





Abstract
We studied the ecological distribution and foraging behavior of three species in the Black-throated Green Warbler (Dendroica virens) species group in the Eastern Highlands of Chiapas, Mexico. The three species, Black-throated Green, Hermit (D. occidentalis) and Townsend's (D. townsendi) Warblers are largely allopatric on the breeding grounds and variably sympatric during the winter. We surveyed the composition of 166 mixed-species flocks and recorded habitat variables associated with the flock location along an elevational transect. We found that Black-throated Green and Townsend's Warblers segregated by altitude, whereas Hermit Warbler overlapped extensively with both other species. Even though Townsend's and Hermit Warblers co-occurred commonly in the same flocks, the latter showed strong selective use of pine trees and the former showed weaker selectivity of oak trees. Black-throated Green Warbler showed no tree-type selection. The combined differences in elevational distribution and tree-type use resulted in very low ecological overlap for the three species. The Black-throated Green Warbler was the most distinct in its foraging behavior, occurring more often in the outer and upper branches of trees, and using hanging, aerial, and hovering maneuvers more than its congeners. In addition, it showed a higher movement frequency and a lower rate of between-branch flights. The Black-throated Green Warbler also is the most morphologically distinct as a result of its overall smaller body size, which is consistent for the trend towards larger body size in more conifer-specialized species in this genus. We propose that as new breeding populations were founded, and speciation occurred, individuals of the more derived species occupied decreasingly productive winter habitats.
Introduction
Closely related species of migratory land birds show a strong tendency to winter allopatrically (Salomenson 1955, Chipley 1980, Greenberg 1986), even when breeding sympatrically. One marked exception is the Black-throated Green Warbler (Dendroica virens) complex. These five species show marked allopatric breeding distribution, yet extensive winter sympatry: four species can be found wintering in similar habitats in a broad area of Mesoamerica from Central Mexico through Nicaragua.
The allopatric breeding distribution is probably a result of the recent speciation in the complex (Mengel 1964, Bermingham et al. 1992). Mengel (1964) hypothesized that the species in the complex originated from events occurring throughout the Pleistocene, and recent analysis of mtDNA suggest that recent vicariant events may be responsible for the formation of at least the Black-throated Green/Hermit (D. occidentalis)/Townsend's (D. townsendi) Warbler lineage. Molecular data suggest that the Hermit and Townsend's Warblers are the most closely related species (hybrids are frequent, Morrison and Hardy 1983, Rowher and Wood 1998) in the complex (Bermingham et al. 1992).
In this paper we examine the ecological segregation in the Black-throated Green Warbler complex during the winter in Mexico. All three species commonly occur in pine-oak woodlands, but Black-throated Green Warbler also is found in subtropical forest on the Caribbean slopes of northern Mesoamerica and in high-elevation zones of Costa Rica and Panama. Within the pine-oak belt of northern Mesoamerica, the Hermit Warbler has been noted to occur primarily in pines (Hutto 1980, 1985), whereas the Townsend's Warbler shows little specialization, occurring in both forest and secondary habitats. From current published accounts it is difficult to tease apart broad geographic patterns of occurrence from actual habitat selection based on locally available choices facing the settling warblers. In this study we investigated the pattern of habitat use and details of foraging behavior in a small region of distribution overlap to determine whether the different species show complementary ecological specialization.
Methods
Study Sites
The study was conducted January–March 1993 in the highlands of Chiapas, Mexico, from Naja and Ocosingo (17.04°N, 92.15°W, elev. 920 m) to San Cristobal de las Casas (16.45°N, 92.40°W, elev. 2,300 m). The sharp transition between Caribbean slope and montane forest of eastern Chiapas offers an opportunity to examine the details of species' distributions across a local habitat gradient. Rainfall at the eastern end of the gradient (Ocosingo) averages 1.8 m year−1 with a distinct dry season between March and June; the western site (San Cristobal) receives less rain (12 m year−1, Garcia 1988), but considerable moisture from fog. The dominant mature vegetation along the entire transect is pine-oak woodland (Breedlove 1981), reaching a canopy height of approximately 15–20 m with different mixtures of perennial (live) oaks (Quercus spp.) and medium to long-needled pines (Pinus spp.).
Mixed-Species Flock Surveys
For species that are dependent upon mixed-species flocks, an alternative to fixed-plot, point, or transect sampling is to census the composition of mixed flocks (see Robbins et al. 1992). This is the method we used to quantify the distribution of Dendroica. We surveyed 166 mixed-species flocks between Ocosingo and San Cristobal de las Casas. We located flocks in tracts of pine-oak woodland accessible from roads, dividing our effort between low (800–1,500 m) middle (1,500–2,000 m) and high (>2,000 m) elevation sites. We attempted to balance the number of flocks located in each zone (n = 60, 54, 52, respectively). We surveyed each flock for an average of 24 min (range = 15–50 min), and we recorded the number (and when possible the sex and age class) of each species of Dendroica, the percentage of pines, oaks, and other broad-leafed trees within an estimated 25-m-radius circle around the center of the flock, the dominant canopy height, the height of the canopy and subcanopy formed by pine, oak, or broad-leafed trees, and elevation. For consistency, the same observer made all estimates. The type of tree (i.e., oak, pine, broad-leafed, acacia, etc.) was recorded when possible for the first observation of an individual in a flock.
We separated flocks into three overlapping groups. Each group consisted of those flocks containing at least one individual of a given species, which sometimes resulted in the same flocks being included in the different groups. This imposes the appropriate bias on interspecific habitat use against finding a significant difference where flock overlap is high. We calculated the mean and standard error for each of the habitat variables for each of the species, weighted by the number of individuals of a species in that flock. These values were subjected to a one-way MANOVA to detect interspecific differences in habitat use. Proportional variables were arcsine transformed. Post-hoc, pairwise species comparisons using Tukey's HSD Test (Statsoft 1995) were conducted for all significant variables in the model. A forward, stepwise discriminant function analysis (DFA) was then conducted to determine the overall Mahalanobis' distances between species' centroids and the important variables contributing to the two discriminant function factors.
Foraging Behavior
C. E. Gonzales made all of the foraging observations by following 212 individuals of the three species; 90, 70, and 57 each for Townsend's, Hermit, and Black-throated Green Warblers, respectively. She selected the first individual encountered in a mixed-species flock and recorded its movements, and estimated movement distances making a contemporaneous record using a cassette tape recorder. Activities were timed in the laboratory with a stopwatch. Foraging variables were categorized into two types: locational, and a combination of locomotory and prey- attack behaviors. Locomotory variables included hop, short flight, switching (bird moves along branch leaning over in successive opposite directions), and potential attack behaviors including standing glean (standing and grabbing prey with bill), aerial maneuvers (lunges and upward strikes), hover (stationary beating of wings at the end of an aerial maneuver), and hanging. Some foraging behaviors are indistinguishable from search behaviors and so total foraging attack-rate is overestimated from these data. Successful foraging was recorded when a prey was observed or the warbler followed a foraging attack with a bill wipe. Locational variables include the height of the bird above the ground, the relative height of the bird (bird height/tree height), the number of visits to epiphytic plants, and whether the bird was in the inner or outer 50% of the branch on which it was foraging. For statistical analysis, we used only one value per individual watched, and the sample size is based on the number of individuals. Behavioral data were transformed to rates (per min). We examined interspecific differences in foraging variables with a stepwise DFA.
Results
Flock Overlap
We observed a total of 562 of the three focal species of warblers in 166 mixed-species flocks between Ocosingo and San Cristobal de las Casas. These individuals comprised 17% of the birds seen in insectivorous mixed-species flocks in the study area. We observed Black-throated Green Warblers in 66%, Hermit Warblers in 45%, and Townsend's Warblers in 34% of the flocks. All flocks had at least one Dendroica species, 50% of the flocks had only one, 46% had two, and only 2% had three species. Based on expected values generated from the joint probabilities of co-occurrence, we determined that the number of species per flock was significantly different from a random distribution (χ23 = 59.3, P < 0.001). In particular, flocks with no species, two species, and three species were underrepresented.
The abundances of Townsend's and Black-throated Green Warblers were significantly negatively correlated (rs = −0.52, P < 0.01), as were those of Hermit and Townsend's Warblers (rs = −0.25, P < 0.05). In contrast, the abundances of Townsend's and Hermit Warblers were positively correlated (rs = 0.71, P < 0.01). The degree of flock overlap (the sum of the minimum value of the standardized proportion of individuals of each species in a flock) between Hermit and Townsend's Warblers was the highest of the pairwise comparisons (0.52), whereas it was 0.33 for the Hermit/Black-throated Green Warbler comparison, and only 0.06 for Townsend's/Black-throated Green Warbler.
Flock Habitat
The overall MANOVA model (species versus habitat variables) was significant (Wilks' Lambda = 0.41, P < 0.001). The three species showed significant interspecific variation in six of the habitat variables (Table 1 presents significant post-hoc comparisons). The DFA also was significant (Wilks' Lambda = 0.31, F10,602 = 47.8, P < 0.01). The first factor had a canonical correlation coefficient of 0.82, and the only important variable was elevation (loading = 0.95). The second factor had a correlation coefficient of 0.25, and two important variables (oak canopy height = 0.66 and percentage pine = 0.87). Based on the DFA, elevation was the variable producing the greatest segregation between species, and tree-canopy composition contributed less to the overall segregation. The partial Wilks' Lambda for elevation was 0.31 and for other variables was greater than 0.9. The overall Mahalanobis' distance was greatest between Townsend's and Black-throated Green Warblers (10.7), smallest between Townsend's and Hermit Warblers (1.57), and intermediate for the Hermit and Black-throated Green Warblers (5.05).
Means ± SD for habitat variables estimated at the location of 166 mixed-species flocks and weighted by the abundance of each Dendroica species between Ocosingo and San Cristobal de las Casas, Chiapas, Mexico. Significant comparisons are based on Tukey's post-hoc tests for variables with overall significant interspecific variation based on an ANOVA

Means ± SD for habitat variables estimated at the location of 166 mixed-species flocks and weighted by the abundance of each Dendroica species between Ocosingo and San Cristobal de las Casas, Chiapas, Mexico. Significant comparisons are based on Tukey's post-hoc tests for variables with overall significant interspecific variation based on an ANOVA
With respect to elevation, Townsend's and Hermit Warblers had similar mean values, both occurring primarily in high-elevation flocks. However, Hermit Warbler had the greatest variation in elevation of occurrence with a coefficient of variation for this variable approximately double the other species (CVs = 17%, 13%, and 30% in Black-throated Green, Townsend's and Hermit Warblers, respectively).
Tree Use Within Flocks
The three species showed distinct differences in the types of trees in which they were observed. Black-throated Green Warbler was the most generalized (Fig. 1); Townsend's Warbler occurred most frequently in oaks (60%), and Hermit Warbler specialized on pines (97%). We tested these values using the mean canopy cover for the different species derived in the habitat analysis as an expected value. Black-throated Green Warbler was not significantly different in its tree use when compared to tree cover (χ22 = 0.11). However, Townsend's Warbler was found significantly more often in oaks than expected (χ22 = 19.5, P < 0.001), and Hermit Warbler significantly more often in pines (χ22 = 28.5, P < 0.001). Percentage overlap in tree use (as in flock occurrence) was 0.81 for Townsend's and Black-throated Green Warblers, 0.36 for Townsend's and Hermit Warblers, and 0.43 for Hermit and Black-throated Green Warblers.

Percentage use of oaks and pines by different species of Dendroica based on location of first observation of individuals, compared to the expected value based on the average percentage of canopy volume occupied by tree types within the habitat sampling area for each flock (weighted by the abundance of a particular species in the flock). BTG = Black-throated Green Warbler
Foraging Behavior
Foraging rate (number of successful foraging maneuvers per min) was not significantly different between the three species (3.9–4.5 min−1). A DFA on foraging behavior variables also produced two significant factors (Wilks' Lambda = 0.52, F18,412 = 9.1, P < 0.001). Variables made relatively equal contributions to the discrimination with partial Wilks' Lambdas ranging from 0.83–0.98. The overall Mahalanobis' distance was much larger for the Townsend's/Black-throated Green (3.41) and Hermit/Black-throated Green (3.37) than the Townsend's/Hermit (0.74) comparison. Black-throated Green Warbler was distinctly different from the other two species in occurring in the upper and outer branches of trees and using a high frequency of hops, aerial maneuvers, hovers, and switching behavior, and a lower frequency of inter-branch flights (Table 2). We did not include epiphyte visitation in the DFA (because epiphytes are far more common at higher elevations); Hermit and Townsend's Warblers visited epiphytic plants far more frequently than Black-throated Green Warbler (Table 2).
Mean number of events min−1 (± SD) for foraging behaviors and location of three species of Dendroica in Chiapas, Mexico
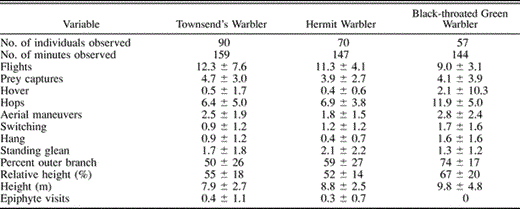
Mean number of events min−1 (± SD) for foraging behaviors and location of three species of Dendroica in Chiapas, Mexico
Discussion
The Pattern of Ecological Segregation
The three species showed distinct ecological segregation. The Black-throated Green Warbler was largely separated by elevation from the two western species, particularly from the Townsend's Warbler; Hermit Warbler was most common at higher elevations but had a broader elevational distribution than the other two species. Hermit and Townsend's Warblers co-occurred commonly in mixed-species flocks and, to a lesser extent, so did Hermit and Black-throated Green Warblers. However, Black-throated Green and Townsend's warblers rarely co-occurred. The three species had distinct patterns of occurrence in pine and oaks. Hermit Warbler was a pine specialist, Townsend's Warblers preferred oaks, and Black-throated Green Warblers occurred in tree types in proportion to their abundance in the habitat. The combined overlap derived from species co-occurrence in flocks and overlap in tree use (the product of the two, Cody 1974) was very low (0.06) for Black-throated Green and Townsend's Warblers, and low for Hermit/Townsend's (0.19) and Hermit/Black-throated Green Warbler (0.14) comparisons.
Hermit and Townsend's Warblers were very similar in the details of their foraging behavior and were distinct from Black-throated Green Warblers in using fewer hops while searching, more (shorter) flights between branches, and fewer lunges, upward strikes, hanging behaviors, and hovers (see also Rabenold 1980).
The overall morphology of these birds is most similar for the western species (Rising 1988). Specifically, the western species are significantly larger than the Black-throated Green Warbler. The evolution of larger body size in the western species is consistent with the generalization derived from eastern Dendroica (Greenberg 1979), where conifer specialist species are larger than species that occur in broad-leafed or both broad-leafed and coniferous habitats. Townsend's and Hermit Warblers are more strictly associated with pure or near-pure stands of conifers than the smaller Black-throated Green and Black-throated Gray Warblers (Morrison 1982, Parrish 1995, Pearson 1997, Wright et al. 1998). Greenberg (1979) hypothesized that foraging in the interior of large coniferous trees requires less agility, and hence selection for small body size should be weaker than for active foragers in broad-leafed vegetation.
Evolution of Habitat Segregation
Despite differences in the proposed scenarios for speciation in this group, it is generally agreed that western species originated from one or more invasions from an eastern boreal population (Mengel 1964, Bermingham et al. 1992). The following scenario is based on three assumptions: (1) colonization proceeded from east to west, (2) large changes in ecology or morphology occur in small colonizing populations, because of the lack of genetic structuring or geographically based morphological variation in migratory wood warblers, and (3) the expansion of breeding distribution requires similar expansion into new regions or habitats on the wintering range (Salomenson 1955, Cox 1968).
Step 1. The proto-Black-throated Green Warbler occupied Appalachian and eastern boreal forests of North America. During an interglacial period, small populations colonized the northern Rocky Mountains. These rapidly expanding populations extended their wintering range, without greatly increasing migration distance, by migrating to coniferous and mixed forests of western Mexico. As breeding populations expanded, birds wintered farther south, coming into contact with the proto-Black-throated Green Warbler populations wintering primarily on the lower Caribbean slopes of Mesoamerica. Adaptation to more montane and more coniferous habitats included the evolution of larger body size (Greenberg 1979) and less agile (i.e., less aerial) foraging behavior.
Step 2. During a more recent interglacial period, expanding populations colonized the Sierra and Cascade Ranges. Because existing populations in the complex wintered abundantly throughout northern Mesoamerica, new winter habitat could only be obtained by specializing on habitats under-utilized by existing populations. During the period of small population sizes, habitat preference for pine (a habitat less preferred by proto-Townsend's Warblers) was rapidly established as it allowed for reduced competition with existing populations.
Step 3. Behavioral specialization on pine allowed Hermit Warblers to expand into areas occupied by Black-throated Green Warblers, which use a variety of broad-leafed habitats within their winter range.
We can further expand this scenario for the entire species complex by incorporating what is known from the published literature of the Black-throated Gray Warbler. Black-throated Gray Warblers probably split from the Black-throated Green lineage earlier than the other western species (Mengel 1964, Bermingham et al. 1992); it is small-bodied and morphologically similar to Black-throated Green Warbler, occupying mixed oak-conifer and oak woodlands during the breeding season and occurring in lowland oak and broad-leafed habitats during the winter in western Mexico (Hutto 1980). Therefore, it appears that the proto-Black-throated Gray Warbler expanded into mixed habitats and occupied the western lowlands, leaving montane coniferous breeding habitats and highland wintering habitats to the later-colonizing western species. It is worth noting that the Golden-cheeked Warbler (Dendroica chrysoparia) occurs at elevations similar to Townsend's Warbler during the winter and, like the Townsend's, prefers to forage in oaks (Rappole et al. 1999). The small population size may allow this extensive ecological overlap.
Ecological “Leap Frog” Migration
If competition underlies the colonization of new winter ranges or habitats as breeding populations expand, new populations should be at a competitive disadvantage (or at best no advantage) over individuals in existing populations. Based on this we should find that birds in expanding breeding populations occupy either more distant or otherwise poorer winter sites (Salomenson 1955, Greenberg 1980). This is recognizable as a more general statement of the “leap frog” migration pattern (Bell 1996). There is some prima facie evidence in support for the latter phenomenon within the Black-throated Green species group: Black-throated Green Warblers occupy subtropical woodlands, which generally have higher insect abundances than temperate woodlands as one moves up tropical mountains (Janzen 1973, 1980). Furthermore, Hermit Warblers occupy pine habitats which have lower arthropod abundances than the oak habitats preferred by Townsend's Warblers in the highlands of Mexico (Hutto 1980). Consistent with this hypothesis, that Hermit Warbler has been relegated to the least productive habitat, is the observation that Townsend's Warblers are dominant to Hermit Warblers on the breeding grounds (Rowher and Wood 1998). In addition, Hermit Warblers are very similar in their structural morphology and foraging behavior to Townsend's Warblers. It is the lack of any clear divergence in preference for pines or conifers in general on the breeding grounds, the large preference for pines versus oaks in the winter, and the lack of apparent morphological or behavioral specialization that leads us to suggest that winter competition is the primary factor selecting for the preference for pine during the nonbreeding season in Hermit Warblers.
Acknowledgments
We thank A. Cruz, H. Flores, C. Macias, and J. Salgado for assistance with data gathering. Kevin Omland provided criticisms of an earlier draft. Fieldwork was supported by grants from the National Geographic Society and the Scholarly Studies program of the Smithsonian Institution.
Literature Cited

{kind=link}