





Abstract
We studied the context of brood reduction through infanticide by communally breeding Guira Cuckoos (Guira guira) in central Brazil. During seven reproductive seasons, we monitored 142 nests from egg laying until fledging. Almost all nests (97%) lost eggs through ejection, and chick deaths occurred in 72% of all nests with hatchlings. There was evidence for infanticide in 38% of the nests that exhibited some mortality. We compared egg and chick mortality in the early part of the season with the later part, when insect abundance declines, but found no significant differences. Less than one-third of all nests monitored showed asynchronous hatching of eggs, and in those that did, chick death was not in reverse hatch order. Although there are several plausible explanations for infanticide, we highlight one likely candidate, which is its interpretation as a sexually selected trait where individuals gain reproductive benefits by provoking the group's nesting failure.
Infanticide is a widespread phenomenon among animals of various taxa (Hrdy 1979, Hrdy and Hausfater 1984). One of the prevailing hypotheses to explain this behavior suggests that it may be practiced by non-parental adults to provide them with a breeding opportunity. Termed sexually selected infanticide, this behavior results from competition among individuals of the same sex, enabling some to breed and thereby increase their reproductive success relative to others (Trivers 1972, Hrdy 1979). Sexually selected infanticide in birds has been only sporadically reported, most frequently in cooperative or colonial species, and seldom for territorial monogamous birds (Vehrencamp 1977, Bertram 1979, Trail et al. 1981, Crook and Shields 1985, Freed 1986).
Infanticide is a common event for the communally breeding Guira Cuckoo (Guira guira), a species with an extensive geographical range in South America (Sick 1997, Macedo and Melo 1999), and with a poly-gynandrous mating system (Quinn et al. 1994). The lengthy breeding season (August to March) may allow as many as five successive nestings for groups (up to 13 adults; modal group size of 6) in the same territory. The failure of a nesting bout allows renesting of a group within a much shorter interval than if the nesting is successful (30 days earlier, on average; Macedo 1994). Communal clutches can be very large, with some cases of up to 24 eggs laid in the nest by different females. However, the clutches are reduced due to egg ejection by group members (Macedo 1992). The hatched brood is then further reduced by predation, disease, and other causes, as well as by the frequent practice of infanticide (Macedo and Melo 1999). Guira Cuckoo group members do not share equitably in feeding and guarding nestlings, but whatever food they provide is distributed randomly among chicks, with no evidence of recognition of their own offspring by different adults (Macedo 1994).
In a previous paper (Macedo and Melo 1999) we confirmed the occurrence of infanticide in this species, and here we present additional quantification of its context and frequency, which includes the degree and seasonality of hatching asynchrony, mortality of different-aged chicks, seasonal shifts in egg ejection and infanticide, and degree of mortality in different-sized broods.
To date, we have found no evidence for cannibalism. The young also cannot compete directly for resources, since they are limited to the nest until fledging and are later fed by adults for several weeks. The possibility that infanticide is exercised selectively (progeny choice) also is unlikely, for reasons stated previously. Thus, in this study, we considered primarily whether Guira Cuckoo nestling mortality could be interpreted as sexually selected infanticide. The most direct evidence to support a conclusion of sexually selected infanticide would be to establish the parentage of victims and killers. Because of the difficulty of capturing adults, we evaluated the sexual selection hypothesis indirectly, by determining the seasonality of egg eviction and infanticide. An early seasonal bias in these parameters would favor the explanation of sexually selected infanticide, because nest failure early in the season would allow the group to renest faster, providing infanticidal adults with a new reproductive opportunity.
Methods
We used reproductive data from seven seasons (1987, 1988, 1990, 1994, 1997, 1998 and 1999) conducted in the area surrounding Brasilia, Brazil (15°47′S, 47°56′W). The field site (about 30 km2), composed of native savanna vegetation, urbanized plots, and cultivated fields, included the breeding territories of 25 groups in 1987 (with 33 nesting bouts), 22 groups in 1988 (28 nesting bouts), 16 groups in 1990 (20 nesting bouts), 12 groups in 1994 (18 nesting bouts), 17 groups in 1997 (25 nesting bouts), 13 groups in 1998 (31 nesting bouts), and 10 groups in 1999 (16 nesting bouts). Limited banding data indicate groups used the same territories throughout a season, and group composition remained relatively stable (sometimes between seasons).
Reproductive activity is mostly restricted to the rainy season, which occurs from approximately mid-August to mid-March. However, peak activity in our study area occurs in September and October, and most of our data were collected from August to December. We recorded nest contents every day and searched the ground beneath each nest tree for remains of any tossed eggs. With the exception of the 1994 and 1997 seasons, we numbered eggs sequentially as they were laid, to verify the laying and tossing patterns. Groups deserted some nests without incubating, usually after a number of eggs had been laid and tossed. In some nests, laying occurred during a protracted period with many eggs eliminated through tossing (in one exceptional situation a group laid and tossed 24 eggs during 11 days).
The number of eggs laid by groups varied widely, and because desertion occurs at unpredictable times during the laying phase, we report the mean clutch size only for those cases where incubation was initiated, because this affords some certainty that the clutch was completed. Clutches of only one or two eggs were never incubated. Thus, incubated clutches ranged in size from 4 to 21 eggs. Adults in the vicinity of the nesting tree were counted at each nest-check to determine group size; at least three consistent maximum-number counts in different days were needed before group size was determined. We minimized visits during the incubation period, but once hatching initiated, daily visits were resumed to check on nestling presence. Whenever possible, chicks were individually marked with colored dye soon after hatching until unique combinations of colored leg bands could be applied. Until fledging (approximately 12 days), nestling deaths were recorded whenever a chick disappeared, was observed being ejected or carried away from the nest by an adult, or when its body was found.
At 21 nests we monitored adult behaviors, including nestling feeding, ejection of chicks from the nest, and presence near the nesting tree (radius of about 10 m). Observations were carried out 15 to 20 m from the nesting tree for several hours each day (ranging from 4 to 10 continuous hours between 08:00 and 18:00), and usually began soon after the first chick hatched and continued for at least one week, when the majority of infanticides occur (Macedo and Melo 1999). During four field seasons, we attempted to capture adult group members by using a hand-reared lure bird, either: (1) surrounded by mats of monofilament nylon nooses; or (2) inside a fall trap, operated manually at a distance. Although these were the best methods we have found yet to capture birds, the success rate was limited, resulting in a small sample across all field seasons (n = 105 adults), representing approximately 21% of the total population under study. Captured birds were banded with unique combinations of colored leg bands and marked with dye to facilitate quick identification.
We used two-tailed t-tests to compare independent samples of clutch sizes and percent of hatched eggs in synchronous versus asynchronous broods. Chi-square analyses were used to test for differences in nests fledging full versus reduced broods in these categories. We conducted correlation analyses to determine the association between brood size and brood mortality within group-size categories, and performed simple linear regressions of group size on several reproductive parameters. Means are given with standard errors, and significance level was set at P = 0.05. All statistical analyses were run using SYSTAT software (SYSTAT, Inc. 1992).
Results
Mortality Patterns and Documentation of Infanticide
We found 171 active nests during the study period, of which 142 were monitored (Table 1). Almost all nests lost a few eggs (97% of 142 nests), whether or not the clutch was then completed. The number of eggs lost varied widely, from zero to as many as 24 eggs. On average, broods were smaller than clutches (Table 1), and were sometimes further reduced through infanticide. In the 42 nests that suffered brood mortality (72% of all nests with chicks), chick loss averaged 2.6 ± 0.3 (range: 0–8 nestlings nest−1) and was classified into two categories, partial or complete brood loss.
Summary of reproductive characteristics and mortality patterns for Guira Cuckoos in seven reproductive seasons
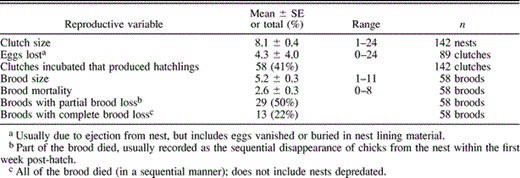
Summary of reproductive characteristics and mortality patterns for Guira Cuckoos in seven reproductive seasons
To discern whether there might be a seasonal bias in either egg or chick mortality, we divided the season into “early” (July–Oct) and “late” (Nov–Mar) phases. We found no difference in the percentage of eggs tossed in 113 “early” versus 29 “late” clutches (Mann-Whitney U = 1,696, P = 0.77), or in the percent brood mortality between 48 “early” versus 14 “late” broods (Mann-Whitney U = 242.5, P = 0.11).
In one group we were able to capture and mark one adult that committed infanticide (details on infanticides in Macedo and Melo 1999). The group (four or five members) had four nesting bouts in the season, the first one ending in desertion of the nest. In the three subsequent nestings, a total of 14 chicks were produced, although only five fledged. In these repeated nestings we observed two infanticides, one by the marked adult. This adult fed chicks in at least two of the nesting bouts: of the 221 feedings observed in both nests, it performed approximately 16% of the feeds, and continued delivering food to nestlings after killing the chick.
Hatching Asynchrony and Chick Mortality
Of the nests where hatching dates could be ascertained for all chicks (n = 34), less than one-third hatched asynchronously, over a mean period of 2.4 days (range 2–4). Synchronous nests (all hatchings within 24 hr, n = 24) did not differ significantly from asynchronous ones (Table 2) in: the size of clutches incubated (t9 = −2.1, P = 0.07); the percentage of eggs that hatched (t9 = −1.4, P = 0.19); the number of nests with full broods versus those with partial or complete brood losses (χ21 = 0.72, P = 0.64); and the percent mortality of broods (t7 = −0.4, P = 0.73). In addition to hatching asynchrony not being typical, the frequency or degree of asynchrony did not increase later in the season. Nine of the 10 asynchronous nests started their hatching during the first half of the rainy season, between mid September and the end of November. Only one nest hatched asynchronously at the end of the rainy season, in February.
Nesting characteristics and chick mortality patterns in synchronously and asynchronously hatching broods of Guira Cuckoos. Sample sizes reflect only nondepredated nests for which hatching dates of all chicks were known
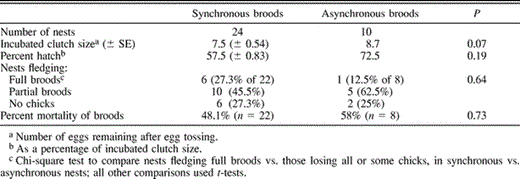
Nesting characteristics and chick mortality patterns in synchronously and asynchronously hatching broods of Guira Cuckoos. Sample sizes reflect only nondepredated nests for which hatching dates of all chicks were known
Seven asynchronous nests suffered either partial or total brood mortality, of which details regarding dates of chicks hatching and disappearing are available for three cases (nests A81/94, B91/97, C16-1/97; codes reflect usage in previous publications). In nest A81/94 chicks were not individually marked, and individual dates of hatching and dying are not known. However, none of the eight chicks that hatched over three days survived for longer than the first week, whatever may have been their hatching order. In nests B91/97 and C16-1/97, nestlings were marked as soon as they hatched, and the exact order of hatching and dying is shown in Table 3. From these two nests, the order of hatching appears to be unrelated to the chance of dying in asynchronous broods.
Sequence of chick hatching and dying in two asynchronously hatching broods. The letter “C” designates the chick's identity, and Day 1 refers to the first day that either hatching (Hatch sequence) or dying (Death sequence) occurred in the brood
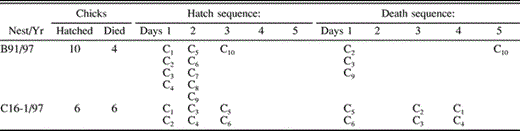
Sequence of chick hatching and dying in two asynchronously hatching broods. The letter “C” designates the chick's identity, and Day 1 refers to the first day that either hatching (Hatch sequence) or dying (Death sequence) occurred in the brood
Group Size and Reproductive Parameters
Guira cuckoo group sizes are associated with the provisioning capacity for rearing chicks (Macedo 1994): adults in large groups appear to feed chicks at the same rate as adults in smaller groups (food delivery rates were corrected for brood size to yield per-capita food consumption). Thus, one expectation would be that broods of different sizes produced by same-sized groups would be reduced to roughly the same size. For this analysis, 41 groups were used, and divided into the following categories: small, with less than 5 members (n = 6); average, with 5–8 members (n = 26); and large, with 9–10 members (n = 9). In two of these categories brood size showed a significant or close to significant association with the percent brood mortality (small groups: r = 0.27, n = 6, P = 0.61; average groups: r = 0.45, n = 26, P = 0.02; large groups: r = 0.64, n = 9, P = 0.06). Another expectation is that group size should be a good predictor for several reproductive variables. Regressions were therefore calculated for clutch size (F1 110 = 33.2, P < 0.001), percent clutch mortality (F1 107 = 0.06, P = 0.81), brood size (F1,75 = 2.1, P = 0.15), and percent brood mortality (F1,52 = 0.01, P = 0.92). Thus, group size has a significant effect upon clutch size, but not upon brood size, or percent mortality of either clutches or broods.
Total Elimination of Broods
Of the 58 nests that produced nestlings for which fates could be determined, 13 (22%) resulted in total nest failure. Predation was the likely cause of disappearance of nestlings in only two of these nests because: (1) no dead or wounded chicks were found in the nest vicinity; (2) all chicks disappeared simultaneously; and (3) the nest showed visible signs of disturbance. In the remaining 11 cases, several factors point toward elimination of nestlings by adults. For example, in eight instances, all nestlings disappeared sequentially within the first week of hatching, a pattern that is not a likely result of predation. In over half the nests resulting in total nest failure, there was indirect evidence of infanticide: dead nestlings, some with wounds, were found inside the nest or on the ground. In one of these nests, infanticide was observed directly.
Discussion
When infanticide is a major source of mortality, we expect it to have evolved as the result of enhanced fitness benefits for the infanticidal adults. It should also have considerable impact upon parenting behavior and the social patterns of the species in question (Hrdy et al. 1995). This Guira Cuckoo population suffered high rates of chick mortality: of the 58 nests with hatchlings and complete data sets, 42 (72%) fledged fewer chicks than hatched. Of these, 16 (38%) were characterized by direct observations of infanticide or by strong circumstantial evidence in the form of wounded or dead chicks inside the nest or on the ground.
In general, we found no support for the proposition that Guira Cuckoos may commit infanticide as a brood reduction mechanism in the Lackian sense (Lack 1947, 1954) to benefit reproductive individuals in the group. Hatching asynchrony is not a common pattern in this species, and when it does occur, there are no readily apparent benefits in terms of survival of the clutch or brood. For example, the comparison of reproductive parameters between synchronous and asynchronous broods (Table 2) showed no advantages for the latter in any of the characteristics evaluated.
One consequence of hatching asynchrony that has been observed in several species is that last-hatched chicks experience greater mortality (Ricklefs 1965, Fujioka 1985, Magrath 1990). In the small number of Guira Cuckoo nests hatching asynchronously, and for which dates of hatching and dying of all chicks were known, there is no evidence that last-hatched young are victimized by infanticide more frequently than older chicks. An associated expectation is that asynchronous hatching could be more common or extreme in parts of the season when food is less predictable. This has been shown to occur, for example, in Great Tits (Parus major,Gibb 1950) and in Common Terns (Sterna hirundo,Nisbet and Cohen 1975). Asynchronous nests in the Guira Cuckoo population studied did not occur predominantly toward the end of the reproductive season, which is when the rain tapers off and insects decrease in abundance.
If the recorded chick deaths were to contribute to the fitness of reproductive members in the group, then one expectation would be that larger broods should be reduced proportionately more than smaller broods, when comparisons are made for groups with the same number of adult members. Furthermore, group size should be a strong indicator of reproductive outcomes, since a group's provisioning capacity should be linked to the number of nestlings that could be reared successfully. These predictions were only partially supported: brood size reduction was related to the initial number of nestlings produced in average-sized groups, and nearly so in large groups. One interesting result was that, although group size significantly affects clutch size, it does not determine the number of hatchlings produced or the mortality patterns of eggs or chicks. Apparently, egg ejection disrupts the degree of association between group size and hatchlings produced.
The fact that over 20% of all nests with hatchlings experienced total brood failure is not compatible with brood reduction favoring reproductive group members in general. A more plausible scenario is that infanticide may provide a breeding opportunity for the infanticidal individual (sexually selected infanticide). For example, in the communal Acorn Woodpecker (Melanerpes formicivorus,Koenig 1990), sexually selected ovicide was induced through experimental manipulations of perceived paternity. However, our data do not provide unequivocal support for the sexually selected infanticide explanation. For instance, we did not find a greater incidence of egg ejection and infanticide early in the season, when renesting opportunities may be greater. Additionally, total brood loss is not the overall standard of mortality within broods, as one could expect in sexually selected infanticide. It may be that group composition obscures patterns somewhat. In groups where reproductive opportunity is balanced, we can speculate that infanticide should be unlikely in any part of the season, while in groups with a greater degree of reproductive skew, infanticide and ovicide should be more prevalent early in the season. Infanticidal individuals may also be unable to complete the elimination of the whole brood within the period of chick vulnerability (which lasts only about five or six days), resulting in partial brood elimination in many cases.
To support the hypothesis of sexually selected infanticide, killers should be unrelated to their victims; even stronger evidence would be to establish that, by committing infanticide, the killer promotes its own reproductive interests. Unfortunately, such evidence remains unavailable. If future research provides incontestable support for the sexually selected nature of infanticide in Guira Cuckoos, it would be crucial to assess the importance of this behavior in the shaping of mating patterns and parental investment, and in light of the recent developments in optimal skew and incomplete control models that predict reproductive sharing within social groups (Reeve et al. 1998, Johnstone and Cant 1999).
We thank José Luis Osorno, Guarino Colli and James Quinn for their insights on previous drafts of the manuscript. Field help was provided by Leandro Baumgarten, Juliana Bosi, Alice Guimarães, Daniela Sankievicz, Rafael Oliveira, Marcelo Simon, Andrei Polejack, Carmen Araújo, Barbara Fonseca, and Marcos Lima. RHM is a Research Fellow of the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq); MC and LM received financial support from CNPq. This work was funded by Fundo Nacional do Meio Ambiente, the Animal Behavior Society (Developing Nations Research Grant), and by the Fundação de Apoio a Pesquisa do DF (FAP-DF).
Literature Cited
