairs that historically nested in each of those four areas. We randomly assigned ranks to all territories identified since 1973 in each stratum, and then used rankings to determine at which territories we trapped and which were back-up sites. We rejected sites for trapping if: (1) the territory was not occupied, as verified by at least two surveys lasting a total of at least 5 hr; (2) field researchers determined that trapping at an occupied site would be dangerous to either the researchers or the birds; or (3) the target bird was not trapped after at least three attempts separated by approximately 1 week.
We trapped adult Prairie Falcons in April and May 1999–2002 using dho gaza nets with a Great Horned Owl (Bubo virginianus) as a lure (Bloom et al. 1992). We placed nets in occupied territories away from the nesting site but as close as possible to the cliff containing the nesting scrape, usually below the cliff. We captured and handled all birds according to Animal Care Protocol guidelines approved by Boise State University (IACUC Number 692–98–020). We frequently captured the male from the target trapping territory, and in some cases we also captured falcons from neighboring nesting territories. We placed hoods on all birds captured and held them in a metal can or an “aba” (Maechtle 1998) to minimize stress. We weighed and measured all captured birds, and we applied an aluminum U.S.G.S. band to the left leg and a black anodized band with white alphanumeric symbols (Acraft Sign & NamePlate Company, Edmonton, Alberta) to the right leg of all previously unbanded falcons. We classified birds with wing chords >320 mm as females (U.S. Department of the Interior 1977), and we classified adult falcons as “first-year birds” (11–13 months old) if the legs or cere were bluish or if the breast showed streaks rather than spots (Steenhof 1998).
Each year we attached PTTs to 10 adult female Prairie Falcons. We radio-tagged females only if they weighed at least 835 g and only if there was evidence that the trapped bird was a resident of the randomly selected target territory. The PTT-100 units (Microwave Telemetry Inc., Columbia, Maryland) weighed 33.6 ± 0.9 g (range: 32.1–35.5 g) and contained sensors that measured battery voltage, motion, and temperature. We attached PTTs using a backpack harness (Steenhof et al. 2006).
We programmed each PTT so that it would function for approximately 13 months with a 750-hr battery. PTTs transmitted approximately every 60 sec, at 401.65 MHz. We scheduled each PTT to transmit for 8 hr (hereafter a “transmission period”), every 3–7 days, with more frequent sampling during times of the year when we expected the most long-range movement. We programmed the duty cycle timer so that the PTT did not transmit for defined “off-periods” in each of four seasons: 167 hr in the first 10 cycles, 72 hr in the next 40 cycles, 114 hr in the next 23 cycles, and 80 hr in all remaining cycles. Actual transmission times varied with temperature and individual PTT. Most batteries lasted longer than their predicted 750 hr.
We used the ARGOS-TIROS satellite system (Strikwerda et al. 1986, Service Argos Inc. 2001) to obtain estimates of PTT locations and thereby track long-range movements of falcons. We received estimated locations, along with temperature, location class (LC), and activity sensor data, every four days via e-mail from Service Argos' Automated Distribution System. Accuracy of ARGOS-acquired locations depended, in part, on the number and distribution of transmissions received during a satellite pass. Location classes assigned by ARGOS (3, 2, 1, 0, A, B, Z) predict relative accuracy of the location estimates in decreasing order, based on the physics of the satellite telemetry system. However, Vincent et al. (2002) determined that accuracy of LC A locations was similar to LC 1 locations and better than LC 0 locations. Britten et al. (1999) determined that location estimates from similar-sized PTTs used on Peregrine Falcons (Falco peregrinus) were less accurate in field situations than the nominal error reported by ARGOS.
To delineate “use areas” during the non-nesting season, we visually inspected estimated locations of each bird in ArcView 3.2 and MS Access 2000 and identified obvious clusters that were grouped in space and time. We did not use data after a bird was known to have died or lost its transmitter (Steenhof et al. 2006), and we only used clusters that represented more than two transmission periods—or at least 10 days. Thus, this process excluded areas that falcons passed through briefly. We did not include the lowest-quality location estimates, B's and Z's, to define use areas, and we did not include location estimates that were >100 km from both the previous and subsequent location estimates.
We classified clusters of locations from 1 July through 15 September, including all associated location estimates within 100 km after those dates as “summer use areas,” and we classified clusters of locations from 10 December through 30 January, including all associated location estimates before and after those dates as “winter use areas.” For birds that did not migrate in fall, we used 15 October to divide clusters into summer and winter use areas. We based this cutoff on estimated departure dates from summer areas and arrival dates on winter areas of birds that migrated. We defined summer migration as all movements from nesting areas to summer use areas, fall migration as movements between final summer use areas and winter use areas, and spring migration as movements from final winter use areas back to nesting areas.
We used the median date between the last date that satellites received transmissions from the NCA and the first date that satellites received transmissions ≥100 km outside the NCA as the estimated departure date for each bird. We included data from one bird that dispersed to a nesting location outside the NCA (see results below) because the timing and patterns of its departure from that nesting area were similar to other birds we studied in the NCA. Similarly, we used the median date between the last date that satellites received transmissions from an individual's summer use area and the first date that satellites received transmissions ≥100 km from the summer area as the estimated fall departure date for each bird. We calculated spring departure dates as the median date between the last date that satellites received transmissions from an individual's winter use area and the first date that satellites received transmissions ≥100 km away.
During the 2000–2004 nesting seasons, we tried to find females that we had radio marked in previous years at nesting areas, using a combination of visual observations and radio-telemetry. We used telemetry location estimates to determine general locations of females wearing functional PTTs. We searched for marked birds in their previously used territories and, if necessary, in adjacent territories. Unless the bird was known to have died or moved to a different territory, we observed her previously occupied territory (or territories) for at least 5 hr on at least two visits or until we found unbanded falcons occupying the territory. Searches began in early March and continued through the breeding season. We also searched for marked birds throughout the canyon during surveys of abundance and productivity in 2002 and 2003 (USGS, unpubl. data), and we conducted intensive searches of areas where other scientists and technicians had reported incidental sightings of marked Prairie Falcons in the canyon. We used Leica APO Televid and Kowa TSN-820 spotting scopes with 20–60 zooms to spot antennas and read bands. We calculated breeding dispersal distances only if we were able to identify the nest, the mortality location, or the center of nesting activity in consecutive years. We tried to determine nesting success of all females that had been radio marked in prior years. We aged nestlings using a photographic aging key (Moritsch 1983) and considered nesting attempts successful if ≥1 young reached 30 days of age.
Statistical Analyses
We converted raw ARGOS data to database files and used Microsoft Access to sort and process data. We analyzed data with ARCView ver.3.2 and ARC GIS 9 Geographic Information System (GIS) software (Earth Systems Research Institute Inc., Redlands, California).
We calculated maximum travel periods and minimum travel rates during periods that individuals showed long-range (>100 km) movements. We based our calculations of time on the earliest signal received from a new area and latest signal received from a previously used area. To calculate distance, we selected a “representative” location estimate from each transmission period based on the location class (in order of preference: 3, 2, 1, 0, A, B). If there were two or more locations with the best location class, we used the one that occurred earliest in the transmission period. When necessary, we used B location estimates to identify timing and location of migration because we had few high-quality location estimates during the brief migration period and because the scale at which we were working was compatible with the additional possible error. We also assessed rates of travel from individuals that made significant long-range movements during a single transmission period. We used National Weather Service (2005) data to review weather conditions on days and times that falcons moved long distances; we used data from the weather station closest to the origin of a falcon's flight path on a given day.
We computed distances and azimuths moved by each falcon during long-range travel using a “great circle” formula that uses estimated radius of the earth to calculate distance between two geographic coordinates expressed as radians (D. Douglas, USGS, pers. comm.). We also used the great circle formula to calculate direct distances between use areas and NCA nesting locations. Our estimated minimum distances traveled during summer, fall, and spring migration periods were based on the sums of distances between the best location estimates from each transmission period that the bird was in transit during each of those periods. We computed total minimum distance traveled in a year by adding the estimated travel distances in each migration period. For birds with multiple summer use areas, we also included great-circle distances between centroids of successive use areas.
We analyzed movement directions using Oriana software (Kovach Computing Services 2004) for circular statistics. We used Systat 8.0 (SPSS 1998) to run two sample t-tests, analysis of variance, and Pearson correlations for assessing factors related to departure dates, travel rates, and distances moved. To compare nest success rates among groups, we created contingency tables and used StatXact 3.1 software (Cytel Software 1989) to calculate likelihood-ratio statistics (G-tests) and one-sided asymptotic P-values. Except where noted, values reported are means ± SD, and we used an α level of 0.05 for all tests.
Results
We set traps at 57 nesting territories from 1999– 2002, and captured 50 female Prairie Falcons. We did not radio-mark 10 of these females because six were underweight or in poor condition and four were from nontarget territories. We trapped 21 of the radio-marked females during incubation and 10 during brood rearing. We were unable to determine stage of nesting for nine radio-marked females. Two females that we radio-marked had been trapped and banded as adults in November 1998 in southwest Idaho. At least nine were probably first-year birds when we radio-marked them.
Data Quality
We received 15 972 transmissions with location estimates from April 1999 through September 2003: 5% were classed in the highest Location Class (LC) categories LC 2 or 3, 16% were LC 1, 42% were LC 0, 16% were LC A, and 21% were LC B or Z. The period in which we received useful data from PTTs on live birds ranged from less than 2 months to 21 months. The amount of data received varied among birds depending on mortality, PTT failure, battery life, and whether falcons had removed their harnesses and transmitters; in 1999, four falcons shed their harnesses before the nesting season ended, and two others shed them in October (Steenhof et al. 2006). We know at least two PTTs failed prematurely because we observed live birds carrying PTTs that stopped transmitting before their scheduled lifespan. Battery life of PTTs that functioned normally on live birds averaged 535 ± 58 days (range: 470–637 days). We obtained data for a second summer from 13 birds with functioning PTTs, and five PTTs continued transmitting through a second fall and into early winter.
Summer Migration
All falcons with functioning PTTs left the NCA after the nesting season. Departure dates ranged from 23 June to 20 July (mean = 7 July ± 7 days). Departure dates varied significantly by year (F4,41 = 6.0, P < 0.001), with birds departing earlier over time (r = −0.46, P < 0.001). Mean departure dates ranged from 2 July in 2002 to 13 July in 1999. Dates of departure did not differ significantly among study area strata (F3,41 = 1.1, P = 0.37), and departure dates of females that successfully raised young did not differ from females who did not produce fledging-age young (t = −1.1, P = 0.27, df = 41). The two earliest departure dates and the two latest departure dates were for birds that probably never laid eggs. Departure dates of successful females (n = 33) were not related to estimated fledging dates of their young (r = 0.21, P = 0.25). Some females left the NCA only 15 days after their young had left the nest. Departure dates of the same individuals in two consecutive years varied by 1–18 days (mean = 8.6 ± 6.3 days n = 11), with six leaving earlier in the second year and five leaving later. Departure date did not correlate with distance to the first summer area (r = 0.13, P = 0.38, n = 46). However, when we excluded the one individual that summered south of the NCA (see below), we found a significant relationship between departure date and latitude of the first summering area; birds that summered farther north tended to leave the NCA later (r = 0.33, P = 0.03).
Most falcons crossed the Continental Divide on the Idaho-Montana border northeast of the NCA (Fig. 1). Bearings of initial legs ranged from 309–108° (mean vector for all trips = 42°; 95% confidence interval = 34–50°; n = 46). Individual falcons appeared to use similar migration paths from year to year. However, our sample sizes were small and varying lengths of programmed PTT off-periods confounded our ability to compare travel rates and paths of the same individuals in consecutive years.
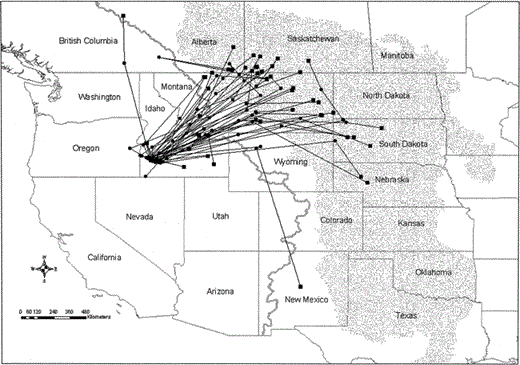
Summer migration routes of 34 female Prairie Falcons from southwest Idaho nesting grounds, June–July 1999–2003. Includes routes from two consecutive years for 12 individuals. Solid black circles represent the best location estimates for transmission periods and represent approximately 36% of the location estimates received during the migration period for each bird. Black squares depict the best location estimates during the first transmission periods within the summer use areas. Gray line represents the Continental Divide, and the stippled area depicts the Great Plains
Most falcons reached their summering areas within 1 week. The maximum number of travel days from nesting areas to summering areas ranged from 3–28 days (median = 7, mean = 9.4 ± 5.2 days, n = 46). Maximum travel days correlated positively with total distance traveled (r = 0.45, P < 0.01, n = 46). Mean minimum travel rate, based on time elapsed between the last location in the NCA and the first location on the summering area, ranged from 21–299 km per day. These estimates of travel rates are biased because they are influenced strongly by length of the PTT off-period and because of lack of information about deviations from straight-line segments between location estimates.
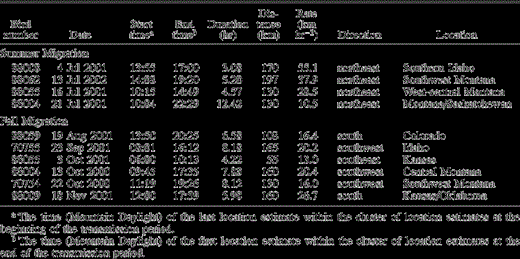
We obtained more specific information on travel times and rates from four individuals that showed relatively long directional movements on days in July when their PTTs were transmitting. These falcons covered up to 197 km in a day, with travel rates ranging from 10–55 km hr−1 (mean = 33.0 ± 18.6 km hr−1; Table 1). Most long-range movements occurred in the afternoon, suggesting that falcons may have relied on thermals for northeastward movements. One female did not leave her NCA nesting area until sometime after 13:55 (MDT) on 4 July 2001. Her estimated travel rate on that day (55 km hr−1) was faster than travel rates of individuals that had already departed the NCA on their northeastward migration (Table 1).
Rates and timing of movements of female Prairie Falcons on single days when PTTs transmitted during days with long-range movements. Based on location estimates with Location Classes A or better
Rates and timing of movements of female Prairie Falcons on single days when PTTs transmitted during days with long-range movements. Based on location estimates with Location Classes A or better
We documented only four cases in which falcons spent more than one transmission period at an intermediate location during summer migration. However, not all falcons took the most direct route to summering areas. At least 7 of 34 falcons flew >100 km longer than the straight-line distance between the nesting area and the summering area. One bird traveled >1000 km farther than the shortest route between nesting and summering areas. Most indirect routes involved an initial northeast bearing across the Continental Divide, then a turn to the northwest or the southeast. Two birds with indirect routes had stopovers. At least one bird took an indirect route in two consecutive years. Falcons taking indirect routes did not spend significantly more time traveling (mean = 12.5, range = 6–22 days) than those that followed direct routes (mean = 8.7, range = 3–28 days; t = 1. 7, P = 0.13, df = 9).
Summer Use Areas
Prairie Falcons summered in six U.S. states and three Canadian provinces (Fig. 2). We identified 50 summering areas used by 31 different falcons. Forty-four percent of use areas were in Montana, and 24% were in Saskatchewan. Falcons also summered in South Dakota, Idaho, Alberta, Nebraska, New Mexico, Wyoming, and British Columbia. Most summering areas were northeast of the NCA (range: 344°–135°). Bearings from nesting areas to 34 summer areas that falcons first used after leaving the NCA averaged 49.42° (SE = 5.15) and were significantly different from random (Rayleigh test of uniformity: P < 0.001). Twenty-seven of these 34 summer areas (80%) were 25°–82° from the respective nesting area. Distance between Prairie Falcon nesting areas and initial summering destinations averaged 822 ± 291 km. The closest summering area (140 km) was in Washington County, Idaho, and the most distant summering area (1287 km) was in central New Mexico.
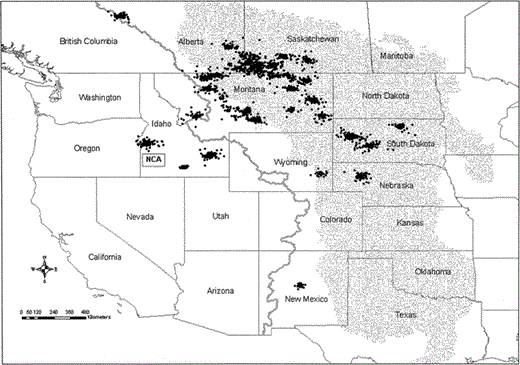
Location estimates of 31 satellite-tracked female Prairie Falcons within their summer use areas. Summer use areas are defined as clusters of locations that included least 10 consecutive days between 1 July and 15 September. The rectangle depicts the general location of the Snake River Birds of Prey National Conservation Area (NCA). Gray line represents the Continental Divide, and the stippled area depicts the Great Plains
Most falcons (28) had only one summer use area each year. Eleven of 28 falcons with single summer use areas survived with a functioning PTT into a subsequent summer, and all returned to the same general use area the following year. Three falcons had more than one use area during summer. Two of these birds survived with a functioning PTT for a second summer; both had similar use areas in the second summer, but the dates on which they used their summer areas varied. In addition to the three falcons with multiple summer areas, at least one falcon never established a use area during summer and moved nearly continuously.
Fall Migration
Prairie Falcons left their final summering areas as early as 14 August and as late as 6 December. Median fall departure date was 12 October (mean = 9 October ± 27 days, n = 29). Fall departure dates showed no correlation with latitude of the last summering area (r = −0.03, P = 0.88). Fall departure dates of the same individuals in two consecutive years varied by 0–38 days (mean = 17.4 ± 13.9 days, n = 5), with two leaving earlier in the second year, two leaving later, and one leaving on the same day in the second year.
Most falcons migrated along two distinct corridors in fall (Fig. 3). One corridor extended southwest from the summering areas and crossed the Continental Divide. The other went south-southeast into the southern Great Plains. The south-southeast corridor followed the boundary of the Short Grass Prairie and Central Mixed Grass Prairie Bird Conservation Regions (U.S. NABCI Committee 2000). Bearings of initial legs showed a distinct bimodal pattern (Fig. 3) with 15 falcons heading southeast and 14 heading southwest; no birds headed directly south. Bearings ranged from 108–246°. Although the mean vector for all trips was 182° (95% confidence interval = 168–196°; n = 29), we recorded no birds heading between 174° and 195° from their summering areas (Fig. 3). Bearings of birds that headed southwest averaged 217° (95% confidence interval = 209°–225°), and bearings of those that headed southeast averaged 149° (95% confidence interval = 137°– 160°). Falcons moved up to 1948 km between summer and wintering areas (mean = 1035 ± 396 km). The longest fall migrations were between summering areas in Saskatchewan and wintering areas in Texas.
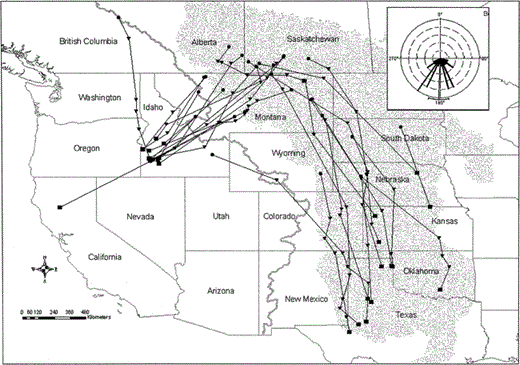
Fall migration routes of 24 satellite-tracked Prairie Falcons, August–December, 1999–2002. Includes routes from two consecutive years for 5 individuals. Solid black circles depict summer use areas, triangles represent the best location estimates for transmission periods during migration, and black squares depict the best location estimates during initial transmission periods within winter use areas. Inset depicts initial bearings of fall movements between summer and winter areas. Gray line represents the Continental Divide, and the stippled area depicts the Great Plains
Most falcons reached their wintering areas within 2 weeks. Maximum number of travel days from summer areas to winter areas ranged from 3.1–29.8 (median = 10.2, mean = 11.3 ± 6.0 days, n = 28). Maximum number of travel days correlated positively with total distance traveled (r = 0.47, P = 0.01, n = 28). Estimated mean minimum travel rate, based on time elapsed between the last summer location and the first location on the wintering area, ranged from 30–205 km day−1.
We obtained more specific information on travel times and rates from six individuals that showed relatively long directional movements on single days during fall migration (Table 1). All these movements occurred on days after birds had left their summering areas. These falcons covered up to 165 km in a day, with travel rates ranging from 13–27 km hr −1 (mean =18.8 ± 4.8 km hr −1; Table 1). Each of the six birds had tail winds for all or part of their travel periods. Travel rates during fall migration were slower than rates during summer migration. In contrast to summer migration, birds migrated throughout the daylight period during fall and showed no tendency to travel mainly during the afternoon (Table 1). Also in contrast to summer migration, most falcons took a relatively direct route to wintering areas, and only one of 23 followed a route that was >100 km longer than the straight-line distance between the summering and wintering areas. Only four falcons spent more than one transmission period in a given area during fall migration.
PTTs on five individuals provided data on two consecutive fall migrations. All five falcons used similar southward migration paths in consecutive years. Minor differences in travel routes of the same individuals in consecutive years may have resulted from differences in sampling intervals rather than from differences in actual routes traveled.
Winter Use Areas
Falcons wintered in nine states (Fig. 4). Idaho had the most wintering birds (10), followed by Texas (7), Kansas (3), and Montana (2). Nebraska, Oklahoma, South Dakota, New Mexico, and California each had one falcon during winter. We identified 33 wintering areas used by 27 different Prairie Falcons. Four falcons used the same areas in summer and winter (1 each in Idaho, Montana, South Dakota, and New Mexico). Three of the four falcons that did not migrate in fall were first-year birds when we radio marked them.
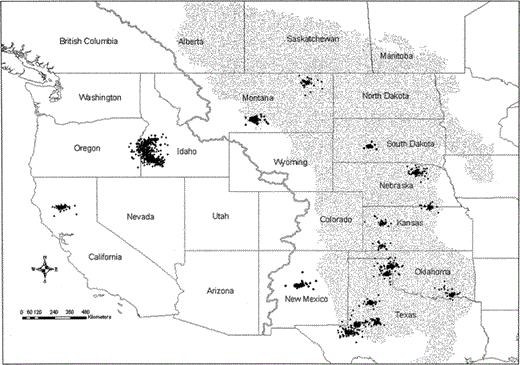
Location estimates of 27 satellite tracked female Prairie Falcons within their winter use areas August 1999–February 2003. Use areas are defined as clusters of locations that included at least 10 consecutive days between 10 December and 30 January. Gray line represents the Continental Divide, and the stippled area depicts the Great Plains
Distances from wintering areas to NCA nesting areas ranged from 11–1977 km (mean = 920 ± 735 km). Fourteen of 27 falcons wintered >1000 km from their nesting areas. Ten wintered in southwest Idaho within 200 km of their nesting areas, and five of these had winter use areas within the NCA. Mean arrival dates on wintering areas were similar for birds that wintered in the Great Plains (13 October ± 31 days, n = 12) and birds that wintered in Idaho (18 October ± 14 days, n = 9; t =1.5, P = 0.15, df =16.3), but arrival dates on the Great Plains showed more variation. Falcons arrived on Great Plains wintering areas as early as 24 August and as late as 9 December. In contrast, no falcons arrived on Idaho wintering areas until 5 October, and none arrived after 18 November.
Five falcons survived with a functioning PTT into a second winter, and all five used the same areas they had used in previous winters. We knew that two of these five falcons had used the same winter areas prior to radio marking because they were initially banded within their winter use areas in November 1998. Only one falcon had more than one use area in the same winter.
Spring Migration
Female Prairie Falcons remained on their wintering areas east of the Continental Divide as late as 8 March. Estimated departure dates from these distant wintering areas ranged from 25 February to 8 March (mean = 3 March ± 4.5 days, n = 9). Four falcons that wintered in the foothills north of the NCA returned to the NCA much earlier in spring: estimated departure dates from wintering areas in southwest Idaho but outside the NCA ranged from 15 January to 15 February (mean = 2 February ± 13.3 days). These falcons took less than one transmission period to move from their wintering areas to their NCA nesting areas. Instead of retracing summer or fall migration routes, each falcon flew directly from its wintering area east of the Continental Divide to the NCA. None of the falcons deviated >100 km from a straight-line distance from their wintering area to the NCA (Fig. 5). In contrast to summer migration, none of the migrating falcons spent more than one transmission period at an area en route to the NCA in spring. Most falcons completed the return trip to the NCA within 2 weeks. Maximum number of travel days from wintering areas east of the Continental Divide to NCA nesting areas ranged from 3–16 days (median = 10; mean = 10 ± 4 days). We found no correlation between distance traveled and maximum number of travel days (r = 0.56, n = 7, P = 0.20), probably because sample sizes were small and transmission periods did not coincide with travel times. Mean minimum travel rate, based on time elapsed between the last winter location and the first location on the nesting area, ranged from 109–290 km per day.
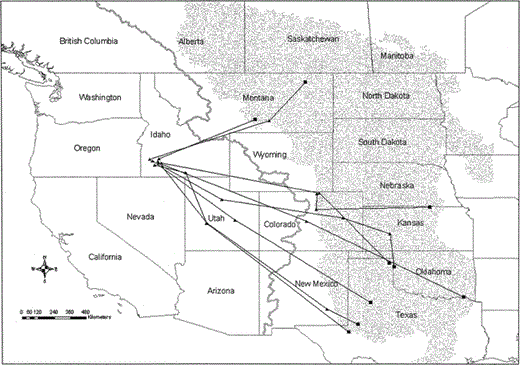
Routes used by nine Prairie Falcons to return to NCA nesting areas from wintering areas east of the Continental Divide in February and March, 2000–2003. Black squares depict the best location estimates for the final transmission periods in winter use areas. Solid triangles depict the best location estimates for transmission periods during and at the end of the travel period. Gray line represents the Continental Divide, and the stippled area depicts the Great Plains
Total Distances Traveled
The total distance traveled among use areas during a year ranged from 281 to 4636 km (mean = 2602 ± 1305 km, n = 17). The bird with the longest travel route summered in Saskatchewan and wintered in Texas. The bird with the shortest travel route spent the entire non-nesting season in Idaho, 140 km north of the NCA. Mean distance moved by six falcons that summered in the northern Great Plains and wintered in the southern Great Plains (4037 ± 500 km) was significantly greater (t = 7.9, P < 0.001, df = 12) than the mean distance moved by eight that summered in the northern Great Plains and wintered in Idaho (1892 ± 508 km).
Nesting Seasons Following Radio Marking
We confirmed locations of 24 Prairie Falcons during the first breeding season after they were radio marked. Sixteen falcons that returned to the NCA in the nesting season after they were radio marked were still wearing functioning PTTs. Five of eight individuals that overwintered in southwest Idaho nested successfully in the year after marking compared to only three of eight that wintered elsewhere (G = 1.0, P = 0.15). Breeding dispersal distances in the year following trapping ranged from 0 m–124 km (n = 22 because the original nesting locations of two individuals marked in 1999 were unknown). Mean dispersal distance was 6.1 ± 2.6 km, but median dispersal distance was only 0.3 km. All but one of 24 falcons returned to the NCA. The exception was an individual marked in 2000 that dispersed in 2001 to the South Fork of the Owyhee River along the Idaho-Nevada border, 124 km south of the NCA. She was observed on the same section of cliff in the South Fork Owyhee River Canyon during the 2001, 2002, and 2003 nesting seasons; she was paired with a mate and showed signs of nesting there in 2002 and 2003.
Within the NCA, most falcons returned to the same territory that they used in the previous year or to an adjacent one. Distances between nests or centers of nesting activity ranged from 0– 2155 m in the year following radio marking (mean = 457 ± 502 m; median = 246 m) and 29–954 m in subsequent years. Females sometimes nested in a section of cliff that was used by a neighbor in other years. We found little evidence for mate fidelity in our sample of marked females. At least five females that wore PTTs mated with a new male in the year following radio marking. At least one of these mate changes involved a male that had died, and at least two involved a male that paired with a different female the second year.
Discussion
Our findings confirm Enderson's (1964) suggestion that most Prairie Falcons have three distinct seasonal use areas. However, in contrast to Enderson's results, we confirmed that adult female Prairie Falcons show a high degree of fidelity to each of their seasonal use areas. All birds we monitored in more than one season returned to the same general area that they used in that season the previous year.
Use of postnesting areas prior to fall migration is unusual but not unique among raptors. Juvenile Bald Eagles (Haliaeetus leucocephalus) and Eurasian Kestrels (Falco tinnunculus) move north after fledging (Broley 1947, Schifferli 1965, Hunt et al. 1992), but postnesting movements of adults are less common. Like female Prairie Falcons, adult and juvenile Ferruginous Hawks (Buteo regalis) from Washington, Idaho, and Utah exhibited a generally clockwise annual migration that involved eastward and northward movements to postnesting areas followed by southward movements to wintering areas and a return trip to the nesting area (Schueck et al. 1998, Watson 2003). Plissner et al. (2000) offered three possible explanations for northward postbreeding movements of adult American Avocets (Recurvirostrata americana) that nest in the Great Basin: (1) a more favorable environment for molting, (2) the opportunity to assess and evaluate potential new breeding areas, and (3) the opportunity to exploit seasonally superabundant food resources. It is unlikely that the first two explanations apply to Prairie Falcons. Although Prairie Falcons probably molt most of their flight feathers while on their summer use areas (Enderson 1964), they do not lose the ability to fly, and they would not benefit from the types of predator-safe environments that waterfowl use during postbreeding molt migrations (Berthold 2001). And although many areas used by NCA Prairie Falcons during summer were near other breeding Prairie Falcons, we have no evidence that any individuals ever dispersed to nest in these areas. The one female who moved to a new breeding area explored that area in late winter, not during the postbreeding season.
Migration from the NCA to a postnesting area in summer is likely a strategy for Prairie Falcons to cope with seasonal declines in food availability. Piute ground squirrels (Spermophilus mollis, formerly S. townsendii) are the principal prey of Prairie Falcons during the nesting season (Steenhof and Kochert 1988). Each year ground squirrels begin 6–8 months of underground torpor in late May and become completely unavailable to falcons in the NCA by early July (Van Horne et al. 1997). In addition, warmer temperatures in July and August alter the activity cycles of alternative prey (Diller and Johnson 1982) making them less available to avian predators.
Although changes in prey availability may be the ultimate explanation for the falcons' summer migration, a combination of factors likely act as proximate cues to trigger a falcon's departure each year. Dunstan et al. (1978) reported that two adult females left the NCA on 12 and 13 July 1976 and that eight other radio-marked females were last recorded in the NCA between 19 June and 19 July 1975–1977. The consistent period in which females departed from the NCA in different years suggests that circannual factors may influence timing of departure (Berthold 2001). The fact that females departed soon after young fledged suggests that males might be responsible for providing food to fledglings, as has been confirmed in other raptor species (Watson 2003). Dunstan et al.'s (1978) data on departure dates and last contact dates of 18 radio-marked Prairie Falcons (including four pairs with both sexes marked) also suggest that adult females left the NCA before males.
The distribution of Prairie Falcon summering areas closely matches the distribution of Richardson's ground squirrels (Spermophilus richardsoni; Hall 1981), probably an important prey species for Prairie Falcons on their summer areas. Juvenile Richardson's ground squirrels emerge in May (Michener 1985) and are above ground through late September in southern Alberta (Michener 1979), so they are available to falcons through the entire summer. Richardson's ground squirrels are the primary prey species of Prairie Falcons that nest in southwestern Wyoming (MacLaren et al. 1988) and southern Alberta (Hunt 1993). Ferruginous Hawks from Washington feed extensively on Richardson's ground squirrels during the postnesting season in Alberta, Saskatchewan, and Montana (Watson 2003), and Enderson (1964) described a Prairie Falcon postnesting area on the Laramie Plains of Wyoming where Richardson's ground squirrels, Horned Larks (Eremophila alpestris), thirteen-lined ground squirrels (Spermophilus tridecemlineatus), and white-tailed prairie dogs (Cynomys leucurus) were abundant.
The length of time that falcons spent at summer areas varied considerably among individuals. Departure dates from summer areas showed a much wider range (>4 months) than departure dates from nesting areas (<1 month), and three falcons stayed on their postnesting areas until the next nesting season. We suspect that food availability probably influenced how long falcons stayed at their postnesting areas. Most female Prairie Falcons left their summer areas in September or October. This corresponds to the time when juvenile Richardson's ground squirrels typically hibernate (Michener 1979). Falcons that stayed at summer areas after 15 October likely had abundant alternative prey; falcons that departed before September may have been in areas with low ground squirrel numbers or poor ground squirrel reproduction.
Falcons that left their summering areas between August and December traveled within two distinct corridors. Like falcons nesting in the Canadian prairie provinces (Schmutz et al. 1991), they either headed southeast or southwest, thereby avoiding the forested Rocky Mountains. Many moved along the boundary of the Short Grass Prairie and Central Mixed Grass Prairie Bird Conservation Regions. Schmutz et al. (1991) recognized the Prairie Falcon's affinity for grasslands during migration and speculated that grasslands provided higher prey abundance than other habitats. Enderson (1964) believed that their low-level hunting style limits Prairie Falcons to open habitats in suitable terrain with no tall vegetation.
Travel rates of female Prairie Falcons were similar to those reported for Peregrine Falcons. Satellite tracked Peregrine Falcons covered an average of 141–204 km per day during migration (Fuller et al. 1998, McGrady et al. 2002, Ganusevich et al. 2004). During travel between seasonal use areas, Prairie Falcons moved an average of 21–299 km per day. Prairie Falcons were similar to Peregrine Falcons (Fuller et al. 1998, McGrady et al. 2002) in that travel rates during fall migration were slower than rates during spring migration. The relatively direct, rapid travel of Prairie Falcons from winter to nesting areas is similar to what has been observed in other species (Berthold 2001). Prairie Falcon migration is probably similar to that of Peregrine Falcons in that they spend part of their days hunting, and total distance moved per day varies with wind speed, wind direction, cloud cover, precipitation, and the availability of thermals (Cochran 1975). Within a single day we documented travel rates of 13–55 km hr−1 for Prairie Falcons, similar to Peregrine Falcon flapping flight speeds of 35–70 km hr−1 recorded by Cochran and Applegate (1986).
Female Prairie Falcons from the NCA used widely separated wintering areas on both sides of the Continental Divide. Falcons from southwestern Idaho wintered in the same general regions of the southern Great Plains where breeders from Canada and Wyoming wintered (Schmutz et al. 1991). Ten of 27 falcons returned to southwest Idaho to winter, and five of these wintered within the NCA boundaries. The strategy of wintering in the southern Great Plains likely evolved because the energetic benefits of wintering in a warmer climate with abundant prey must have outweighed the costs of migrating a longer distance. Radio-marked falcons that wintered east of the Continental Divide arrived on nesting areas 2–3 weeks later than falcons that wintered in Idaho. These long-distance migrants may have been at a disadvantage once they returned to the nesting grounds; their nesting success tended to be lower than for birds that wintered <200 km from nesting areas. Holthuijzen (1989) found that falcon productivity in the NCA was inversely related to hatching date. We observed that migrants were forced to compete for mates and nesting territories with falcons that had arrived earlier. Late arrival of long-distance migrants may have resulted, in part, from the effects of radio marking (Steenhof et al. 2006). Alternatively, wintering southeast of the NCA may have been more effective in the past when Idaho winters were more severe (NOAA, unpubl. data). Recent changes in climate may mean that selective pressures now favor falcons that winter near the nesting area or those who minimize travel costs by wintering at their northerly summer areas. Given the variety of movement patterns exhibited by our radio-marked sample, Prairie Falcons might be in a period of microevolutionary change from which a new pattern of annual movements will emerge (Berthold 2001).
We found limited evidence for Schmutz et al.'s (1991) suggestion that Prairie Falcons move in random directions and stop only when patchily distributed prey is encountered in abundance. Only three falcons in our study had more than one summer use area, and only one falcon moved continuously. Exploring may be more common in birds during their first year of life (Berthold 2001). Movements of juvenile Ferruginous Hawks are more extensive and wide-ranging than those of adults, and juvenile Ferruginous Hawks do not use well-defined postnesting areas (Watson 2003). Raptors may need to learn where to find resources as juveniles, and fidelity to use areas and migration routes may develop later in life. The female Prairie Falcons we tracked had already experienced at least one annual migration cycle. However, the youngest birds we tracked were amongst the most mobile and sedentary of our sample. Three falcons marked as first-year birds did not leave their summer areas in winter, and three moved extensively during summer. Another had a single summer use area and a single winter use area; it was the only bird from our sample that wintered in California, and it died there.
Migration patterns vary with age and sex in some bird species (Kerlinger 1989, Berthold 2001). However, information from other studies suggests that adult male and juvenile Prairie Falcons might have migration patterns similar to those we found for adult females. During the 1970s, radio-marked juvenile females and adults of both sexes departed from the NCA in late June and early July and headed generally northeast (Dunstan et al. 1978). Enderson (1964) noted a strong tendency for juvenile Prairie Falcons to move eastward from mountain nests in Colorado and Wyoming to the plains provinces and states following the nesting season, and Schmutz et al. (1991) reported a fledgling from Alberta heading eastward into Saskatchewan after the nesting season. Two falcons banded as nestlings in the NCA were found in Montana in July (Steenhof et al. 1984). Prokop and Marzluff (1994) confirmed that some female Prairie Falcons that breed in the NCA also spend the winter in the NCA, but they found that males were more likely to winter in the NCA than females. However, at least some NCA males migrate to the southern Great Plains; a male Prairie Falcon banded and radio marked as a breeder within the NCA in 1993 was found dead in the Texas Panhandle in December 1994 (U.S. Geological Survey, unpubl. data).
We documented the first case of long-range breeding dispersal in Prairie Falcons. Previously reported breeding dispersal distances of female Prairie Falcons, based on sightings and retrappings, ranged from 0.1–14.4 km for birds marked in the NCA (n = 20; Lehman et al. 2000) and 0.2–16.9 km for birds marked in Alberta, Colorado, and Wyoming (n = 9; Runde 1987). The 124-km breeding dispersal distance we documented exceeded the longest natal dispersal distance (35.6 km) recorded so far for Prairie Falcons marked in the NCA (Lehman et al. 2000), but was within the range of natal dispersal distances reported for falcons from Alberta and Wyoming (0–225 km; Runde 1987). Our new information suggests that although long-range breeding dispersal is rare (<5% of cases), interchange among populations from geographically separated nesting areas is possible.
During each annual cycle, individual Prairie Falcons relied on widely separated landscapes throughout western North America. Prairie Falcon movements suggest large-scale connectivity of grassland and shrubsteppe landscapes. None of the landscapes used by falcons provided the necessary resources in all seasons of the year. Effective management of Prairie Falcons that nest in the NCA must consider conditions in and access to habitats that are important during the non-nesting season. Conservation of Prairie Falcons and their habitats is clearly an international challenge and will require cooperation of agencies from the U.S. and Canada.
Acknowledgments
This study was funded by the U.S. Geological Survey, Forest and Rangeland Ecosystem Science Center, with additional support from Boise State University. The study would not have been possible without our tenacious trapper, J. McKinley. Each year we were fortunate to have the assistance of talented and dedicated volunteers: D.Centili, G. Campbell, N. Batbayar, A. Jacob, and B. Davies provided valuable assistance throughout the nesting season with locating pairs, trapping, and monitoring productivity. We were fortunate to have M. Vekasy (HawkWatch International) as a trapper in 2000. Special thanks goes to E. Hynes whose ability to read color bands provided us important data. B. Lehman, K. Ross, M. Waltman, M. McCaustland, L. Greenwood, S. Alsup and J. Ferrell, L. Ridenhour, J. Doremus, B. Mattox, L. Carpenter, B. Haak, Marc Bechard, L. Leppert, L. Schueck, R. Brady, J. Shook, and L. Tapia also assisted in the field. We thank Idaho Power Company for providing a camping site for our volunteers, and P. Howey and the Microwave Inc. staff for technical assistance. S. Finn and C. Meinke (U.S. Geological Survey, Snake River Field Station) provided valuable assistance in our GIS analyses. J. Schmutz and an anonymous reviewer provided comments and suggestions that improved the manuscript
Literature Cited

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}