





Abstract
We studied the breeding diet and hunting behavior of Crane Hawks (Geranospiza caerulescens) in Tikal National Park, Petén, Guatemala in 1994 and 1995. We observed 227 prey items while conducting observations at six nesting attempts, and during opportunistic sightings in the breeding season. Among 181 identified prey items, rodents comprised 47.5%, lizards 19.9%, frogs 16.0%, bats 6.6%, birds 6.1%, and snakes 2.8%; a juvenile skunk also was represented. Rodents accounted for 77% of estimated biomass, including at least eight species representing terrestrial, cursorial, and arboreal habits. More than half of all prey items weighed <20 g, but 40% weighed >50 g; many were nocturnal species presumably taken from daytime hiding places. We observed hunting attempts in all strata of the forest and in several forest types. Hunting behavior included still-hunting from a perch and probing with head or feet in holes, bromeliads and other epiphytes, palm leaf axils, crotches of branches, behind bark in living and dead trees, and in puddles. Compared to other raptors studied at Tikal, the Crane Hawk had a moderately broad food niche that overlapped most with other raptors deemed dietary generalists. However, the Crane Hawk's unique anatomical features and hunting behavior enabled it to capture diurnally reclusive prey presumably unavailable to many other raptors, thus facilitating relatively low dietary overlap.
Resumen
Resumen. Estudiamos la dieta y el comportamiento de cacería del Gavilán Ranero (Geranospiza caerulescens) durante 1994 y 1995 en el Parque Nacional de Tikal, Petén, Guatemala. Observamos 227 presas durante observaciones de seis nidadas y otras observaciones oportunas en la estación reproductiva. De 181 presas identificadas, roedores constituyeron el 47.5%, lagartijas 19.9%, ranas 16.0%, murciélagos 6.6%, aves 6.1%, y culebras 2.8%; un zorrillo infantíl también fue representada. Roedores constituyeron el 77% de la biomasa estimada, e incluyó al menos de ocho especies con hábitos terrestres, cursoriales, y arbóreos. Más de la mitad de las presas pesaron <20 g, pero el 40% pesaron >50 g; muchas fueron especies nocturnas, por presunción capturadas de escondites durante el día. Observamos intentos de cazar por todas partes del sotobosque y en el dosel en varios tipos de bosque. El comportamiento de caceria incluyó posando quieto en una percha, y buscando con cabeza y pies entre hoyos, bromélias y otras epífitas, hojas de palmas, bifúrcaciones de ramas, detrás de la cortesa de árboles ambos vivos y muertos, y en charcos. En comparación con otros rapaces estudiados en Tikal, el Gavilán Ranero tenía un nicho alimenticio algo amplio que coincidió más con rapaces considerados como generalistas dietéticos. Sin embargo, la anatomía singular del Gavilán Ranero y su comportamiento de cacería permite la captura de presas con hábitos diurnamente secretos que probablemente son indisponibles a otras rapaces, así facilitando menos coincidencia de la dieta entre ellos y los demás especies.
Introduction
Neotropical forests house some of the most species-rich raptor assemblages anywhere (Thiollay 1985), but the biology and ecology of many of these raptors remain poorly known (Bierregaard 1995). One Neotropical raptor that has received little study is the Crane Hawk (Geranospiza caerulescens). The Crane Hawk is a slender, long-legged hawk occurring in lowland forests and swampy woodlands from Mexico south to northern Argentina and southern Brazil (Brown and Amadon 1968). Like the harrier hawks (Polyboroides spp.) of Africa and Madagascar, the Crane Hawk possesses relatively long legs with extremely flexible intertarsal joints. Both genera use this unique tarsal flexibility to extract prey from cracks, hollows, holes, and recesses (Friedmann 1950, Sutton 1954, Brown and Amadon 1968). Although brief reports have documented that Crane Hawks eat a wide variety of small to medium-sized prey (Haverschmidt 1962, Jehl 1968, Bokermann 1978), quantitative data on the species' diet and hunting behavior are lacking. Crane Hawks are generally regarded as uncommon across their range and display great geographic variation in plumage coloration and body size (del Hoyo et al. 1994). The Black Crane Hawk (G. c. nigra), one of the largest subspecies, is a year-round resident of lowland forests from Sinaloa and Tamaulipas, Mexico, to the Panama Canal Zone (del Hoyo et al. 1994). Here we provide data on the diet and hunting behavior of nesting Black Crane Hawks at Tikal National Park, Guatemala, and on dietary niche-breadth and overlap between this and other similarly sized sympatric raptors.
Methods
Study Area
We studied Crane Hawks in Tikal National Park, Petén, Guatemala in 1994 and 1995 as part of The Peregrine Fund's “Maya Project.” Tikal National Park covers 576 km2 in northeastern Guatemala (17°13′N), and is characterized by low, eroded limestone hills ranging from 160–450 m above sea level. During the rainy season, which begins from mid-May to late June and lasts through January, extensive areas of low-lying forest are temporarily flooded, and seasonal water courses swell with runoff from the surrounding karst hilltops. Semi-permanent rain-filled ponds are the only source of year-round surface water. The tropical climate is relatively dry with a mean annual precipitation of 1,350 mm and a pronounced dry season from February to May or June. The structurally and floristically mature forest within the park is classified as tropical semi-deciduous (Pennington and Saruhkan 1968) and varies predictably along a topographic gradient, apparently due to strong correlations between soil characters, topographic position, and degree of inundation (Schulze and Whitacre 1999). Additional details on vegetation, physiography, climate, and land-use patterns of Tikal and the surrounding area are described by Whitacre et al. (1995) and Schulze and Whitacre (1999).
Data Collection
We gathered data on food habits by observing prey deliveries at six breeding attempts and in six opportunistic sightings during the breeding season. We observed nests with 15–60× spotting scopes from 35–60 m distant in blinds built in trees or on the ground. Nests were in dense vine tangles and Crane Hawks returning with prey rarely flew directly to them; we were often able to identify prey when the returning hawk landed in the nest tree and walked or hopped several meters to the nest. Both adults and nestlings ate prey delivered to the nest. We also collected fresh prey remains during nest climbs. We identified prey items to the most precise taxonomic level possible, visually estimated total body length of each prey item (by comparison with the hawk's bill and head size), and placed each into one of four estimated weight classes: >0–20, >20–50, >50–100, and >100 g. During this and other studies at Tikal, potential prey from various taxa, including snakes, lizards, amphibians, birds, bats, and other small mammals, were captured, weighed, and measured (Whitacre et al., unpubl. data, M. Jolón, pers. comm.). We used these data and other published sources (Campbell and Vannini 1989, Emmons 1990) to become familiar with and identify many prey items to the species level. Biomass estimates were based on measurements and adult body masses from the available data set and published literature. To attain higher resolution in our biomass estimates, we placed prey species into modified weight classes (>0–10, >10–20, >20–35, >35–50, >50–75, >75–100, >100–150, and >150 g) a posteriori, assigning each the median weight for the weight class. Values presented are means ± SD.
We collected data on hunting behavior while following radio-tagged hawks and during chance encounters. During observations of hunting attempts, we described behavior and inclination of the hawk's body to the foraging substrate, and recorded foraging height using three categories (0–5, >5–15, and >15 m). To quantify intertarsal joint flexibility, we used a protractor to measure backwards flexion and lateral and medial swing of tarsi in captured, live Crane Hawks. Classification of forest types at hunting sites followed Schulze and Whitacre (1999).
Based on identified prey of Crane Hawks and similarly sized raptors at Tikal, we calculated indices of food-niche breadth and overlap following Levins (1968) and Pianka (1974), respectively. Levins' measure of niche breadth is a more interpretable form of Simpson's (1949) diversity index (Marti 1987), incorporating both richness and evenness, with larger values reflecting greater diversity of prey species in the diet. Pianka's overlap index yields values from 0 to 1, indicating dissimilar (0) to similar (1) diets.
Results
Diet
Because there was no difference in the frequency of major prey types (i.e., rodents, lizards, frogs, bats, birds, snakes, and unidentified) captured in 1994 and 1995 (χ26 = 10.7, P = 0.10), or in the frequency of prey types (χ27 = 3.4, P = 0.85) or weight classes (χ26 = 3.2, P = 0.79) captured by males and females, we pooled all data to describe prey brought to adult females and nestlings. We identified 181 of 227 observed prey items (Table 1). Rodents (Muridae and Heteromyidae) comprised the largest portion of the diet. Terrestrial, cursorial, and arboreal rodents of at least eight species occurred frequently in Crane Hawk diets. Rice rats (Oryzomys spp.), spiny pocket mice (Heteromys spp.), and vesper rats (Nyctomys sumichrasti) accounted for the majority of rodents in the diet. Lizards and frogs were commonly taken, and together made up nearly as large a numerical proportion as rats and mice, although they contributed far less in terms of biomass. Lizards in the diet averaged 14 ± 6 cm in estimated length (range 8–25 cm, n = 28), and at least four species were taken (Table 1). Frogs were the only amphibians identified as prey; most were small and some had toe pads (possibly Hylidae, Leptodactylidae, or Microhylidae). Frogs were sporadically abundant in Crane Hawk diets when anuran activity increased after rains, especially in late May and June. Bats, birds, snakes, and a skunk made up the remaining identified prey items. Bats and birds were both twice as frequent in Crane Hawk diets as snakes. Snakes averaged 75 ± 28 cm in estimated length (range 40–100 cm, n = 5). All snakes were slender and green to brown in color; we believed most were arboreal species. On one occasion, we observed a female Crane Hawk deliver a juvenile, striped hog-nosed skunk (Conepatus semistriatus) to her young.
Frequency and biomass of prey items in the breeding diet of the Crane Hawk in Tikal National Park, Guatemala, 1994–1995. Percentages based on identified prey items

Frequency and biomass of prey items in the breeding diet of the Crane Hawk in Tikal National Park, Guatemala, 1994–1995. Percentages based on identified prey items
We identified 173 prey items at four nest sites; we observed rodents, lizards, and frogs at all four nest sites, birds at three, bats and snakes at two, and a skunk at one. Based on identified prey, the numerical contribution of rodents to the diet at these nest sites was 48.9 ± 11.2% (range 37.3–60.7%), lizards 17.8 ± 6.7% (range 8.3–22.9%), and frogs 16.7 ± 3.9% (range 12.0–21.4%). Nests did not differ in the frequency of these predominant prey types (χ26 = 4.0, P = 0.67). Unidentified prey comprised 10.7–58.6% of observed prey (25.3 ± 22.6%). Of the 224 prey items we assigned to estimated weight classes, the majority (52.2%) were estimated to weigh <20 g, but over a third (39.7%) weighed >50 g. Rodents, the most frequent prey type observed, comprised most of the biomass as well (77.3%; Table 1). In contrast, lizards, the second most-frequent prey type, contributed only a small fraction (2.8%) of prey biomass.
Hunting Behavior
We documented 19 hunting attempts by radio-tagged and unmarked Crane Hawks representing at least seven individuals. In 15 of 19 cases, we observed Crane Hawks actively searching for or pursuing prey. In three cases, Crane Hawks pursued prey on foot along the top of large tree branches, whereas in 12 cases we observed them probing and searching for prey in a variety of places. In these 12 cases, Crane Hawks explored holes in snags, tree trunks, and branches, searched in bromeliads, other epiphytes, and behind bark, and probed in palm leaf axils, in leaf litter on the forest floor, and in a shallow puddle. While searching these locations, body inclination of Crane Hawks was upright in four cases, clinging to the trunk or branch in five cases, and hanging nearly upside down in three cases. We observed Crane Hawks scaling trees in the manner of a woodpecker, reaching their legs into holes while propping their tails against the trunk and balancing with extended wings. The hawks usually used their long legs to grope within potential prey refugia, but occasionally used their head or bill. In 3 of 19 hunting observations (16%), Crane Hawks perched quietly, visually searching the forest floor from the canopy's lowest branches. On one occasion, we encountered a Crane Hawk while it was trying to extract an Empidonax flycatcher trapped in a mist net.
Crane Hawks hunted in all strata of the forest. We located Crane Hawks foraging near the forest floor (0–5 m: 31.5%), in the sub-canopy (>5–15 m: 37%), and in tree crowns (>15 m: 31.5%). We observed Crane Hawks hunting in upland standard forest (42%), which predominated in our study area, and in dry upland forest on hilltops (16%), as well as in transitional (21%), hillbase (16%), and Sabal (5%) forests in upland swales. Because passage through scrub swamp vegetation was difficult and time-consuming, we dedicated less effort to monitoring this forest type. Although we did not observe Crane Hawks hunting in the lowest-lying scrub swamp forests, this may not indicate that they avoided this habitat.
Flexibility of the Intertarsal Joint
We trapped three adult and two immature Crane Hawks, and measured backwards flexion and lateral and medial swing of their intertarsal joints. Backward flexion was 34 ± 8° (range 20–40°), lateral swing was 16 ± 6° (range 13–25°), and medial swing for all birds was 15°.
Food-Niche Breadth and Overlap
During research at Tikal, food habits of Great Black-Hawks (Buteogallus urubitinga), White Hawks (Leucopternis albicollis), Collared Forest-Falcons (Micrastur semitorquatus), Laughing Falcons (Herpetotheres cachinnans), Hook-billed Kites (Chondrohierax uncinatus), Roadside Hawks (Buteo magnirostris), and Bicolored Hawks (Accipiter bicolor) have been studied. Based on these data and our present results for Crane Hawks, we calculated food-niche breadth and overlap at the level of prey “type” (Table 2). Laughing Falcons, Bicolored Hawks, and Hook-billed Kites were dietary specialists (one prey type comprised >90% of diet), and had low values of food-niche breadth, whereas White Hawks, Crane Hawks, Collared Forest-Falcons, Roadside Hawks, and Great Black-Hawks had less specialized diets and intermediate to high food-niche breadth (Table 2). Food-niche overlap between these raptors and Crane Hawks ranged from very low (Laughing Falcon, 0.03) to relatively high (Roadside Hawk, 0.76).
Food-niche breadth and overlap index values based on raptor diets studied at Tikal National Park, Guatemala. Indices are based on identified prey items (n)
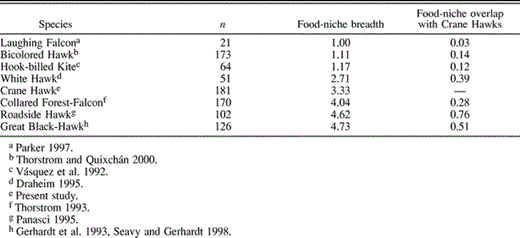
Food-niche breadth and overlap index values based on raptor diets studied at Tikal National Park, Guatemala. Indices are based on identified prey items (n)
Discussion
Crane Hawks have been reported to take a broad assortment of prey including large insects and other arthropods, lizards, snakes, frogs, rodents, nestling birds, bats, and snails (Smithe 1966, Alvarez del Toro 1980, del Hoyo et al. 1994). With the notable exception of invertebrates, the breeding diet of Crane Hawks at Tikal consisted of prey similar to those previously reported (Table 1). Nocturnal animals including rodents, bats, some frogs, a skunk, and a snake made up a significant proportion of Crane Hawk diets, even though all prey apparently were captured during daylight hours. Invertebrates were absent from Crane Hawk diets at the nest, and it is unlikely that insects or other arthropods accounted for a large percentage of the unidentified prey items because we regularly identified these invertebrates during observations of other nesting raptors at Tikal. It is possible that invertebrates were eaten by Crane Hawks away from the nests, but, because of the low energetic reward relative to the cost of transport, were not used for provisioning nestlings.
Of the several hunting methods attributed to Crane Hawks, we observed most but not all at Tikal. Perch-hunting, no doubt the most common mode of hunting among buteonine hawks, has only occasionally been reported in Crane Hawks (Sutton 1954), and comprised only 16% of the hunting incidents we observed. Crane Hawks also have been observed hunting on the wing, coursing over open areas in harrier-like fashion (Sutton 1954, Olmos 1990), sometimes behind fire lines in burning pastures (Dickey and van Rossem 1938). We did not observe this behavior in our forested study area.
The most frequently reported Crane Hawk foraging behavior is the search-and-probe hunting technique, in which they use their long, “double-jointed” legs, and less often their heads, to actively search within potential prey refugia in trees and on the ground (Sutton 1954, Jehl 1968). Crane Hawks have been observed clinging to tree trunks in a variety of positions, including upside down, probing their head and feet into holes and behind bark while using their tail and wings to maintain balance (Sutton 1954, Haverschmidt 1962, Jehl 1968). On the ground, Crane Hawks have been observed poking their legs between rocks in fallow fields (Sutton 1954), and searching along stream banks (Wetmore 1965). We observed all of these features of search-and-probe foraging at Tikal, and this strategy comprised 63% of observed hunting incidents.
Nocturnal prey, along with many reclusive diurnal prey, most likely were captured and extracted from their daytime retreats via the search-and-probe hunting technique. During daylight, rodents and skunks seek shelter in dens on the ground or in tree cavities (Emmons 1990), and bats roost in holes, among foliage, behind bark, and under hanging palm fronds (Altringham 1996). Several species of tree frogs (all nocturnal) are known to breed and hide in bromeliads and the leaf axils of palm trees (Meyer and Foster 1996), and one species of obligate bromeliad-breeding tree frog has been reported as Crane Hawk prey in southern Brazil (Bokermann 1978). The one snake among prey identified to species, the northern cat-eyed snake (Leptodeira septentrionalis), is a nocturnal, arboreal frog predator that occasionally searches bromeliads for prey and sometimes hides in them during the day (Greene 1997). We hypothesize that Crane Hawks utilize a search image focused on potential hiding places of prey rather than on particular prey types themselves.
Nesting birds may also have been captured using the search-and-probe technique. Most birds captured were small and included adult passerines and several nestling parrots. However, most adult birds are probably too elusive for Crane Hawks to capture unless encountered while incubating. In Venezuela, Mader (1981) observed a Crane Hawk using one foot to hang beneath palm fronds while grabbing at a hanging bird nest with the other. We speculate that Crane Hawks often attempt to capture nestlings and/or incubating adult birds by reaching within nests and potential nest cavities.
Crane Hawks have been observed chasing prey through branches and vines on foot (Friedmann and Smith 1950), and we suggest that other diurnal prey, such as lizards, snakes, birds, and some frogs are encountered, flushed, and sometimes captured by Crane Hawks in the course of search-and-probe foraging. On various occasions, we observed Crane Hawks chase what we believed to be lizards on the trunks and branches of trees, and arboreal Norops lizards were frequent in Crane Hawk diets.
The search-and-probe hunting technique of Crane Hawks appears to be facilitated by several unique anatomical features. Crane Hawks have an intertarsal joint that flexes both forward and backward and has a high range of lateral mobility (>30°; Burton 1978, present study). In addition, Crane Hawk tarsal scales are fused into a few large plates, presenting an almost smooth surface (Friedmann 1950, Sutton 1954). This feature may prevent these scales from catching when the hawk reaches into constricted spaces. Crane Hawks also have disproportionately small outer toes (Friedmann 1950, Wetmore 1965), which may enable them to extract prey from narrow spaces where a larger clenched foot could not pass.
Diet and hunting behavior are basic to any understanding of community structure, and raptor communities have inspired some research on community trophic relations (Marti et al. 1993). Our analysis of food habits of similarly sized raptor species at Tikal suggests some ways in which differences in prey selection and hunting behavior may facilitate coexistence of Crane Hawks and these other raptors over large geographic areas. We arbitrarily distinguished three categories of food-niche overlap: low (0.00–0.33), moderate (0.34–0.66), and high (0.67–1.00). Hook-billed Kites, Laughing Falcons, and Bicolored Hawks are dietary specialists taking arboreal snails, snakes, and birds, respectively (Vásquez et al. 1992, Parker 1997, Thorstrom and Quixchán 2000). Because these prey were either absent or comprised only a small fraction of Crane Hawk diets, food-niche overlap between these species and Crane Hawks was low.
Collared Forest-Falcons mainly perch-hunt, detecting active, mobile prey rather than hidden, nocturnal prey, and like Crane Hawks ate mammals but also took numerous birds (Thorstrom 1993). Although Collared Forest-Falcons and Crane Hawks displayed high dietary overlap when values were calculated at the Class level (0.85), overlap dropped considerably (to 0.28) when prey types were distinguished to more precise taxonomic levels (i.e., rats and mice vs. squirrels).
The uncommon Great Black-Hawk and the ubiquitous Roadside Hawk are reported to be dietary generalists and opportunistic hunters (Robinson 1994, Panasci 1995, Seavy and Gerhardt 1998). The wide variety and relatively equal proportion of prey types in their diets explains their high food-niche breadth indices, whereas the intermediate food-niche breadth indices of Crane Hawks and White Hawks reflect the dominance of several prey types among a wide variety of prey in their diets. All four of these raptors take many similar prey types. Among these species, dietary overlap between Crane Hawks and White Hawks was lowest. White Hawks were mainly perch-hunters frequenting treefall gaps more often than expected, presumably taking active prey; also, snakes dominated White Hawk diets whereas few mammals and amphibians were taken (Draheim 1995). Great Black-Hawks took proportionally more lizards, snakes, and birds, and fewer rodents and frogs than did Crane Hawks (Seavy and Gerhardt 1998). Like Crane Hawks, Great Black-Hawks utilized a broad range of habitats, and have been observed using their long legs to search for prey (del Hoyo et al. 1994); however, dietary overlap between these two raptors is probably minimized by differences in prey size, with Crane Hawks taking smaller prey. Crane Hawks showed highest dietary overlap with Roadside Hawks, reflecting the fact that both species preyed largely on rodents and lizards; however, insects, which were not observed in Crane Hawk diets in our study, were frequent in Roadside Hawk diets (18% of identified prey items; Panasci 1995).
Crane Hawks appear to be opportunistic hunters capable of using a variety of hunting methods. The frequently observed use of the search-and-probe foraging technique and the predominance of nocturnal animals in Crane Hawk diets suggest these raptors encountered many prey by inspecting potential prey refugia. Among Tikal's raptors, such heavy reliance on nocturnal prey is shared mainly by the Crested Eagle (Morphnus guianensis) and the Black Hawk-Eagle (Spizaetus tyrannus; Whitacre et al., unpubl. data), neither of which possess the anatomical adaptations noted in Crane Hawks. Although several similarly sized sympatric raptors at Tikal also exhibited rather broad diets, the unusual hunting technique used by Crane Hawks probably enables them to capitalize on food resources largely unavailable to the others, thus minimizing dietary overlap.
Acknowledgments
This is a contribution of the “Maya Project,” a conservation research effort of The Peregrine Fund; we thank the individuals, trusts, and foundations that support this organization. We also thank A. Rosales, J. and G. López A., C. Marroquín V., M. Córdova A., E. Martínez R., B. González C., A. Enamorado G., L. E. Jones, Kia Belice Sutter, J. Waddell, and J. Madrid M. for assisting with data collection. We thank the directors and staff of Tikal National Park for their support and cooperation. We are indebted to Mario Jolón for conducting a rodent identification workshop and providing us with additional information and assistance, and H. W. Greene for identifying Leptodeira septentrionalis and providing insight into tropical predator assemblages. M. J. Bechard, S. J. Novak, R. P. Gerhardt, L. Kiff, and several anonymous reviewers made helpful suggestions on the manuscript.
Literature Cited
Author notes
Present address: Owyhee Canyonlands Research, P.O. Box 582, Caldwell, ID 83606, [email protected]
