





Abstract
We documented effects of Brown-headed Cowbird (Molothrus ater) parasitism on Chipping Sparrows (Spizella passerina) in southwest Colorado. Overall, 17 of 77 nests (22%) were parasitized. Abandonment was significantly higher among parasitized nests (29%) than unparasitized nests (5%). More Chipping Sparrows hatched and fledged per unparasitized nest than per parasitized nest. Reduction of host hatching and fledging rates in parasitized nests was attributable to smaller clutch size and higher abandonment. The major source of nest failure was predation, not parasitism; overall, 53% failed from predation, and only 6% failed from parasitism. There were no differences in weight, ulna length, tarsometatarsus length, or length of outermost primary between sparrow nestlings in parasitized and unparasitized nests. Only 18% of cowbird eggs laid resulted in a fledged cowbird. We observed no differences in nest placement or nest concealment between unparasitized and parasitized nests.
Introduction
Brown-headed Cowbirds (Molothrus ater) are often implicated in the decline of many migratory songbirds and in the endangered status of Kirtland's Warbler (Dendroica kirtlandii,Walkinshaw 1972, Mayfield 1973, 1977), Least Bell's Vireo (Vireo bellii pusillus,Goldwasser et al. 1980, Franzreb 1990), Southwestern Willow Flycatcher (Empidonax traillii extimus,Sogge et al. 1997), Golden-cheeked Warbler (Dendroica chrysoparia), and Black-capped Vireo (Vireo atricapillus,Benson and Benson 1990). In these species, cowbird parasitism often results in a complete loss of host nestlings because these hosts do not possess effective anti-parasite defenses, other than abandonment, which is effective only when the host renests. Loss of host nestlings, however, is not a foregone conclusion for all hosts. Some hosts, such as Red-winged Blackbirds (Agelaius phoeniceus), are aggressive toward intruding parasites and can raise their own young along with cowbirds because these hosts are larger than cowbirds; further, they experience an overall low level of parasitism in their populations (Ortega and Cruz 1991, Ortega 1991, 1998, Prather et al. 2000). Other hosts, such as Yellow Warblers (Dendroica petechia), can raise their own young with cowbirds because they have an incubation period equally short or shorter than the cowbird's incubation period of 10–11 days, and their young develop as rapidly as cowbird nestlings (Ortega and Ortega 2000).
Other than rates of parasitism, little has been reported regarding how cowbird parasitism affects the nesting success of Chipping Sparrows (Spizella passerina). This is surprising as Chipping Sparrows breed throughout most of the United States and much of Canada and serve as one of the many hosts to Brown-headed Cowbirds (Friedmann and Kiff 1985, Ortega 1998). Rates of cowbird parasitism on Chipping Sparrows vary greatly, from 0% in Louisiana and California (Goertz 1977, Verner and Ritter 1983) to 92% in Ontario (Scott and Lemon 1996). With a pressing need to conserve migratory songbirds and with the growing interest in controlling cowbird populations, it is imperative to document more than rates of parasitism. We must also document why nests fail, how parasitism affects the quantity and quality of hosts fledged, what ecological factors increase the risk of cowbird parasitism and predation, and what anti-parasite defenses exist in the population. It is equally important to document the quality of hosts for the cowbird.
Although the overall abundance of Chipping Sparrows has been relatively stable since 1966 and has even increased throughout much of their range, De Sante and George (1994) suggested that habitat disturbance may have increased risks of parasitism on Chipping Sparrows. In the intermountain grasslands Breeding Bird Survey region (the region that encompasses southwestern Colorado, J. Sauer, pers. comm.), Chipping Sparrows declined at 0.2% per year from 1966–1998, but throughout Colorado, they declined 1.6% per year from 1966–1998 (P = 0.23), and in New Mexico, they declined 3% per year (P < 0.01, Sauer et al. 1999). In this study, we documented the effects of brood parasitism and predation on a population of Chipping Sparrows in southwestern Colorado.
Methods
From May through July, 1996–1998, we searched for Chipping Sparrow nests at the Colorado State University San Juan Basin Research Center, 8 km south of Hesperus, La Plata County, Colorado (37°14′N, 108°3′W). The Research Center, located along the La Plata River, consists of pastures grazed by cattle at various intensities (0–0.4 head ha−1). The riparian pastures are dominated by narrow-leaf cottonwoods (Populus angustifolia) and to a lesser degree by riverbirch (Betula fontinalis). The riparian zone varies from approximately 5–200 m on either side of the river; shrubs and other heavy understory are scarce. The upland pastures are dominated by Gambel oak (Quercus gambelii) and grasses.
We visited nests every one to three days and recorded the contents of each nest until all birds fledged or until the nest failed through predation or abandonment. We considered predation of nestlings to have occurred if all nestlings disappeared from the nest before they were eight days old or if one or more nestlings disappeared without signs of brood reduction, that is, if larger nestlings were missing. We estimated nest height and nest-plant height using 2-m poles. On each visit, we took the following measurements on all nestlings to the nearest 0.1 mm with a Mitutoyo®: tarsometatarsus, ulna, outermost primary (quill plus emerging feather). We also measured weight to the nearest 0.1 g with a 10-g, 30-g, or 50-g Pesola spring scale, using the smallest scale possible. In addition to the 1996–1998 data, we included one parasitized nest from 1995 in one of the same pastures to increase our sample size of Chipping Sparrow growth. This nest was used for growth information only and was not included in any other analysis.
In 1998, we quantified nest concealment as soon as possible after finding each active nest by taking photographs from 3 vantage points: 1 m directly below the nest, 1 m directly above the nest, and 1 m at the level of the nest toward the center of the nest plant. On photocopies (enlarged by 200%) of the photographs, we drew circles around nests and highlighted all vegetation covering the nests. We placed a transparent grid with 6-mm squares over the highlighted photocopy and counted the total number of squares the nest covered and the number of squares with vegetation covering the nest. We included all squares with ≥50% coverage and defined nest concealment as percent coverage (number of squares with vegetation over the nest divided by the total number of squares occupied by the nest).
Statistical Analyses
Most of our data did not fit a normal distribution; therefore, we used nonparametric statistical tests. For contingency tables, we used chi-square goodness of fit tests with Yates' correction, and to detect differences in rank values between sample populations, we employed two-tailed Mann-Whitney U-tests, corrected for ties when appropriate (Zar 1996). We used the Spearman rank correlation test to determine if numbers of depredated and parasitized nests were correlated with the number of available nests in one-week periods over the nesting season (Zar 1996). We considered a P-value of less than 0.05 to be significant. Standard deviations are provided with all mean values.
Results
We found a total of 77 nests: 9 in 1996, 28 in 1997, and 40 in 1998. We found the majority (90%) in the early stages of nesting (nest construction, egg laying, or early incubation). We found more Chipping Sparrow nests in Gambel oak pastures (n = 65) than in riparian pastures (n = 12), and the majority of nests (77%) were constructed in Gambel oak. Other nests were constructed in hawthorn (Crataegus spp.), narrow-leaf cottonwood, junipers (Juniperus spp.), riverbirch, and unidentified herbs.
Overall, 17 of 77 nests (22%) were parasitized by cowbirds; all parasitized nests contained only one cowbird egg. One nest contained only a cowbird egg; therefore, we did not know the outcome as it could have been abandoned or parasitized following depredation. In another nest, which we followed from the egg-laying stage, all but one of the nestlings disappeared; the remaining nestling died, apparently of starvation. It was unclear whether the other nestlings starved and were removed by the parents or whether they were preyed upon. These two nests were included as unsuccessful nests, but they were not included in further analyses of final outcomes (abandonment and predation).
Thirty of the 77 nests (39%) were successful in fledging at least one Chipping Sparrow. In no case did a cowbird fledge from a nest without a Chipping Sparrow also fledging. Forty nests (53%) failed due to predation; 37 of these nests were fully depredated, and three were abandoned after partial predation. Of the 37 fully depredated nests, 33 (89%) were in the egg-laying or incubation stages when predation occurred, and only four nests (11%) with chicks were depredated. Five additional nests were abandoned after a host egg was removed, presumably by a cowbird.
A higher proportion of unparasitized nests (26 of 60, 43%) than parasitized nests (4 of 17, 24%) were successful in fledging at least one host, but the difference was not statistically significant (χ21 = 1.4, P > 0.20). Failure due to predation was similar between unparasitized nests (7 of 17, 41%) and parasitized nests (33 of 59, 56%, χ21 = 0.6, P > 0.40). However, a significantly higher proportion of parasitized nests (5 of 17, 29%) than unparasitized nests (3 of 59, 5%) were abandoned (χ21 = 5.9, P < 0.05). More Chipping Sparrows hatched and fledged from unparasitized nests than from parasitized nests considering all nests and successful nests only (Table 1).
Mean number of Chipping Sparrows hatched and fledged per nest and per egg among unparasitized nests and parasitized nests, San Juan Basin Research Center, La Plata County, Colorado, 1996–1998.[sp[fy181,1][cf11]a[cf1][rp [cf2]U[cf1] and associated [cf2]P[cf1]-values reflect Mann-Whitney [cf2]U[cf1]-tests
![Mean number of Chipping Sparrows hatched and fledged per nest and per egg among unparasitized nests and parasitized nests, San Juan Basin Research Center, La Plata County, Colorado, 1996–1998.[sp[fy181,1][cf11]a[cf1][rp [cf2]U[cf1] and associated [cf2]P[cf1]-values reflect Mann-Whitney [cf2]U[cf1]-tests](https://oup.silverchair-cdn.com/oup/backfile/Content_public/Journal/condor/103/1/10.1093_condor_103.1.127/1/m_i0010-5422-103-1-127-t01.gif?Expires=1748892867&Signature=baMiUHJaE-LeNzHVV4LO4aCKGyheVUSaa3lgxP49qrmHfaEbsxtTQhAnc-KMkkp3vGaT9L6feF3hDu0dzUSUv2VLfJ-VQhTXaD3iGktk-G93StmENPm0lmSqiPifbnYRuoHr-TasLICJUK0BuFJjGtuBBOUccY7zPIZarAo7tSnQ6tDmAwzsVQYjVFjk5~My6WdFTtqJyhgt4JTHIdduBjrdsq4AzAD1U5chg3MdGwNeNkSrdtJ3Lq92AcPTE17U7kMC2CXKZAkuQPjnH3iBvmE203t0CbhZv8Py5GW65pjaentVD7BrdcTe3FehPVVmziHarYws91yzRUZ1Wt5LTQ__&Key-Pair-Id=APKAIE5G5CRDK6RD3PGA)
Mean number of Chipping Sparrows hatched and fledged per nest and per egg among unparasitized nests and parasitized nests, San Juan Basin Research Center, La Plata County, Colorado, 1996–1998.[sp[fy181,1][cf11]a[cf1][rp [cf2]U[cf1] and associated [cf2]P[cf1]-values reflect Mann-Whitney [cf2]U[cf1]-tests
Reduction of host fledglings in parasitized nests was not attributable to brood reduction but rather to smaller clutch size and higher frequency of abandonment among parasitized nests. The mean clutch size of all Chipping Sparrow nests with completed clutches was 3.6 ± 0.8 eggs (range: 2–5 eggs, n = 48). The clutch size of parasitized nests (2.9 ± 0.8 eggs; range: 2–4 eggs, n = 8) was significantly smaller than clutch size of unparasitized nests (3.7 ± 0.7 eggs; range: 2–5 eggs, n = 40, U = 72, P < 0.01). Among successful nests, there were no differences between parasitized and unparasitized nests regarding the mean number of hosts hatched and fledged per egg (Table 1). However, excluding depredated nests, significantly more hosts hatched and fledged per egg in unparasitized nests than parasitized nests (Table 1). Failure of undepredated nests was due to abandonment of parasitized nests. However, the major source of nest failure was clearly predation, not cowbird parasitism; overall, 53% failed due to predation, and only 6% failed due to cowbird parasitism. The mean incubation period (from the last egg laid) was 10.4 ± 1.0 days (n = 36), and the mean age of fledging was 8.5 ± 0.8 days (n = 17). Our sample size of Chipping Sparrow nestlings raised with a cowbird was too small (n = 6 nestlings) to conduct statistical analyses. However, there were no obvious differences between parasitized and unparasitized nests in the measurements of weight, ulna length, tarsometatarsus length, or length of outermost primary (Fig. 1).
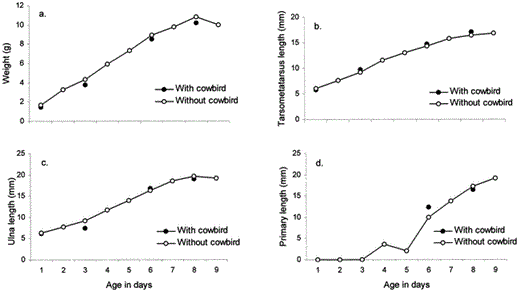
Chipping Sparrow growth (a. weight, b. tarsometatarsus length, c. ulna length, d. length of outermost primary) in nests with and without a Brown-headed Cowbird nestling. Growth is based upon measurements of 75 individuals in 24 unparasitized nests and 6 individuals in 4 parasitized nests
The success of cowbirds was low in the nests of Chipping Sparrows. A mean of 0.23 ± 0.44 (n = 17) cowbirds hatched per parasitized nest and per egg (as all parasitized nests contained only one cowbird egg), and a mean of only 0.18 ± 0.39 (n = 17) cowbirds fledged per parasitized nest and per egg.
Nests in riparian pastures did not differ from nests in oak pastures regarding percent success (riparian: 50%, n = 12, oak: 37%, n = 65, χ21 = 0.3, P > 0.50) or percent parasitism (riparian: 17%, n = 12, oak: 23%, n = 65, χ21 = 0.01, P > 0.90). We observed no differences between unparasitized and parasitized nests regarding mean nest height or percent nest concealment (Mann-Whitney U-tests, all P > 0.05).
The date of clutch initiation did not differ among years (Kruskal-Wallis test, corrected for ties, H = 0.26, df = 2, P = 0.88); therefore, we pooled these data among years. The number of cowbird eggs laid was significantly correlated with the number of clutches initiated in one-week periods (rs = 0.70, P = 0.04, Fig. 2a). Predation was highest when more nests were available; however, the correlation fell shy of the accepted P-value (rs = 0.64, P = 0.06, Fig. 2b).
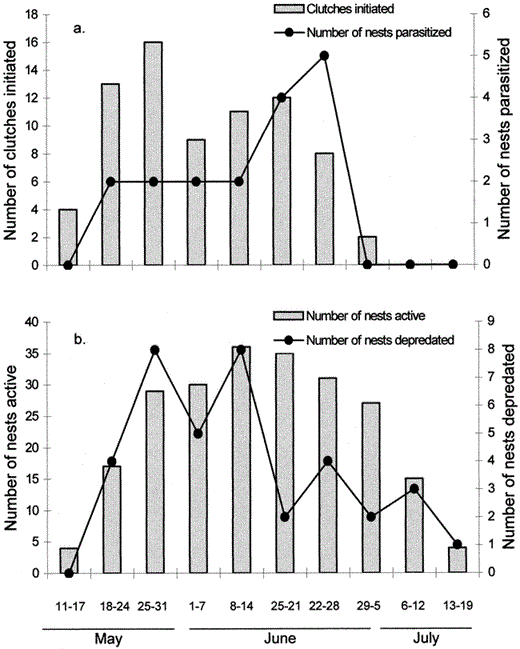
(a) Number of Chipping Sparrow clutches initiated and number of nests that were parasitized by Brown-headed Cowbirds during one-week periods, and (b) number of Chipping Sparrow nests active and number of nests that were depredated during one-week periods, 1996–1998
Discussion
Unparasitized nests appeared to be considerably more successful than parasitized nests in terms of both the percent of nests fledging at least one host and the mean number of hosts fledged. Parasitized nests failed more frequently due to predation (although not significantly so) than unparasitized nests, but this does not entirely explain the difference in success. The other source of failure among parasitized nests was nest abandonment, which occurred significantly more often compared to unparasitized nests. Although abandonment of parasitized nests in this study was nearly 30%, this rate is not as high as Graham (1988) found in Ontario (52%, n = 100). Abandonment rates of parasitized nests are also high among other Spizella species. For example, 59% (n = 22) of parasitized Clay-colored Sparrow (Spizella pallida) nests were abandoned in Manitoba (Hill and Sealy 1994), and 62% (n = 16) of parasitized Field Sparrow (Spizella pusilla) nests were abandoned in Illinois (Best 1978). Similarly, B. Strausberger and D. Burhans (pers. comm.) found that Field Sparrows in Illinois abandoned 46% (n = 76) of parasitized nests; however, they found that Field Sparrows did not abandon artificially parasitized nests unless the researchers also presented a mounted cowbird. Other studies also suggest that nest abandonment may not be due to recognition of cowbird eggs (Rothstein 1975, 1982, Hosoi and Rothstein 2000). In our study, only 5 of 77 nests (6%) appeared to be abandoned due to cowbird activity; therefore, relatively few of the total nests were lost directly to parasitism. All parasitized nests that were not abandoned or depredated fledged at least one Chipping Sparrow along with a cowbird.
Some of the pairs that abandoned their nests may have renested. Indeed, Pease and Grzybowski (1995) suggested that analysis of females over the breeding season is preferable to analysis of nest parameters because parasitized hosts may abandon their nests and be successful in subsequent brood attempts, canceling out earlier nest failures. In other words, following a particular female over the entire breeding season may show a higher level of success, presuming all nests are found and followed. Therefore, an analysis of nest parameters may actually overestimate the effects of cowbird parasitism on a population.
Another effect of cowbird parasitism was a lower mean number of hosts fledged in parasitized nests compared with unparasitized nests. Among successful nests, we observed no difference in the mean number of hosts fledged per egg laid between parasitized and unparasitized nests; therefore, the lower mean number of hosts fledged per parasitized nest was due to the lower clutch size of parasitized nests. This is similar to Middleton's (1998) observation in Ontario that cowbird parasitism reduced mean Chipping Sparrow clutch size by approximately one. In addition to the lower observed clutch size among parasitized nests, we also observed a higher rate of partial predation among parasitized nests (47%) than among unparasitized nests (14%). We assumed that if parasitized nests were abandoned after one or more host eggs disappeared, then cowbirds had removed the eggs. Indeed, removing the five abandoned nests leaves a similar percentage of partial predation between parasitized nests (18%) and unparasitized nests (14%). The loss of hosts produced in parasitized nests, therefore, occurred during the egg-laying or incubation stages, not during the nestling period. Although our sample size of successful parasitized nests was low, in these nests Chipping Sparrows appeared to develop as well as they did in unparasitized nests (Fig. 1). This is attributable to both their short incubation period and rapid nestling development. Similar to Middleton's (1998) observations, we found that Chipping Sparrows can fledge in as little as 8 days after hatching. Both the short incubation period and rapid development are similar to what we found in Yellow Warblers at the same study site, where hosts also fledged along with cowbirds (Ortega and Ortega 2000).
Compared with other hosts in southwestern Colorado, Chipping Sparrows do not appear to be a high-quality host. Only 18% of the cowbird eggs laid in Chipping Sparrow nests resulted in a fledged cowbird. This poor success is similar to the fledging success of cowbirds in Ontario (20%, Middleton 1998) but much lower than some other hosts at our study site. For example, of the cowbird eggs laid at our study site from 1992–1998, Yellow Warblers fledged 42% (n = 31), Warbling Vireos (Vireo gilvus) fledged 44% (n = 18), and Plumbeous Vireos (Vireo plumbeus) fledged 33% (n = 6). Even though Chipping Sparrows appear to be poor hosts, they represent 20% of the cases of parasitism among 10 common hosts at our study site. This suggests that cowbirds may not select hosts according to quality, as measured by cowbirds fledged per egg.
We found no differences between parasitized and unparasitized nests regarding nest height, plant height, or nest plant. We also observed no differences in parasitism between oak and riparian pastures. Although many Chipping Sparrow nests are very well concealed by vegetation, percent concealment also did not differ between parasitized and unparasitized nests or between depredated and undepredated nests, lending support to the hypothesis that cowbirds cue in on activity of their hosts to locate nests (Norman and Robertson 1975, Thompson and Gottfried 1976, 1981, Ortega 1998). We found the majority of Chipping Sparrow nests in Gambel oak pastures. Gambel oak is a dominant component of vegetation in much of the Four Corners region (Harper et al. 1985) where Chipping Sparrows may be declining (Sauer et al. 1999). Gambel oak dominated lands in the arid southwest are also heavily grazed by cattle. Interestingly, in a preliminary study on the effects of grazing regimes, we found that Chipping Sparrow success was significantly improved, both in terms of predation and cowbird parasitism, in pastures that were ungrazed during the breeding season. Although cowbird parasitism significantly reduced the number of Chipping Sparrows fledged, predation was unequivocally the major source of nest failure among Chipping Sparrows in this study.
Acknowledgments
We thank our numerous field assistants: Susan Allerton, John Arnett, Stacia Backensto, Florence Kittleman, Becky Merris, Jonathan Nardelli, Cristin Rapp, and Jessica Young. Financial assistance was provided by grants from the National Geographic Society, the National Fish and Wildlife Foundation, Howard Hughes Medical Institute, and the Colorado Alliance for Minority Participation. We also thank David Schafer for access to the San Juan Basin Research Center and for his logistical support, and two anonymous reviewers for their comments.
Literature Cited

{kind=link}
{kind=link}