





Abstract
Coronary artery disease (CAD) poses a substantial threat to global health, leading to significant morbidity and mortality worldwide. It has a significant genetic component that has been studied through genome-wide association studies (GWAS) over the past 17 years. These studies have made progress with larger sample sizes, diverse ancestral backgrounds, and the discovery of multiple genomic regions related to CAD risk. In this review, we provide a comprehensive overview of CAD GWAS, including information about the genetic makeup of the disease and the importance of ethnic diversity in these studies. We also discuss challenges of identifying causal genes and variants within GWAS loci with a focus on non-coding regions. Additionally, we highlight tissues and cell types relevant to CAD, and discuss clinical implications of GWAS findings including polygenic risk scores, sex-specific differences in CAD genetics, ethnical aspects of personalized interventions, and GWAS guided drug development.
1. From inception to present: tracing 17 years of GWAS progress
Coronary artery disease (CAD), caused by atherosclerosis, is ubiquitous in our society, and its end-stage complications [e.g. myocardial infarction (MI) and stroke] are the primary cause of morbidity and mortality worldwide.1 The heritability of CAD is estimated to be 40–70%, which suggests that there is a considerable genetic contribution to its pathology.2
The first genome-wide association study (GWAS) for CAD was performed in 2007, identifying the 9p21 risk locus to be strongly associated with CAD, which has been replicated in many other GWAS for CAD.3–5 Within the last 17 years, the sample size has increased to >1 million, the price of the genotyping array has decreased, sequencing power and statistical methodologies (such as imputation and haplotype tagging) have improved, and national and international consortia have been established (Figure 1). These developments have contributed dramatically to expanding the genome-wide significant CAD loci to 393, complemented with ever more accurate prioritization of candidate causal genes and single nucleotide polymorphisms (SNPs). Table 1 summarizes the CAD genome-wide significant loci identified by recent GWAS for CAD.6–9 In addition to previous CAD GWAS reviews,10–17 we explore challenges in deciphering the causality of variants and genes. Moreover, we propose tissues or cell types most relevant to atherosclerosis and provide a comprehensive picture of pathophysiological pathways beyond lipid metabolism. Finally, we discuss the clinical implications of GWAS in terms of risk stratification, sex-specific and ethnic-specific differences in CAD genetics, and drug development.
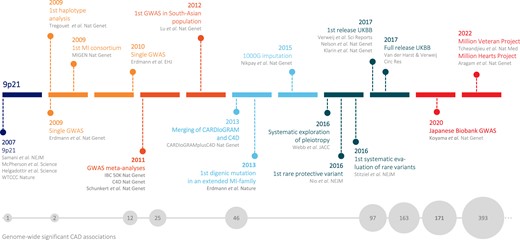
Important milestones in CAD genetics. The original version of this figure was published in a previous review from 2018 Erdmann et al.10 and subsequently updated by the authors.
Summary of genome-wide significant CAD risk loci
| Locus ID | Genes (protein) | Genes (RNA) | Index rsIDs | Coordinates (hg38) | Traits | Ethnicity |
|---|---|---|---|---|---|---|
| 1 | SKI4, MORN13 | rs2843152 | Chr1:2314131 | L | EUR | |
| 2 | PRDM167, ACTRT22 | rs7413494, rs2493298 | Chr1:3069321-3409348 | BP | EUR | |
| 3 | DHDDS7, RPS6KA13 | DHDDS-AS12 | rs12046497 | Chr1:26521149 | BH, L | Multi |
| 4 | ARID1A7, KDF12 | LOC1019287282 | rs79598313 | Chr1:26958422 | ALC, BH, BP, I, L | EUR |
| 5 | FHL37, INPP5B6 | rs61776719 | Chr1:37995647 | BH, BMI, BP | EUR | |
| 6 | ZMYND125, FOXO64 | rs12047439, rs2152314 | Chr1:41343968-42480791 | L, BP | Multi | |
| 7 | PCSK916, USP244 | rs34232196, rs11591147, rs151193009, rs472495 | Chr1:55023869-55055640 | ALC, BC, L, PSY | EUR, JPN | |
| 8 | PLPP310, FGGY8 | LINC013584, AC093425.12 | rs11206803, rs56170783, rs71646019, rs12733512 | Chr1:56411837-59181306 | BH, BMI, BP, SM | EUR, HISP, JPN, Multi |
| 9 | PSRC18, CELSR26 | rs12740374, rs655246 | Chr1:109274968-109289661 | ALC, BC, BH, BP, I, L, PSY | BLK, EUR, HISP | |
| 10 | PHTF13, MAGI32 | rs1230666 | Chr1:113630788 | EUR | ||
| 11 | NGF8, TSPAN22 | rs11806316, rs61797068 | Chr1:115210861-115359893 | EUR | ||
| 12 | ECM16, MTMR115 | ADAMTSL4-AS12, AL356356.12 | rs67807996, rs11585169 | Chr1:150023307-150599561 | BH, BMI, BP, I, SM | EUR |
| 13 | TDRKH9, RIIAD13 | TDRKH-AS13 | rs11810571 | Chr1:151789832 | PSY | EUR |
| 14 | IL6R5, TPM32 | rs6686750 | Chr1:154447367 | I | EUR | |
| 15 | NME79, ATP1B15 | rs61806987 | Chr1:169345595 | BH | EUR | |
| 16 | KIAA00405, TNN1 | rs4650716 | Chr1:175159575 | EUR | ||
| 17 | DENND1B4, CRB12 | rs12022672 | Chr1:197618876 | SM | Multi | |
| 18 | DDX597, CAMSAP24 | DDX59-AS12 | rs6700559 | Chr1:200676945 | EUR | |
| 19 | LMOD17, IPO94 | IPO9-AS12 | rs2820315 | Chr1:201903136 | BH, BMI, BP, L, PSY | EUR |
| 20 | DSTYK3, NUAK23 | AC119673.22 | rs12048743 | Chr1:205145745 | L | Multi |
| 21 | HHAT7, SYT142 | RNU5A-8P2 | rs60154123 | Chr1:210295654 | EUR | |
| 22 | MIA36, AIDA4 | MIA3-AS12 | rs17163363 | Chr1:222655362 | BP | EUR, HISP, JPN |
| 23 | AGT5, COG21 | rs699 | Chr1:230710048 | BP | EUR | |
| 24 | COLEC112 | RNASEH1-DT3 | rs56236159 | Chr2:3588888 | BC, BP, I, L | Multi |
| 25 | IAH13, ADAM173 | rs2715879 | Chr2:9375422 | EUR | ||
| 26 | OSR13, TTC322 | rs16986953 | Chr2:19742712 | EUR | ||
| 27 | APOB5, TDRD152 | rs13306206, rs515135 | Chr2:21019859-21063185 | BC, L, PSY | EUR, JPN | |
| 28 | ABCG810, ABCG56 | rs4245791, rs76866386 | Chr2:43847292-43848344 | BC, I, L, PSY | EUR | |
| 29 | PRKCE7, SRBD11 | rs582384 | Chr2:45669298 | BP | EUR | |
| 30 | BCL11A4, REL1 | MIR4432HG2 | rs243071 | Chr2:60391893 | EUR | |
| 31 | GFPT15, NFU13 | rs4346430 | Chr2:69441670 | Multi | ||
| 32 | GGCX7, VAMP86 | PARTICL2 | rs10176176 | Chr2:85534925 | BMI, L | EUR, JPN |
| 33 | IL1F105, PSD43 | rs6759676 | Chr2:113078771 | BP, I | EUR | |
| 34 | SAP1305, WDR331 | rs114192718 | Chr2:128028089 | EUR | ||
| 35 | ARHGAP155, KYNU1 | rs4662330 | Chr2:143428906 | BP | EUR | |
| 36 | ZEB212, GTDC13 | LINC014123, AC009951.42 | rs6740731, rs17408036, rs10928241 | Chr2:144513025-145073861 | BP, SM | EUR, HISP |
| 37 | ACVR2A4, MBD52 | rs35611688 | Chr2:147620292 | EUR | ||
| 38 | FIGN5, GRB141 | rs10930115 | Chr2:164073872 | BP, PSY, SM | EUR, JPN | |
| 39 | TFPI4, CALCRL3 | rs840616 | Chr2:187331742 | BMI, BP, L | EUR | |
| 40 | NBEAL17, CARF7 | rs148812085 | Chr2:203012510 | BMI, BP, L, PSY, SM | EUR, HISP | |
| 41 | FN17, ATIC2 | rs1250247 | Chr2:215434906 | BH, BMI, BP, L | EUR | |
| 42 | TNS15, TMBIM12 | rs2161967 | Chr2:217815806 | BP, L | EUR, HISP | |
| 43 | IRS15, NYAP22 | rs952227 | Chr2:226197364 | ALC, BMI, BP, D, L, SM | EUR | |
| 44 | PID15, DNER3 | rs4140748 | Chr2:229140789 | L | EUR, JPN | |
| 45 | GIGYF27, SNORC4 | rs283485 | Chr2:232780981 | BH, BMI | EUR | |
| 46 | COL6A34, RBM441 | rs11677932 | Chr2:237315312 | BP | EUR | |
| 47 | FGD55, NR2C21 | rs34991912 | Chr3:14884844 | BH, BP | EUR, JPN | |
| 48 | ZNF5897, CDC25A6 | AC134772.12 | rs7633770, rs7617773 | Chr3:46647072-48152025 | BH, BP | EUR |
| 49 | IP6K23, IHO13 | AC121247.12 | rs34759087 | Chr3:49124851 | BH, BMI, PSY | EUR |
| 50 | ITIH43, NISCH2 | rs77347777 | Chr3:52814191 | BMI, BP | EUR | |
| 51 | ITGB55, UMPS4 | rs17843797 | Chr3:124734175 | BMI | EUR | |
| 52 | DNAJC135, NPHP34 | rs10512861 | Chr3:132539117 | I, L | EUR | |
| 53 | PCCB7, PPP2R3A5 | NCK1-DT3 | rs34330586 | Chr3:136081567 | BMI, I, L | EUR |
| 54 | MRAS5, PIK3CB3 | rs185244 | Chr3:138374047 | BMI, BP, L | EUR | |
| 55 | ATP1B34, PLS12 | rs7622417 | Chr3:141907157 | BMI, L | Multi | |
| 56 | ARHGEF268, DHX367 | ARHGEF26-AS13 | rs357494 | Chr3:154219964 | BMI, BP, L | EUR, HISP |
| 57 | CCNL14, TIPARP4 | rs4266144 | Chr3:157134803 | BH, BMI | EUR | |
| 58 | MECOM2, SEC621 | rs16853198 | Chr3:169122391 | BP | Multi | |
| 59 | FNDC3B2, ECT21 | rs34229028 | Chr3:172399666 | EUR | ||
| 60 | rs73070813 | Chr3:187168807 | EUR | |||
| 61 | HGFAC2 | rs16844401 | Chr4:3447925 | BC, BH, L | EUR | |
| 62 | GNPDA21 | rs10938398 | Chr4:45184122 | BMI, I, L | Multi | |
| 63 | FIP1L12, LNX11 | rs17083333 | Chr4:53705899 | EUR | ||
| 64 | REST9, SPINK22 | rs781663 | Chr4:56915588 | BC, BH, I, L | EUR, JPN | |
| 65 | SHROOM35, CCDC1582 | rs12500824 | Chr4:76495474 | L, BP | EUR | |
| 66 | ANTXR22, PRDM81 | rs10857147 | Chr4:80259918 | L, BP | EUR, HISP | |
| 67 | RASGEF1B4, PRKG21 | rs36002015 | Chr4:81704567 | EUR | ||
| 68 | UNC5C2, BMPR1B2 | rs3775058 | Chr4:95196220 | EUR | ||
| 69 | MAD2L15, PDE5A4 | rs7678555 | Chr4:119988346 | BP | EUR | |
| 70 | PCDH182 | rs7439567 | Chr4:137543688 | BP | Multi | |
| 71 | ZNF8275, MMAA1 | rs13124853 | Chr4:145863622 | BP, L | EUR | |
| 72 | EDNRA14, ARHGAP103 | AC093908.12 | rs13120678, rs1878406, rs6841581 | Chr4:147352245-147480038 | BC, BP | EUR, JPN |
| 73 | GUCY1A110, MAP92 | RP11-588K22.22 | rs7440763, rs3796587 | Chr4:155512368-155716921 | ALC, BP, L | EUR, HISP, JPN |
| 74 | PALLD4, DDX60L1 | rs869396 | Chr4:168766849 | BP | EUR | |
| 75 | IRX13 | rs10041378 | Chr5:4094052 | JPN | ||
| 76 | SEMA5A7, TAS2R11 | SNHG185 | rs17263917 | Chr5:9552226 | BC | EUR |
| 77 | BASP12 | rs1032763 | Chr5:17118821 | Multi | ||
| 78 | ITGA15, PELO1 | rs4074793 | Chr5:52897294 | L, I | EUR, Multi | |
| 79 | ARL152, HSPB31 | AC027329.12 | rs7719168 | Chr5:53996560 | BP, L | EUR |
| 80 | MAP3K14, IL6ST2 | rs62362364, rs3936510 | Chr5:56145744-56565039 | BMI, BP, L, SM | EUR | |
| 81 | ANKDD1B7, HMGCR6 | rs12916 | Chr5:75360714 | BC, BMI, L, PSY | EUR | |
| 82 | LNPEP7, ERAP27 | AC009126.12 | rs27660 | Chr5:97012071 | BP, I, L, PSY | Multi |
| 83 | FER11, FBXL174 | LINC010233 | rs288173, rs112949822 | Chr5:108016593-108749489 | BH, BMI, BP, I | EUR, Multi |
| 84 | NREP1, WDR361 | rs256277 | Chr5:112071113 | Multi | ||
| 85 | DMXL12, TNFAIP81 | rs13169691 | Chr5:119112584 | EUR | ||
| 86 | SRFBP15, LOX3 | AC008568.12 | rs4345341 | Chr5:121943056 | BP | EUR |
| 87 | PRDM63, SNX241 | rs73796819 | Chr5:123084735 | Multi | ||
| 88 | FBN28, SLC27A62 | rs6883598 | Chr5:128590497 | BP | EUR | |
| 89 | IRF14, RAD504 | rs10477741, rs2706399 | Chr5:132459618-132532010 | L | EUR | |
| 90 | FGF16, SPRY42 | rs249760, rs3776307 | Chr5:142536127-143114600 | L | EUR | |
| 91 | SGCD4, MED72 | rs39929 | Chr5:156696640 | L, BP | Multi | |
| 92 | ADAM195, FNDC91 | rs11465228 | Chr5:157575687 | Multi | ||
| 93 | FOXC15, GMDS3 | rs72836800 | Chr6:1617092 | BH | EUR | |
| 94 | BLOC1S51, TXNDC51 | rs6597292 | Chr6:7975026 | Multi | ||
| 95 | PHACTR16, GFOD12 | rs9349379 | Chr6:12903725 | ALC, BC, BP, C | EUR, HISP, JPN | |
| 96 | HDGFL14, PRL1 | rs6909752 | Chr6:22612400 | AF | EUR | |
| 97 | HLA-A4, HLA-G3 | HLA-F-AS15, HLA-H3 | rs29266 | Chr6:29607502 | L | EUR |
| 98 | MICA4, PPP1R104 | rs139141104, rs9266631 | Chr6:31021244-31379121 | L | EUR, JPN | |
| 99 | CLIC13, DDAH23 | SNHG322, LOC1005075472 | rs3130683 | Chr6:31920590 | BH, BMI, BP, L, PSY | EUR |
| 100 | LEMD23, UQCC23 | rs62397561 | Chr6:33816328 | Multi | ||
| 101 | BLTP3A9, SNRPC8 | ILRUN-AS12 | rs9469899, rs17609940 | Chr6:34825347-35067023 | BH, BMI, L, PSY | EUR |
| 102 | CDKN1A5, ETV72 | DINOL2 | rs1321309 | Chr6:36670859 | BP, I, L | EUR |
| 103 | KCNK510, SAYSD11 | rs733701, rs10947789 | Chr6:39204086-39207146 | BP | EUR | |
| 104 | PEX63, PTK71 | rs1034246 | Chr6:43100632 | EUR | ||
| 105 | VEGFA4, MRPS18A1 | rs6905288, rs149372871 | Chr6:43791136-44080314 | ALC, BMI, BP, L, PSY | EUR, HISP, JPN | |
| 106 | RAB2310, PRIM26 | LOC1005061884, RP11-203B9.42 | rs11752218, rs9367716 | Chr6:57280764-57295774 | EUR | |
| 107 | CD1096 | CD109-AS12 | rs79717953, rs56171536 | Chr6:73695048-73706145 | JPN, Multi | |
| 108 | TENT5A4, IBTK1 | LINC0254`22 | rs9361867 | Chr6:81886242 | EUR | |
| 109 | UFL16, FHL55 | rs2983896 | Chr6:96581995 | BP | EUR | |
| 110 | CENPW8, TRMT112 | rs35510806 | Chr6:126357186 | BH, I, L, PSY | EUR | |
| 111 | TCF2113, SLC2A127 | TARID3 | rs6919211, rs2327426, rs2492304 | Chr6:133678730-134057013 | BH, BP | EUR, HISP, JPN |
| 112 | PLEKHG14, MTHFD1L2 | RAET1K2 | rs17080093 | Chr6:150676304 | BP | EUR |
| 113 | PLG29, LPA19 | LPAL22 | rs6932293, rs186696265, rs56195640, rs192425087, rs28867400, rs932631509, rs10455872, rs73596816, rs79018195, rs1998043, rs4252120, rs184278183 | Chr6:160114846-160756724 | BC, BMI, BP, C, L, PSY | BLK, EUR, HISP, JPN |
| 114 | MAD1L16, ELFN12 | rs62435159 | Chr7:1871645 | BMI | EUR | |
| 115 | DAGLB8, RAC17 | rs10951983 | Chr7:6406396 | I, L | EUR | |
| 116 | TMEM106B7, THSD7A2 | rs11509880 | Chr7:12222285 | L, PSY | EUR | |
| 117 | ETV14 | rs2073533 | Chr7:13990114 | EUR | ||
| 118 | TWIST15, HDAC92 | rs2107595 | Chr7:19009765 | BP | EUR, JPN | |
| 119 | MACC12, ITGB82 | rs10486389 | Chr7:20260793 | EUR | ||
| 120 | TBX208, DPY19L12 | DPY19L2P12 | rs2215614 | Chr7:35237481 | BH | EUR |
| 121 | CCM25, TBRG41 | rs2107732 | Chr7:45038379 | BP | EUR | |
| 122 | SEMA3C5 | rs917191 | Chr7:80941555 | BMI | Multi | |
| 123 | ZKSCAN16, AP4M15 | STAG3L5P2 | rs35146811 | Chr7:100123371 | BH, BP, L | EUR |
| 124 | COG59, BCAP298 | WBP1LP23, RPL7P322 | rs12112877, rs10953541 | Chr7:107300879-107604100 | L, PSY, SM | EUR |
| 125 | CFTR3, ASZ12 | rs13222797 | Chr7:117459992 | BMI, L | EUR | |
| 126 | ZC3HC15, KLHDC102 | rs11556924 | Chr7:130023656 | BC, BP, I, L, PSY, SM | EUR | |
| 127 | PARP122, HIPK21 | rs5887949 | Chr7:140060741 | BH | EUR | |
| 128 | NOS35, KCNH21 | rs3918226 | Chr7:150993088 | ALC, BC, BP, L, SM | EUR | |
| 129 | MSR13, CNOT71 | rs1480146 | Chr8:16272064 | Multi | ||
| 130 | NAT23, PCM13 | rs4646249 | Chr8:18402921 | L | EUR | |
| 131 | LPL14, CSGALNACT13 | rs894211, rs66778572, rs268 | Chr8:19956018-20058609 | ALC, BC, BH, BMI, BP, I, L, PSY, SM | BLK, EUR, HISP | |
| 132 | BMP15, PHYHIP2 | rs56408342 | Chr8:22190977 | EUR | ||
| 133 | DOCK56, KCTD92 | rs1510758 | Chr8:25204291 | EUR | ||
| 134 | PPP2R2A2 | rs6557894 | Chr8:26229397 | BH | EUR | |
| 135 | RP15 | rs9298506 | Chr8:54524964 | ALC, BH, I, L | Multi | |
| 136 | C8orf342 | rs2380472 | Chr8:68519476 | JPN | ||
| 137 | GEM6, CDH172 | rs34917849 | Chr8:94266079 | BP | EUR | |
| 138 | NDUFAF65, TP53INP11 | rs3019173 | Chr8:95008746 | Multi | ||
| 139 | rs10093110 | Chr8:105553186 | EUR | |||
| 140 | TRIB14, NSMCE21 | rs2001846 | Chr8:125466208 | ALC, BMI, BP, D, I, L | EUR, HISP | |
| 141 | NFIB2, MPDZ1 | LURAP1L-AS12 | rs10961206 | Chr9:13724052 | EUR | |
| 142 | ACER27, RPS61 | AL391834.12 | rs10811183 | Chr9:19436057 | L | EUR |
| 143 | CDKN2A13, CDKN2B11 | ERVFRD-36, MIR31HG4 | rs11523031, rs2891168, rs6475608, rs76959412 | Chr9:21843843-22101703 | BC, BP, C, L | BLK, EUR, HISP, JPN |
| 144 | WNK23, CARD192 | rs4617217 | Chr9:93226686 | Multi | ||
| 145 | ABCA12, NIPSNAP3B2 | rs35093463 | Chr9:104823957 | L | JPN | |
| 146 | KLF44, RAD23B1 | rs1967604 | Chr9:107768043 | BC, L | EUR | |
| 147 | SVEP14 | rs111245230 | Chr9:110407495 | BC, BP | EUR | |
| 148 | ASTN22 | rs7873551 | Chr9:116482848 | BH, PSY | Multi | |
| 149 | PHF195, C54 | rs41312891 | Chr9:120964472 | BH | EUR, Multi | |
| 150 | DAB2IP5, STOM1 | CUTALP2 | rs885150 | Chr9:121657894 | BC, BH, BMI, L | EUR |
| 151 | ABO6, CEL3 | rs651007 | Chr9:133278431 | BC, BP, D, I, L, PSY | BLK, EUR, HISP | |
| 152 | CAMK1D4, CDC1232 | rs17566555 | Chr10:12233948 | PSY | EUR | |
| 153 | ARHGAP214, GPR1581 | rs7077962 | Chr10:24765745 | Multi | ||
| 154 | JCAD7, SVIL1 | rs9337951 | Chr10:30028144 | BP, SM | EUR, HISP | |
| 155 | ZEB16, ARHGAP121 | ZEB1-AS15, PPIAP312 | rs161231 | Chr10:31466991 | L | Multi |
| 156 | CXCL1212, RASGEF1A2 | FRG1JP2 | rs1870634, rs494207, rs527052 | Chr10:43985363-44245808 | C | BLK, EUR, HISP |
| 157 | BICC17, TFAM1 | rs10740732 | Chr10:58593128 | BP | Multi | |
| 158 | TSPAN147, SH2D4B2 | LOC1019295742 | rs17680741 | Chr10:80491758 | EUR | |
| 159 | LIPA6, IFIT11 | rs1051338 | Chr10:89247603 | I, L | EUR, JPN | |
| 160 | PLCE112, SLC35G12 | rs1223583, rs55753709 | Chr10:93996743-94269413 | BMI, BP, L, PSY | EUR | |
| 161 | LOXL49, R3HCC1L3 | rs884811 | Chr10:98164006 | BH, BP | EUR | |
| 162 | CNNM28, BORCS75 | rs77787671 | Chr10:103016448 | BMI, BP, PSY | EUR, HISP, JPN | |
| 163 | STN13, SH3PXD2A2 | rs2067831 | Chr10:103883465 | BP, SM | EUR | |
| 164 | TCF7L25 | rs7903146 | Chr10:112998590 | BH, BMI, BP, D, L, SM | Multi | |
| 165 | C10orf823, GFRA11 | rs2218584 | Chr10:116649131 | L | Multi | |
| 166 | WDR111 | rs17101521 | Chr10:121156039 | BMI, BP, L | Multi | |
| 167 | HTRA16, TACC22 | rs2672592, rs4752700, rs17103659 | Chr10:122471234-122515087 | EUR, HISP | ||
| 168 | BET1L3, PSMD132 | rs73386640 | Chr11:203235 | JPN | ||
| 169 | TRIM54, TRIM221 | rs11601507 | Chr11:5679844 | BC, L | EUR | |
| 170 | DENND2B2, TRIM662 | LOC1027247842, TMEM9B-AS12 | rs56210063 | Chr11:8767618 | Multi | |
| 171 | SWAP708, TMEM41B5 | LOC4400282, SBF2-AS12 | rs4537761, rs360153 | Chr11:9301806-9740727 | BC, BP, D | EUR |
| 172 | CTR92 | MRVl12 | rs11042937 | Chr11:10723847 | EUR | |
| 173 | BMAL16, ARNTL2 | rs11316597 | Chr11:13274205 | BP, PSY | EUR | |
| 174 | BDNF2, LIN7C2 | LINC006782 | rs6265 | Chr11:27658369 | ALC, BMI, BP, I, L, PSY, SM | Multi |
| 175 | WT14, IMMP1L2 | rs7118294 | Chr11:32358975 | BH, BP, L | EUR | |
| 176 | HSD17B127, TTC171 | rs7116641 | Chr11:43675367 | BH, BMI, I | EUR | |
| 177 | SIPA18, RNASEH2C3 | rs2306363 | Chr11:65638129 | ALC, BMI, BP, L | EUR, JPN | |
| 178 | SERPINH117, MAP64 | AP001922.52 | rs590121, rs584961 | Chr11:75563105-75566583 | BH, BMI | EUR |
| 179 | ARHGAP4211, PGR1 | LOC1001283862 | rs633185, rs7947761 | Chr11:100722807-100753868 | ALC, BP | EUR |
| 180 | MMP132, MMP31 | rs1892971 | Chr11:102924877 | EUR | ||
| 181 | PDGFD5, DYNC2H11 | rs2839812 | Chr11:103802566 | BMI, BP | EUR, JPN | |
| 182 | CWF19L25, GUCY1A22 | rs7117780 | Chr11:107215397 | BP | EUR | |
| 183 | RDX5, FDX14 | rs10488763 | Chr11:110373636 | EUR, JPN | ||
| 184 | SIK27, CRYAB5 | rs11213945 | Chr11:111590279 | BP | Multi | |
| 185 | ZPR13, BUD132 | rs964184 | Chr11:116778201 | L | EUR, HISP | |
| 186 | VPS111, MCAM1 | rs1177562 | Chr11:119078621 | L | EUR | |
| 187 | TLCD53, ARHGEF122 | rs4938809 | Chr11:120493228 | EUR | ||
| 188 | UBASH3B6, JHY1 | rs57812455 | Chr11:122654259 | BC, BP, L | EUR | |
| 189 | ST3GAL45, TIRAP2 | GSEC3 | rs10790800 | Chr11:126392743 | L | EUR |
| 190 | C1S4, CHD42 | rs72447384 | Chr12:7071137 | I | EUR | |
| 191 | YBX36, OLR11 | rs2607903 | Chr12:10723974 | JPN | ||
| 192 | PDE3A4, AEBP22 | rs10841443 | Chr12:20067099 | BP | EUR | |
| 193 | ZCRB17, PPHLN17 | rs12820154 | Chr12:42389305 | BP | Multi | |
| 194 | ZNF6414, CCDC1842 | rs1859440 | Chr12:48033546 | BC, BP | EUR | |
| 195 | SMUG13, HOXC43 | FLJ128252, RP11-834C11.42 | rs1133773, rs75160195 | Chr12:54102435-54127810 | EUR, JPN | |
| 196 | COQ10A6, STAT24 | rs143405823 | Chr12:56329935 | BH, L, PSY | Multi | |
| 197 | LRP17, STAT65 | rs11172113 | Chr12:57133500 | BH, BMI, BP, L, PSY | EUR | |
| 198 | ATP2B15, GALNT43 | AC010201.22 | rs2681472 | Chr12:89615182 | ALC, BC, BMI, BP, PSY | EUR, JPN |
| 199 | FGD67, VEZT3 | rs11107903 | Chr12:95114195 | BC | EUR, JPN | |
| 200 | CORO1C4, SELPLG1 | rs1558802 | Chr12:108642472 | L, PSY | Multi | |
| 201 | ATXN24, SH2B32 | rs10774625, rs11066015 | Chr12:111472415-111730205 | ALC, BC, BMI, BP, I, L, PSY, SM | EUR, JPN | |
| 202 | TBX34, TBX51 | rs34606058 | Chr12:114915563 | BC, BH, BP | EUR, Multi | |
| 203 | KSR24, FBXW81 | rs11830157 | Chr12:117827636 | EUR | ||
| 204 | HNF1A6, C12orf435 | RNU4-22, HNF1A-AS12 | rs2244608 | Chr12:120979185 | ALC, BC, I, L, PSY | EUR |
| 205 | ZNF6646, CCDC9211 | rs7133378 | Chr12:123924955 | BMI, D, L, PSY, SM | EUR | |
| 206 | SCARB111, UBC2 | rs7485656, rs7296737 | Chr12:124831101-124852410 | I, L | BLK, EUR | |
| 207 | ZNF2682, ZNF1402 | AC131212.32 | rs36127550 | Chr12:133203723 | BMI, BP | Multi |
| 208 | FLT15, SLC46A32 | rs17086617, rs74412485 | Chr13:28388549-28468295 | BH, BP | EUR, JPN | |
| 209 | PDS5B8, N4BP2L25 | rs7991314 | Chr13:32551937 | BC, BH, BMI, SM | EUR | |
| 210 | DOCK93, SLC15A11 | rs8000794 | Chr13:98782556 | EUR | ||
| 211 | COL4A119, COL4A212 | rs11617955, rs3783113, rs11619113, rs4773141, rs9515203, rs7333991 | Chr13:110165755-110456089 | BP, C, L | BLK, EUR, JPN | |
| 212 | F102, MCF2L1 | rs4907571 | Chr13:112964182 | BP | EUR | |
| 213 | ARID4A2, ARMH42 | PSMA3-AS12 | rs2145598 | Chr14:58327283 | L | EUR |
| 214 | EIF2B27, MLH36 | rs10131894 | Chr14:74980176 | EUR | ||
| 215 | ITPK14, ATXN32 | rs1956028 | Chr14:93040852 | BC | Multi | |
| 216 | SERPINA14, DDX241 | rs112635299 | Chr14:94371805 | ALC, BH, BMI, BP, I, L, PSY | EUR | |
| 217 | HHIPL111, CYP46A15 | AL160313.12 | rs36033161, rs2895811 | Chr14:99657150-99667605 | BP | EUR, JPN |
| 218 | EXD12, KNSTRN2 | OIP5-AS13 | rs3959554 | Chr15:41151726 | BMI, BP | EUR |
| 219 | NEDD43, PIGB3 | AC018926.12 | rs11633313 | Chr15:55843511 | Multi | |
| 220 | LIPC5, ALDH1A25 | HSP90AB4P2 | rs588136 | Chr15:58438299 | L, PSY | EUR |
| 221 | TRIP45, RBPMS23 | rs6494488 | Chr15:64732005 | L | EUR | |
| 222 | SMAD37, SMAD63 | IQCH-AS12 | rs56062135 | Chr15:67163292 | EUR, HISP | |
| 223 | ADAMTS713, CTSH10 | AC090607.22 | rs7177201, rs7173743, rs112238647, rs7174727 | Chr15:78759363-78858944 | BH, BP,C, SM | EUR, HISP, JPN |
| 224 | CFAP1613, MESD2 | rs2683260 | Chr15:81093211 | EUR | ||
| 225 | AKAP133, KLHL253 | LOC1019296792, AC044860.12 | rs7177107 | Chr15:85580133 | BC | EUR |
| 226 | MFGE85, ABHD24 | rs1807214 | Chr15:89022026 | BP | EUR, JPN | |
| 227 | FES9, FURIN6 | rs7183988 | Chr15:90885359 | BP, L, PSY | BLK, EUR, JPN | |
| 228 | NR2F22 | rs17581137 | Chr15:95603185 | EUR | ||
| 229 | MYH115, NOMO12 | AC138969.12 | rs12691049 | Chr16:15815656 | AF, BP | EUR |
| 230 | FTO5, AKTIP1 | rs1421085 | Chr16:53767042 | ALC, BH, BMI, BP, D, I, L, PSY, SM | BLK, EUR | |
| 231 | CETP6, HERPUD13 | rs12446515 | Chr16:56953103 | AF, BMI, BP, C, L | EUR | |
| 232 | NOB13, NFAT52 | PDXDC2P3 | rs9302604 | Chr16:69542991 | BP, PSY, SM | EUR |
| 233 | DHODH7, DHX383 | rs12445401 | Chr16:72114520 | ALC, L, PSY | EUR | |
| 234 | CFDP16, TMEM170A5 | AC009054.22 | rs8046696 | Chr16:75408245 | ALC, BC, BH, BP, L, SM | EUR, JPN |
| 235 | CMIP8, PLCG26 | AC009148.12 | rs10493891, rs7189462 | Chr16:81477137-81874262 | BP, L | EUR |
| 236 | CDH134, OSGIN11 | rs7500448 | Chr16:83012185 | BP | EUR | |
| 237 | rs55880988 | Chr16:88479845 | L, I | Multi | ||
| 238 | SMG65, SRR3 | rs4790881 | Chr17:2165638 | L | EUR | |
| 239 | TOM1L27, DRC37 | CCDC144BP2, USP32P22 | rs12936927 | Chr17:17823651 | ALC, BMI, PSY, SM | EUR, HISP |
| 240 | SSH28, CORO65 | SH3GL1P22 | rs11080107 | Chr17:29611406 | BH, BMI | EUR |
| 241 | COPRS2, NF11 | rs76954792 | Chr17:31706495 | BP | EUR | |
| 242 | FBXL207, MED15 | rs1077715 | Chr17:39527295 | ALC, BC, L, SM | EUR | |
| 243 | CAVIN18, DHX586 | NBR24, AC099811.53 | rs12952244, rs8068844, rs9912587 | Chr17:42118063-43021069 | BMI, D, I, L | EUR, Multi |
| 244 | ARHGAP275, GOSR25 | LINC022102, NSFP12 | rs5026246, rs1706731, rs1881193, rs1881194, rs2532235, rs2696671, rs17608766 | Chr17:45495530-46935905 | AF, BP, PSY | EUR, HISP |
| 245 | SNF810, ATP5MC110 | SUMO2P174, LOC1053718142 | rs46522, rs4643373, rs5820757 | Chr17:48911235-49340581 | ALC, BH, BMI, BP, I, L, PSY | EUR |
| 246 | BCAS34, PPM1D2 | RPL12P382 | rs11655024 | Chr17:61155004 | BMI, BP | EUR, JPN |
| 247 | PECAM15, ERN12 | AC046185.32 | rs11079536 | Chr17:64315043 | BC, BP | EUR, JPN |
| 248 | WIPI16, BPTF3 | rs2909217 | Chr17:68467844 | L | EUR | |
| 249 | TRIM4710, UNC13D8 | ATF4P33 | rs2410859, rs78532451 | Chr17:75845204-75883011 | BP, L | EUR, Multi |
| 250 | rs112735431 | Chr17:80385145 | JPN | |||
| 251 | CTAGE13, GATA63 | rs9951447 | Chr18:22429728 | EUR, JPN | ||
| 252 | NPC14, RMC14 | rs1623003 | Chr18:23585199 | ALC, BP, L, SM | EUR | |
| 253 | rs4327120 | Chr18:38953012 | SM | Multi | ||
| 254 | LIPG5, SMAD74 | AC022919.12 | rs9945890, rs12965923 | Chr18:48989546-49687312 | L | EUR |
| 255 | LMAN16, MC4R5 | rs11663411, rs476828 | Chr18:59293278-60185354 | ALC, BH, BMI, BP, D, I, L, PSY, SM | EUR | |
| 256 | rs150651184 | Chr19:282148 | BLK | |||
| 257 | DOT1L6, PLEKHJ15 | AC006130.32 | rs191615952 | Chr19:2236098 | BH, BP | Multi |
| 258 | KDM4B5, TICAM12 | rs12185519 | Chr19:4967728 | BH, BMI, L | Multi | |
| 259 | ANGPTL46, MAP2K71 | rs116843064 | Chr19:8364439 | ALC, BMI, L | EUR | |
| 260 | LDLR18, SMARCA46 | ZNF433-AS14, AC008770.32 | rs55997232, rs10422256, rs72658867, rs34774090, rs167479 | Chr19:11077441-11416089 | ALC, BC, BP, L, PSY | BLK, EUR, HISP, JPN |
| 261 | CYP4F22, EPS15L12 | rs77420750 | Chr19:15868934 | EUR | ||
| 262 | KLF26, AP1M12 | rs10420373 | Chr19:16319866 | I | JPN | |
| 263 | MYO9B6, HAUS85 | rs7246865 | Chr19:17108295 | BC, BMI, BP | EUR | |
| 264 | MAP1S9, FCHO16 | rs10410487 | Chr19:17718799 | BC, BH, I | EUR | |
| 265 | ELL2, ISYNA12 | rs78030362 | Chr19:18464383 | BC, BMI, BP | EUR | |
| 266 | ZNF1007, ZNF4933 | rs8102320 | Chr19:21754083 | BP | Multi | |
| 267 | ZNF5078, DPY19L31 | rs12976411 | Chr19:32391114 | EUR | ||
| 268 | EIF3K2, ZFP302 | ZNF7815, LINC015352 | rs73025613 | Chr19:37843721 | Multi | |
| 269 | TGFB115, B9D26 | AC011462.53 | rs60315715, rs11466359, rs1800469 | Chr19:41284182-41354391 | BC, BP | EUR, HISP, JPN |
| 270 | APOE12, APOC18 | rs429358, rs7412, rs183657985 | Chr19:44908684-45239240 | BC, BH, BP, C, I, L | BLK, EUR, JPN | |
| 271 | RSPH6A4, DMWD4 | PPP5D1P2, GEMIN7-AS12 | rs8108474 | Chr19:45798221 | BH, BP, L | EUR |
| 272 | RRBP16, DSTN3 | rs1132274 | Chr20:17615510 | BH, L | EUR, Multi | |
| 273 | ITCH7, NCOA64 | FER1L42 | rs6088595 | Chr20:34770696 | BC, BH, BMI, D, I, L, SM | EUR |
| 274 | PROCR7, EDEM24 | NORAD2 | rs17406518 | Chr20:35211373 | BC | EUR |
| 275 | MAFB4 | rs2207132 | Chr20:40513876 | BMI, I, L | EUR | |
| 276 | PLCG16, TOP14 | rs6102343 | Chr20:41295639 | BMI, L | EUR | |
| 277 | PLTP3, CD403 | rs8124182 | Chr20:45980262 | L | EUR | |
| 278 | PREX15, ARFGEF22 | RN7SL197P2 | rs2008614 | Chr20:48816613 | BP | EUR |
| 279 | rs4809766 | Chr20:49942802 | EUR | |||
| 280 | GNAS3, ZNF8312 | APCDD1L-DT2, LINC017112 | rs6026739 | Chr20:59164414 | BH, BP | EUR |
| 281 | rs151206628 | Chr21:10592292 | HISP | |||
| 282 | NRIP13, HSPA131 | rs2064040 | Chr21:15212021 | Multi | ||
| 283 | MAP3K7CL5, BACH14 | LINC001893 | rs2832227 | Chr21:29160755 | EUR | |
| 284 | SLC5A36, MRPS63 | AP000318.22 | rs28451064, rs149487184 | Chr21:34221526-34233562 | BC, BMI, BP | EUR |
| 285 | ARVCF8, TANGO25 | DGCR112 | rs4819811, rs71313931 | Chr22:19621733-19972661 | BC, BP, I | EUR |
| 286 | SUSD27, GGT56 | POM121L9P3, GGTLC4P2 | rs12484557, rs5760309 | Chr22:24159893-24281863 | EUR | |
| 287 | OSM4, LIF3 | rs6006426 | Chr22:30273894 | EUR | ||
| 288 | FBXO71 | rs137525 | Chr22:32900153 | Multi | ||
| 289 | SCUBE13, TTLL122 | rs139012 | Chr22:43227966 | EUR | ||
| 290 | PNPLA32, PRR52 | rs738408 | Chr22:43928850 | ALC, BC, L | Multi | |
| 291 | TBL1X | rs5934659 | ChrX:9610064 | EUR | ||
| 292 | OPHN11, AR1 | rs1410127 | ChrX:68060539 | Multi | ||
| 293 | CYSLTR11, TAF9B1 | rs398484 | ChrX:78343972 | Multi | ||
| 294 | VDAC1P1 | rs2066280 | ChrX:80922131 | EUR | ||
| 295 | SETP4 | rs2342572 | ChrX:84814364 | Multi | ||
| 296 | CHRDL11, RGAG11 | rs7884019 | ChrX:110566261 | EUR | ||
| 297 | MAP7D31, FHL11 | rs5929743 | ChrX:136236818 | EUR | ||
| 298 | ARHGEF61 | RP11-308D16.21 | rs5975828 | ChrX:136904390 | EUR | |
| 299 | DNASE1L11, ATP6AP11 | rs147967693 | ChrX:154410919 | Multi |
| Locus ID | Genes (protein) | Genes (RNA) | Index rsIDs | Coordinates (hg38) | Traits | Ethnicity |
|---|---|---|---|---|---|---|
| 1 | SKI4, MORN13 | rs2843152 | Chr1:2314131 | L | EUR | |
| 2 | PRDM167, ACTRT22 | rs7413494, rs2493298 | Chr1:3069321-3409348 | BP | EUR | |
| 3 | DHDDS7, RPS6KA13 | DHDDS-AS12 | rs12046497 | Chr1:26521149 | BH, L | Multi |
| 4 | ARID1A7, KDF12 | LOC1019287282 | rs79598313 | Chr1:26958422 | ALC, BH, BP, I, L | EUR |
| 5 | FHL37, INPP5B6 | rs61776719 | Chr1:37995647 | BH, BMI, BP | EUR | |
| 6 | ZMYND125, FOXO64 | rs12047439, rs2152314 | Chr1:41343968-42480791 | L, BP | Multi | |
| 7 | PCSK916, USP244 | rs34232196, rs11591147, rs151193009, rs472495 | Chr1:55023869-55055640 | ALC, BC, L, PSY | EUR, JPN | |
| 8 | PLPP310, FGGY8 | LINC013584, AC093425.12 | rs11206803, rs56170783, rs71646019, rs12733512 | Chr1:56411837-59181306 | BH, BMI, BP, SM | EUR, HISP, JPN, Multi |
| 9 | PSRC18, CELSR26 | rs12740374, rs655246 | Chr1:109274968-109289661 | ALC, BC, BH, BP, I, L, PSY | BLK, EUR, HISP | |
| 10 | PHTF13, MAGI32 | rs1230666 | Chr1:113630788 | EUR | ||
| 11 | NGF8, TSPAN22 | rs11806316, rs61797068 | Chr1:115210861-115359893 | EUR | ||
| 12 | ECM16, MTMR115 | ADAMTSL4-AS12, AL356356.12 | rs67807996, rs11585169 | Chr1:150023307-150599561 | BH, BMI, BP, I, SM | EUR |
| 13 | TDRKH9, RIIAD13 | TDRKH-AS13 | rs11810571 | Chr1:151789832 | PSY | EUR |
| 14 | IL6R5, TPM32 | rs6686750 | Chr1:154447367 | I | EUR | |
| 15 | NME79, ATP1B15 | rs61806987 | Chr1:169345595 | BH | EUR | |
| 16 | KIAA00405, TNN1 | rs4650716 | Chr1:175159575 | EUR | ||
| 17 | DENND1B4, CRB12 | rs12022672 | Chr1:197618876 | SM | Multi | |
| 18 | DDX597, CAMSAP24 | DDX59-AS12 | rs6700559 | Chr1:200676945 | EUR | |
| 19 | LMOD17, IPO94 | IPO9-AS12 | rs2820315 | Chr1:201903136 | BH, BMI, BP, L, PSY | EUR |
| 20 | DSTYK3, NUAK23 | AC119673.22 | rs12048743 | Chr1:205145745 | L | Multi |
| 21 | HHAT7, SYT142 | RNU5A-8P2 | rs60154123 | Chr1:210295654 | EUR | |
| 22 | MIA36, AIDA4 | MIA3-AS12 | rs17163363 | Chr1:222655362 | BP | EUR, HISP, JPN |
| 23 | AGT5, COG21 | rs699 | Chr1:230710048 | BP | EUR | |
| 24 | COLEC112 | RNASEH1-DT3 | rs56236159 | Chr2:3588888 | BC, BP, I, L | Multi |
| 25 | IAH13, ADAM173 | rs2715879 | Chr2:9375422 | EUR | ||
| 26 | OSR13, TTC322 | rs16986953 | Chr2:19742712 | EUR | ||
| 27 | APOB5, TDRD152 | rs13306206, rs515135 | Chr2:21019859-21063185 | BC, L, PSY | EUR, JPN | |
| 28 | ABCG810, ABCG56 | rs4245791, rs76866386 | Chr2:43847292-43848344 | BC, I, L, PSY | EUR | |
| 29 | PRKCE7, SRBD11 | rs582384 | Chr2:45669298 | BP | EUR | |
| 30 | BCL11A4, REL1 | MIR4432HG2 | rs243071 | Chr2:60391893 | EUR | |
| 31 | GFPT15, NFU13 | rs4346430 | Chr2:69441670 | Multi | ||
| 32 | GGCX7, VAMP86 | PARTICL2 | rs10176176 | Chr2:85534925 | BMI, L | EUR, JPN |
| 33 | IL1F105, PSD43 | rs6759676 | Chr2:113078771 | BP, I | EUR | |
| 34 | SAP1305, WDR331 | rs114192718 | Chr2:128028089 | EUR | ||
| 35 | ARHGAP155, KYNU1 | rs4662330 | Chr2:143428906 | BP | EUR | |
| 36 | ZEB212, GTDC13 | LINC014123, AC009951.42 | rs6740731, rs17408036, rs10928241 | Chr2:144513025-145073861 | BP, SM | EUR, HISP |
| 37 | ACVR2A4, MBD52 | rs35611688 | Chr2:147620292 | EUR | ||
| 38 | FIGN5, GRB141 | rs10930115 | Chr2:164073872 | BP, PSY, SM | EUR, JPN | |
| 39 | TFPI4, CALCRL3 | rs840616 | Chr2:187331742 | BMI, BP, L | EUR | |
| 40 | NBEAL17, CARF7 | rs148812085 | Chr2:203012510 | BMI, BP, L, PSY, SM | EUR, HISP | |
| 41 | FN17, ATIC2 | rs1250247 | Chr2:215434906 | BH, BMI, BP, L | EUR | |
| 42 | TNS15, TMBIM12 | rs2161967 | Chr2:217815806 | BP, L | EUR, HISP | |
| 43 | IRS15, NYAP22 | rs952227 | Chr2:226197364 | ALC, BMI, BP, D, L, SM | EUR | |
| 44 | PID15, DNER3 | rs4140748 | Chr2:229140789 | L | EUR, JPN | |
| 45 | GIGYF27, SNORC4 | rs283485 | Chr2:232780981 | BH, BMI | EUR | |
| 46 | COL6A34, RBM441 | rs11677932 | Chr2:237315312 | BP | EUR | |
| 47 | FGD55, NR2C21 | rs34991912 | Chr3:14884844 | BH, BP | EUR, JPN | |
| 48 | ZNF5897, CDC25A6 | AC134772.12 | rs7633770, rs7617773 | Chr3:46647072-48152025 | BH, BP | EUR |
| 49 | IP6K23, IHO13 | AC121247.12 | rs34759087 | Chr3:49124851 | BH, BMI, PSY | EUR |
| 50 | ITIH43, NISCH2 | rs77347777 | Chr3:52814191 | BMI, BP | EUR | |
| 51 | ITGB55, UMPS4 | rs17843797 | Chr3:124734175 | BMI | EUR | |
| 52 | DNAJC135, NPHP34 | rs10512861 | Chr3:132539117 | I, L | EUR | |
| 53 | PCCB7, PPP2R3A5 | NCK1-DT3 | rs34330586 | Chr3:136081567 | BMI, I, L | EUR |
| 54 | MRAS5, PIK3CB3 | rs185244 | Chr3:138374047 | BMI, BP, L | EUR | |
| 55 | ATP1B34, PLS12 | rs7622417 | Chr3:141907157 | BMI, L | Multi | |
| 56 | ARHGEF268, DHX367 | ARHGEF26-AS13 | rs357494 | Chr3:154219964 | BMI, BP, L | EUR, HISP |
| 57 | CCNL14, TIPARP4 | rs4266144 | Chr3:157134803 | BH, BMI | EUR | |
| 58 | MECOM2, SEC621 | rs16853198 | Chr3:169122391 | BP | Multi | |
| 59 | FNDC3B2, ECT21 | rs34229028 | Chr3:172399666 | EUR | ||
| 60 | rs73070813 | Chr3:187168807 | EUR | |||
| 61 | HGFAC2 | rs16844401 | Chr4:3447925 | BC, BH, L | EUR | |
| 62 | GNPDA21 | rs10938398 | Chr4:45184122 | BMI, I, L | Multi | |
| 63 | FIP1L12, LNX11 | rs17083333 | Chr4:53705899 | EUR | ||
| 64 | REST9, SPINK22 | rs781663 | Chr4:56915588 | BC, BH, I, L | EUR, JPN | |
| 65 | SHROOM35, CCDC1582 | rs12500824 | Chr4:76495474 | L, BP | EUR | |
| 66 | ANTXR22, PRDM81 | rs10857147 | Chr4:80259918 | L, BP | EUR, HISP | |
| 67 | RASGEF1B4, PRKG21 | rs36002015 | Chr4:81704567 | EUR | ||
| 68 | UNC5C2, BMPR1B2 | rs3775058 | Chr4:95196220 | EUR | ||
| 69 | MAD2L15, PDE5A4 | rs7678555 | Chr4:119988346 | BP | EUR | |
| 70 | PCDH182 | rs7439567 | Chr4:137543688 | BP | Multi | |
| 71 | ZNF8275, MMAA1 | rs13124853 | Chr4:145863622 | BP, L | EUR | |
| 72 | EDNRA14, ARHGAP103 | AC093908.12 | rs13120678, rs1878406, rs6841581 | Chr4:147352245-147480038 | BC, BP | EUR, JPN |
| 73 | GUCY1A110, MAP92 | RP11-588K22.22 | rs7440763, rs3796587 | Chr4:155512368-155716921 | ALC, BP, L | EUR, HISP, JPN |
| 74 | PALLD4, DDX60L1 | rs869396 | Chr4:168766849 | BP | EUR | |
| 75 | IRX13 | rs10041378 | Chr5:4094052 | JPN | ||
| 76 | SEMA5A7, TAS2R11 | SNHG185 | rs17263917 | Chr5:9552226 | BC | EUR |
| 77 | BASP12 | rs1032763 | Chr5:17118821 | Multi | ||
| 78 | ITGA15, PELO1 | rs4074793 | Chr5:52897294 | L, I | EUR, Multi | |
| 79 | ARL152, HSPB31 | AC027329.12 | rs7719168 | Chr5:53996560 | BP, L | EUR |
| 80 | MAP3K14, IL6ST2 | rs62362364, rs3936510 | Chr5:56145744-56565039 | BMI, BP, L, SM | EUR | |
| 81 | ANKDD1B7, HMGCR6 | rs12916 | Chr5:75360714 | BC, BMI, L, PSY | EUR | |
| 82 | LNPEP7, ERAP27 | AC009126.12 | rs27660 | Chr5:97012071 | BP, I, L, PSY | Multi |
| 83 | FER11, FBXL174 | LINC010233 | rs288173, rs112949822 | Chr5:108016593-108749489 | BH, BMI, BP, I | EUR, Multi |
| 84 | NREP1, WDR361 | rs256277 | Chr5:112071113 | Multi | ||
| 85 | DMXL12, TNFAIP81 | rs13169691 | Chr5:119112584 | EUR | ||
| 86 | SRFBP15, LOX3 | AC008568.12 | rs4345341 | Chr5:121943056 | BP | EUR |
| 87 | PRDM63, SNX241 | rs73796819 | Chr5:123084735 | Multi | ||
| 88 | FBN28, SLC27A62 | rs6883598 | Chr5:128590497 | BP | EUR | |
| 89 | IRF14, RAD504 | rs10477741, rs2706399 | Chr5:132459618-132532010 | L | EUR | |
| 90 | FGF16, SPRY42 | rs249760, rs3776307 | Chr5:142536127-143114600 | L | EUR | |
| 91 | SGCD4, MED72 | rs39929 | Chr5:156696640 | L, BP | Multi | |
| 92 | ADAM195, FNDC91 | rs11465228 | Chr5:157575687 | Multi | ||
| 93 | FOXC15, GMDS3 | rs72836800 | Chr6:1617092 | BH | EUR | |
| 94 | BLOC1S51, TXNDC51 | rs6597292 | Chr6:7975026 | Multi | ||
| 95 | PHACTR16, GFOD12 | rs9349379 | Chr6:12903725 | ALC, BC, BP, C | EUR, HISP, JPN | |
| 96 | HDGFL14, PRL1 | rs6909752 | Chr6:22612400 | AF | EUR | |
| 97 | HLA-A4, HLA-G3 | HLA-F-AS15, HLA-H3 | rs29266 | Chr6:29607502 | L | EUR |
| 98 | MICA4, PPP1R104 | rs139141104, rs9266631 | Chr6:31021244-31379121 | L | EUR, JPN | |
| 99 | CLIC13, DDAH23 | SNHG322, LOC1005075472 | rs3130683 | Chr6:31920590 | BH, BMI, BP, L, PSY | EUR |
| 100 | LEMD23, UQCC23 | rs62397561 | Chr6:33816328 | Multi | ||
| 101 | BLTP3A9, SNRPC8 | ILRUN-AS12 | rs9469899, rs17609940 | Chr6:34825347-35067023 | BH, BMI, L, PSY | EUR |
| 102 | CDKN1A5, ETV72 | DINOL2 | rs1321309 | Chr6:36670859 | BP, I, L | EUR |
| 103 | KCNK510, SAYSD11 | rs733701, rs10947789 | Chr6:39204086-39207146 | BP | EUR | |
| 104 | PEX63, PTK71 | rs1034246 | Chr6:43100632 | EUR | ||
| 105 | VEGFA4, MRPS18A1 | rs6905288, rs149372871 | Chr6:43791136-44080314 | ALC, BMI, BP, L, PSY | EUR, HISP, JPN | |
| 106 | RAB2310, PRIM26 | LOC1005061884, RP11-203B9.42 | rs11752218, rs9367716 | Chr6:57280764-57295774 | EUR | |
| 107 | CD1096 | CD109-AS12 | rs79717953, rs56171536 | Chr6:73695048-73706145 | JPN, Multi | |
| 108 | TENT5A4, IBTK1 | LINC0254`22 | rs9361867 | Chr6:81886242 | EUR | |
| 109 | UFL16, FHL55 | rs2983896 | Chr6:96581995 | BP | EUR | |
| 110 | CENPW8, TRMT112 | rs35510806 | Chr6:126357186 | BH, I, L, PSY | EUR | |
| 111 | TCF2113, SLC2A127 | TARID3 | rs6919211, rs2327426, rs2492304 | Chr6:133678730-134057013 | BH, BP | EUR, HISP, JPN |
| 112 | PLEKHG14, MTHFD1L2 | RAET1K2 | rs17080093 | Chr6:150676304 | BP | EUR |
| 113 | PLG29, LPA19 | LPAL22 | rs6932293, rs186696265, rs56195640, rs192425087, rs28867400, rs932631509, rs10455872, rs73596816, rs79018195, rs1998043, rs4252120, rs184278183 | Chr6:160114846-160756724 | BC, BMI, BP, C, L, PSY | BLK, EUR, HISP, JPN |
| 114 | MAD1L16, ELFN12 | rs62435159 | Chr7:1871645 | BMI | EUR | |
| 115 | DAGLB8, RAC17 | rs10951983 | Chr7:6406396 | I, L | EUR | |
| 116 | TMEM106B7, THSD7A2 | rs11509880 | Chr7:12222285 | L, PSY | EUR | |
| 117 | ETV14 | rs2073533 | Chr7:13990114 | EUR | ||
| 118 | TWIST15, HDAC92 | rs2107595 | Chr7:19009765 | BP | EUR, JPN | |
| 119 | MACC12, ITGB82 | rs10486389 | Chr7:20260793 | EUR | ||
| 120 | TBX208, DPY19L12 | DPY19L2P12 | rs2215614 | Chr7:35237481 | BH | EUR |
| 121 | CCM25, TBRG41 | rs2107732 | Chr7:45038379 | BP | EUR | |
| 122 | SEMA3C5 | rs917191 | Chr7:80941555 | BMI | Multi | |
| 123 | ZKSCAN16, AP4M15 | STAG3L5P2 | rs35146811 | Chr7:100123371 | BH, BP, L | EUR |
| 124 | COG59, BCAP298 | WBP1LP23, RPL7P322 | rs12112877, rs10953541 | Chr7:107300879-107604100 | L, PSY, SM | EUR |
| 125 | CFTR3, ASZ12 | rs13222797 | Chr7:117459992 | BMI, L | EUR | |
| 126 | ZC3HC15, KLHDC102 | rs11556924 | Chr7:130023656 | BC, BP, I, L, PSY, SM | EUR | |
| 127 | PARP122, HIPK21 | rs5887949 | Chr7:140060741 | BH | EUR | |
| 128 | NOS35, KCNH21 | rs3918226 | Chr7:150993088 | ALC, BC, BP, L, SM | EUR | |
| 129 | MSR13, CNOT71 | rs1480146 | Chr8:16272064 | Multi | ||
| 130 | NAT23, PCM13 | rs4646249 | Chr8:18402921 | L | EUR | |
| 131 | LPL14, CSGALNACT13 | rs894211, rs66778572, rs268 | Chr8:19956018-20058609 | ALC, BC, BH, BMI, BP, I, L, PSY, SM | BLK, EUR, HISP | |
| 132 | BMP15, PHYHIP2 | rs56408342 | Chr8:22190977 | EUR | ||
| 133 | DOCK56, KCTD92 | rs1510758 | Chr8:25204291 | EUR | ||
| 134 | PPP2R2A2 | rs6557894 | Chr8:26229397 | BH | EUR | |
| 135 | RP15 | rs9298506 | Chr8:54524964 | ALC, BH, I, L | Multi | |
| 136 | C8orf342 | rs2380472 | Chr8:68519476 | JPN | ||
| 137 | GEM6, CDH172 | rs34917849 | Chr8:94266079 | BP | EUR | |
| 138 | NDUFAF65, TP53INP11 | rs3019173 | Chr8:95008746 | Multi | ||
| 139 | rs10093110 | Chr8:105553186 | EUR | |||
| 140 | TRIB14, NSMCE21 | rs2001846 | Chr8:125466208 | ALC, BMI, BP, D, I, L | EUR, HISP | |
| 141 | NFIB2, MPDZ1 | LURAP1L-AS12 | rs10961206 | Chr9:13724052 | EUR | |
| 142 | ACER27, RPS61 | AL391834.12 | rs10811183 | Chr9:19436057 | L | EUR |
| 143 | CDKN2A13, CDKN2B11 | ERVFRD-36, MIR31HG4 | rs11523031, rs2891168, rs6475608, rs76959412 | Chr9:21843843-22101703 | BC, BP, C, L | BLK, EUR, HISP, JPN |
| 144 | WNK23, CARD192 | rs4617217 | Chr9:93226686 | Multi | ||
| 145 | ABCA12, NIPSNAP3B2 | rs35093463 | Chr9:104823957 | L | JPN | |
| 146 | KLF44, RAD23B1 | rs1967604 | Chr9:107768043 | BC, L | EUR | |
| 147 | SVEP14 | rs111245230 | Chr9:110407495 | BC, BP | EUR | |
| 148 | ASTN22 | rs7873551 | Chr9:116482848 | BH, PSY | Multi | |
| 149 | PHF195, C54 | rs41312891 | Chr9:120964472 | BH | EUR, Multi | |
| 150 | DAB2IP5, STOM1 | CUTALP2 | rs885150 | Chr9:121657894 | BC, BH, BMI, L | EUR |
| 151 | ABO6, CEL3 | rs651007 | Chr9:133278431 | BC, BP, D, I, L, PSY | BLK, EUR, HISP | |
| 152 | CAMK1D4, CDC1232 | rs17566555 | Chr10:12233948 | PSY | EUR | |
| 153 | ARHGAP214, GPR1581 | rs7077962 | Chr10:24765745 | Multi | ||
| 154 | JCAD7, SVIL1 | rs9337951 | Chr10:30028144 | BP, SM | EUR, HISP | |
| 155 | ZEB16, ARHGAP121 | ZEB1-AS15, PPIAP312 | rs161231 | Chr10:31466991 | L | Multi |
| 156 | CXCL1212, RASGEF1A2 | FRG1JP2 | rs1870634, rs494207, rs527052 | Chr10:43985363-44245808 | C | BLK, EUR, HISP |
| 157 | BICC17, TFAM1 | rs10740732 | Chr10:58593128 | BP | Multi | |
| 158 | TSPAN147, SH2D4B2 | LOC1019295742 | rs17680741 | Chr10:80491758 | EUR | |
| 159 | LIPA6, IFIT11 | rs1051338 | Chr10:89247603 | I, L | EUR, JPN | |
| 160 | PLCE112, SLC35G12 | rs1223583, rs55753709 | Chr10:93996743-94269413 | BMI, BP, L, PSY | EUR | |
| 161 | LOXL49, R3HCC1L3 | rs884811 | Chr10:98164006 | BH, BP | EUR | |
| 162 | CNNM28, BORCS75 | rs77787671 | Chr10:103016448 | BMI, BP, PSY | EUR, HISP, JPN | |
| 163 | STN13, SH3PXD2A2 | rs2067831 | Chr10:103883465 | BP, SM | EUR | |
| 164 | TCF7L25 | rs7903146 | Chr10:112998590 | BH, BMI, BP, D, L, SM | Multi | |
| 165 | C10orf823, GFRA11 | rs2218584 | Chr10:116649131 | L | Multi | |
| 166 | WDR111 | rs17101521 | Chr10:121156039 | BMI, BP, L | Multi | |
| 167 | HTRA16, TACC22 | rs2672592, rs4752700, rs17103659 | Chr10:122471234-122515087 | EUR, HISP | ||
| 168 | BET1L3, PSMD132 | rs73386640 | Chr11:203235 | JPN | ||
| 169 | TRIM54, TRIM221 | rs11601507 | Chr11:5679844 | BC, L | EUR | |
| 170 | DENND2B2, TRIM662 | LOC1027247842, TMEM9B-AS12 | rs56210063 | Chr11:8767618 | Multi | |
| 171 | SWAP708, TMEM41B5 | LOC4400282, SBF2-AS12 | rs4537761, rs360153 | Chr11:9301806-9740727 | BC, BP, D | EUR |
| 172 | CTR92 | MRVl12 | rs11042937 | Chr11:10723847 | EUR | |
| 173 | BMAL16, ARNTL2 | rs11316597 | Chr11:13274205 | BP, PSY | EUR | |
| 174 | BDNF2, LIN7C2 | LINC006782 | rs6265 | Chr11:27658369 | ALC, BMI, BP, I, L, PSY, SM | Multi |
| 175 | WT14, IMMP1L2 | rs7118294 | Chr11:32358975 | BH, BP, L | EUR | |
| 176 | HSD17B127, TTC171 | rs7116641 | Chr11:43675367 | BH, BMI, I | EUR | |
| 177 | SIPA18, RNASEH2C3 | rs2306363 | Chr11:65638129 | ALC, BMI, BP, L | EUR, JPN | |
| 178 | SERPINH117, MAP64 | AP001922.52 | rs590121, rs584961 | Chr11:75563105-75566583 | BH, BMI | EUR |
| 179 | ARHGAP4211, PGR1 | LOC1001283862 | rs633185, rs7947761 | Chr11:100722807-100753868 | ALC, BP | EUR |
| 180 | MMP132, MMP31 | rs1892971 | Chr11:102924877 | EUR | ||
| 181 | PDGFD5, DYNC2H11 | rs2839812 | Chr11:103802566 | BMI, BP | EUR, JPN | |
| 182 | CWF19L25, GUCY1A22 | rs7117780 | Chr11:107215397 | BP | EUR | |
| 183 | RDX5, FDX14 | rs10488763 | Chr11:110373636 | EUR, JPN | ||
| 184 | SIK27, CRYAB5 | rs11213945 | Chr11:111590279 | BP | Multi | |
| 185 | ZPR13, BUD132 | rs964184 | Chr11:116778201 | L | EUR, HISP | |
| 186 | VPS111, MCAM1 | rs1177562 | Chr11:119078621 | L | EUR | |
| 187 | TLCD53, ARHGEF122 | rs4938809 | Chr11:120493228 | EUR | ||
| 188 | UBASH3B6, JHY1 | rs57812455 | Chr11:122654259 | BC, BP, L | EUR | |
| 189 | ST3GAL45, TIRAP2 | GSEC3 | rs10790800 | Chr11:126392743 | L | EUR |
| 190 | C1S4, CHD42 | rs72447384 | Chr12:7071137 | I | EUR | |
| 191 | YBX36, OLR11 | rs2607903 | Chr12:10723974 | JPN | ||
| 192 | PDE3A4, AEBP22 | rs10841443 | Chr12:20067099 | BP | EUR | |
| 193 | ZCRB17, PPHLN17 | rs12820154 | Chr12:42389305 | BP | Multi | |
| 194 | ZNF6414, CCDC1842 | rs1859440 | Chr12:48033546 | BC, BP | EUR | |
| 195 | SMUG13, HOXC43 | FLJ128252, RP11-834C11.42 | rs1133773, rs75160195 | Chr12:54102435-54127810 | EUR, JPN | |
| 196 | COQ10A6, STAT24 | rs143405823 | Chr12:56329935 | BH, L, PSY | Multi | |
| 197 | LRP17, STAT65 | rs11172113 | Chr12:57133500 | BH, BMI, BP, L, PSY | EUR | |
| 198 | ATP2B15, GALNT43 | AC010201.22 | rs2681472 | Chr12:89615182 | ALC, BC, BMI, BP, PSY | EUR, JPN |
| 199 | FGD67, VEZT3 | rs11107903 | Chr12:95114195 | BC | EUR, JPN | |
| 200 | CORO1C4, SELPLG1 | rs1558802 | Chr12:108642472 | L, PSY | Multi | |
| 201 | ATXN24, SH2B32 | rs10774625, rs11066015 | Chr12:111472415-111730205 | ALC, BC, BMI, BP, I, L, PSY, SM | EUR, JPN | |
| 202 | TBX34, TBX51 | rs34606058 | Chr12:114915563 | BC, BH, BP | EUR, Multi | |
| 203 | KSR24, FBXW81 | rs11830157 | Chr12:117827636 | EUR | ||
| 204 | HNF1A6, C12orf435 | RNU4-22, HNF1A-AS12 | rs2244608 | Chr12:120979185 | ALC, BC, I, L, PSY | EUR |
| 205 | ZNF6646, CCDC9211 | rs7133378 | Chr12:123924955 | BMI, D, L, PSY, SM | EUR | |
| 206 | SCARB111, UBC2 | rs7485656, rs7296737 | Chr12:124831101-124852410 | I, L | BLK, EUR | |
| 207 | ZNF2682, ZNF1402 | AC131212.32 | rs36127550 | Chr12:133203723 | BMI, BP | Multi |
| 208 | FLT15, SLC46A32 | rs17086617, rs74412485 | Chr13:28388549-28468295 | BH, BP | EUR, JPN | |
| 209 | PDS5B8, N4BP2L25 | rs7991314 | Chr13:32551937 | BC, BH, BMI, SM | EUR | |
| 210 | DOCK93, SLC15A11 | rs8000794 | Chr13:98782556 | EUR | ||
| 211 | COL4A119, COL4A212 | rs11617955, rs3783113, rs11619113, rs4773141, rs9515203, rs7333991 | Chr13:110165755-110456089 | BP, C, L | BLK, EUR, JPN | |
| 212 | F102, MCF2L1 | rs4907571 | Chr13:112964182 | BP | EUR | |
| 213 | ARID4A2, ARMH42 | PSMA3-AS12 | rs2145598 | Chr14:58327283 | L | EUR |
| 214 | EIF2B27, MLH36 | rs10131894 | Chr14:74980176 | EUR | ||
| 215 | ITPK14, ATXN32 | rs1956028 | Chr14:93040852 | BC | Multi | |
| 216 | SERPINA14, DDX241 | rs112635299 | Chr14:94371805 | ALC, BH, BMI, BP, I, L, PSY | EUR | |
| 217 | HHIPL111, CYP46A15 | AL160313.12 | rs36033161, rs2895811 | Chr14:99657150-99667605 | BP | EUR, JPN |
| 218 | EXD12, KNSTRN2 | OIP5-AS13 | rs3959554 | Chr15:41151726 | BMI, BP | EUR |
| 219 | NEDD43, PIGB3 | AC018926.12 | rs11633313 | Chr15:55843511 | Multi | |
| 220 | LIPC5, ALDH1A25 | HSP90AB4P2 | rs588136 | Chr15:58438299 | L, PSY | EUR |
| 221 | TRIP45, RBPMS23 | rs6494488 | Chr15:64732005 | L | EUR | |
| 222 | SMAD37, SMAD63 | IQCH-AS12 | rs56062135 | Chr15:67163292 | EUR, HISP | |
| 223 | ADAMTS713, CTSH10 | AC090607.22 | rs7177201, rs7173743, rs112238647, rs7174727 | Chr15:78759363-78858944 | BH, BP,C, SM | EUR, HISP, JPN |
| 224 | CFAP1613, MESD2 | rs2683260 | Chr15:81093211 | EUR | ||
| 225 | AKAP133, KLHL253 | LOC1019296792, AC044860.12 | rs7177107 | Chr15:85580133 | BC | EUR |
| 226 | MFGE85, ABHD24 | rs1807214 | Chr15:89022026 | BP | EUR, JPN | |
| 227 | FES9, FURIN6 | rs7183988 | Chr15:90885359 | BP, L, PSY | BLK, EUR, JPN | |
| 228 | NR2F22 | rs17581137 | Chr15:95603185 | EUR | ||
| 229 | MYH115, NOMO12 | AC138969.12 | rs12691049 | Chr16:15815656 | AF, BP | EUR |
| 230 | FTO5, AKTIP1 | rs1421085 | Chr16:53767042 | ALC, BH, BMI, BP, D, I, L, PSY, SM | BLK, EUR | |
| 231 | CETP6, HERPUD13 | rs12446515 | Chr16:56953103 | AF, BMI, BP, C, L | EUR | |
| 232 | NOB13, NFAT52 | PDXDC2P3 | rs9302604 | Chr16:69542991 | BP, PSY, SM | EUR |
| 233 | DHODH7, DHX383 | rs12445401 | Chr16:72114520 | ALC, L, PSY | EUR | |
| 234 | CFDP16, TMEM170A5 | AC009054.22 | rs8046696 | Chr16:75408245 | ALC, BC, BH, BP, L, SM | EUR, JPN |
| 235 | CMIP8, PLCG26 | AC009148.12 | rs10493891, rs7189462 | Chr16:81477137-81874262 | BP, L | EUR |
| 236 | CDH134, OSGIN11 | rs7500448 | Chr16:83012185 | BP | EUR | |
| 237 | rs55880988 | Chr16:88479845 | L, I | Multi | ||
| 238 | SMG65, SRR3 | rs4790881 | Chr17:2165638 | L | EUR | |
| 239 | TOM1L27, DRC37 | CCDC144BP2, USP32P22 | rs12936927 | Chr17:17823651 | ALC, BMI, PSY, SM | EUR, HISP |
| 240 | SSH28, CORO65 | SH3GL1P22 | rs11080107 | Chr17:29611406 | BH, BMI | EUR |
| 241 | COPRS2, NF11 | rs76954792 | Chr17:31706495 | BP | EUR | |
| 242 | FBXL207, MED15 | rs1077715 | Chr17:39527295 | ALC, BC, L, SM | EUR | |
| 243 | CAVIN18, DHX586 | NBR24, AC099811.53 | rs12952244, rs8068844, rs9912587 | Chr17:42118063-43021069 | BMI, D, I, L | EUR, Multi |
| 244 | ARHGAP275, GOSR25 | LINC022102, NSFP12 | rs5026246, rs1706731, rs1881193, rs1881194, rs2532235, rs2696671, rs17608766 | Chr17:45495530-46935905 | AF, BP, PSY | EUR, HISP |
| 245 | SNF810, ATP5MC110 | SUMO2P174, LOC1053718142 | rs46522, rs4643373, rs5820757 | Chr17:48911235-49340581 | ALC, BH, BMI, BP, I, L, PSY | EUR |
| 246 | BCAS34, PPM1D2 | RPL12P382 | rs11655024 | Chr17:61155004 | BMI, BP | EUR, JPN |
| 247 | PECAM15, ERN12 | AC046185.32 | rs11079536 | Chr17:64315043 | BC, BP | EUR, JPN |
| 248 | WIPI16, BPTF3 | rs2909217 | Chr17:68467844 | L | EUR | |
| 249 | TRIM4710, UNC13D8 | ATF4P33 | rs2410859, rs78532451 | Chr17:75845204-75883011 | BP, L | EUR, Multi |
| 250 | rs112735431 | Chr17:80385145 | JPN | |||
| 251 | CTAGE13, GATA63 | rs9951447 | Chr18:22429728 | EUR, JPN | ||
| 252 | NPC14, RMC14 | rs1623003 | Chr18:23585199 | ALC, BP, L, SM | EUR | |
| 253 | rs4327120 | Chr18:38953012 | SM | Multi | ||
| 254 | LIPG5, SMAD74 | AC022919.12 | rs9945890, rs12965923 | Chr18:48989546-49687312 | L | EUR |
| 255 | LMAN16, MC4R5 | rs11663411, rs476828 | Chr18:59293278-60185354 | ALC, BH, BMI, BP, D, I, L, PSY, SM | EUR | |
| 256 | rs150651184 | Chr19:282148 | BLK | |||
| 257 | DOT1L6, PLEKHJ15 | AC006130.32 | rs191615952 | Chr19:2236098 | BH, BP | Multi |
| 258 | KDM4B5, TICAM12 | rs12185519 | Chr19:4967728 | BH, BMI, L | Multi | |
| 259 | ANGPTL46, MAP2K71 | rs116843064 | Chr19:8364439 | ALC, BMI, L | EUR | |
| 260 | LDLR18, SMARCA46 | ZNF433-AS14, AC008770.32 | rs55997232, rs10422256, rs72658867, rs34774090, rs167479 | Chr19:11077441-11416089 | ALC, BC, BP, L, PSY | BLK, EUR, HISP, JPN |
| 261 | CYP4F22, EPS15L12 | rs77420750 | Chr19:15868934 | EUR | ||
| 262 | KLF26, AP1M12 | rs10420373 | Chr19:16319866 | I | JPN | |
| 263 | MYO9B6, HAUS85 | rs7246865 | Chr19:17108295 | BC, BMI, BP | EUR | |
| 264 | MAP1S9, FCHO16 | rs10410487 | Chr19:17718799 | BC, BH, I | EUR | |
| 265 | ELL2, ISYNA12 | rs78030362 | Chr19:18464383 | BC, BMI, BP | EUR | |
| 266 | ZNF1007, ZNF4933 | rs8102320 | Chr19:21754083 | BP | Multi | |
| 267 | ZNF5078, DPY19L31 | rs12976411 | Chr19:32391114 | EUR | ||
| 268 | EIF3K2, ZFP302 | ZNF7815, LINC015352 | rs73025613 | Chr19:37843721 | Multi | |
| 269 | TGFB115, B9D26 | AC011462.53 | rs60315715, rs11466359, rs1800469 | Chr19:41284182-41354391 | BC, BP | EUR, HISP, JPN |
| 270 | APOE12, APOC18 | rs429358, rs7412, rs183657985 | Chr19:44908684-45239240 | BC, BH, BP, C, I, L | BLK, EUR, JPN | |
| 271 | RSPH6A4, DMWD4 | PPP5D1P2, GEMIN7-AS12 | rs8108474 | Chr19:45798221 | BH, BP, L | EUR |
| 272 | RRBP16, DSTN3 | rs1132274 | Chr20:17615510 | BH, L | EUR, Multi | |
| 273 | ITCH7, NCOA64 | FER1L42 | rs6088595 | Chr20:34770696 | BC, BH, BMI, D, I, L, SM | EUR |
| 274 | PROCR7, EDEM24 | NORAD2 | rs17406518 | Chr20:35211373 | BC | EUR |
| 275 | MAFB4 | rs2207132 | Chr20:40513876 | BMI, I, L | EUR | |
| 276 | PLCG16, TOP14 | rs6102343 | Chr20:41295639 | BMI, L | EUR | |
| 277 | PLTP3, CD403 | rs8124182 | Chr20:45980262 | L | EUR | |
| 278 | PREX15, ARFGEF22 | RN7SL197P2 | rs2008614 | Chr20:48816613 | BP | EUR |
| 279 | rs4809766 | Chr20:49942802 | EUR | |||
| 280 | GNAS3, ZNF8312 | APCDD1L-DT2, LINC017112 | rs6026739 | Chr20:59164414 | BH, BP | EUR |
| 281 | rs151206628 | Chr21:10592292 | HISP | |||
| 282 | NRIP13, HSPA131 | rs2064040 | Chr21:15212021 | Multi | ||
| 283 | MAP3K7CL5, BACH14 | LINC001893 | rs2832227 | Chr21:29160755 | EUR | |
| 284 | SLC5A36, MRPS63 | AP000318.22 | rs28451064, rs149487184 | Chr21:34221526-34233562 | BC, BMI, BP | EUR |
| 285 | ARVCF8, TANGO25 | DGCR112 | rs4819811, rs71313931 | Chr22:19621733-19972661 | BC, BP, I | EUR |
| 286 | SUSD27, GGT56 | POM121L9P3, GGTLC4P2 | rs12484557, rs5760309 | Chr22:24159893-24281863 | EUR | |
| 287 | OSM4, LIF3 | rs6006426 | Chr22:30273894 | EUR | ||
| 288 | FBXO71 | rs137525 | Chr22:32900153 | Multi | ||
| 289 | SCUBE13, TTLL122 | rs139012 | Chr22:43227966 | EUR | ||
| 290 | PNPLA32, PRR52 | rs738408 | Chr22:43928850 | ALC, BC, L | Multi | |
| 291 | TBL1X | rs5934659 | ChrX:9610064 | EUR | ||
| 292 | OPHN11, AR1 | rs1410127 | ChrX:68060539 | Multi | ||
| 293 | CYSLTR11, TAF9B1 | rs398484 | ChrX:78343972 | Multi | ||
| 294 | VDAC1P1 | rs2066280 | ChrX:80922131 | EUR | ||
| 295 | SETP4 | rs2342572 | ChrX:84814364 | Multi | ||
| 296 | CHRDL11, RGAG11 | rs7884019 | ChrX:110566261 | EUR | ||
| 297 | MAP7D31, FHL11 | rs5929743 | ChrX:136236818 | EUR | ||
| 298 | ARHGEF61 | RP11-308D16.21 | rs5975828 | ChrX:136904390 | EUR | |
| 299 | DNASE1L11, ATP6AP11 | rs147967693 | ChrX:154410919 | Multi |
In total, 393 independent CAD associations were collapsed into 290 autosomal and 9 X-linked genetic loci (locus ID) based on the proximal or most likely causal gene(s), including protein and RNA coding genes. The internal confidence score (cs) for causality is displayed as a superscript number [max. score = 29 for plasminogen (PLG)]. The column ‘index rsIDs' includes independent CAD associations within one gene region. Additional associations for CAD risk factors are shown in column ‘Traits’, including lipids (L), body mass index (BMI), body height (BH), blood pressure (BP), inflammation (I), calcification (C), blood coagulation (BC), diabetes (D), alcohol intake (ALC), smoking (SM), and psychological factors (e.g. sleep) (PSY). The last column shows the main ethnic groups for which CAD risk variants were identified (EUR = European, JPN = Japanese, HISP = Hispanic, BLK = black/African American, and Multi = mixed cohort).
Summary of genome-wide significant CAD risk loci
| Locus ID | Genes (protein) | Genes (RNA) | Index rsIDs | Coordinates (hg38) | Traits | Ethnicity |
|---|---|---|---|---|---|---|
| 1 | SKI4, MORN13 | rs2843152 | Chr1:2314131 | L | EUR | |
| 2 | PRDM167, ACTRT22 | rs7413494, rs2493298 | Chr1:3069321-3409348 | BP | EUR | |
| 3 | DHDDS7, RPS6KA13 | DHDDS-AS12 | rs12046497 | Chr1:26521149 | BH, L | Multi |
| 4 | ARID1A7, KDF12 | LOC1019287282 | rs79598313 | Chr1:26958422 | ALC, BH, BP, I, L | EUR |
| 5 | FHL37, INPP5B6 | rs61776719 | Chr1:37995647 | BH, BMI, BP | EUR | |
| 6 | ZMYND125, FOXO64 | rs12047439, rs2152314 | Chr1:41343968-42480791 | L, BP | Multi | |
| 7 | PCSK916, USP244 | rs34232196, rs11591147, rs151193009, rs472495 | Chr1:55023869-55055640 | ALC, BC, L, PSY | EUR, JPN | |
| 8 | PLPP310, FGGY8 | LINC013584, AC093425.12 | rs11206803, rs56170783, rs71646019, rs12733512 | Chr1:56411837-59181306 | BH, BMI, BP, SM | EUR, HISP, JPN, Multi |
| 9 | PSRC18, CELSR26 | rs12740374, rs655246 | Chr1:109274968-109289661 | ALC, BC, BH, BP, I, L, PSY | BLK, EUR, HISP | |
| 10 | PHTF13, MAGI32 | rs1230666 | Chr1:113630788 | EUR | ||
| 11 | NGF8, TSPAN22 | rs11806316, rs61797068 | Chr1:115210861-115359893 | EUR | ||
| 12 | ECM16, MTMR115 | ADAMTSL4-AS12, AL356356.12 | rs67807996, rs11585169 | Chr1:150023307-150599561 | BH, BMI, BP, I, SM | EUR |
| 13 | TDRKH9, RIIAD13 | TDRKH-AS13 | rs11810571 | Chr1:151789832 | PSY | EUR |
| 14 | IL6R5, TPM32 | rs6686750 | Chr1:154447367 | I | EUR | |
| 15 | NME79, ATP1B15 | rs61806987 | Chr1:169345595 | BH | EUR | |
| 16 | KIAA00405, TNN1 | rs4650716 | Chr1:175159575 | EUR | ||
| 17 | DENND1B4, CRB12 | rs12022672 | Chr1:197618876 | SM | Multi | |
| 18 | DDX597, CAMSAP24 | DDX59-AS12 | rs6700559 | Chr1:200676945 | EUR | |
| 19 | LMOD17, IPO94 | IPO9-AS12 | rs2820315 | Chr1:201903136 | BH, BMI, BP, L, PSY | EUR |
| 20 | DSTYK3, NUAK23 | AC119673.22 | rs12048743 | Chr1:205145745 | L | Multi |
| 21 | HHAT7, SYT142 | RNU5A-8P2 | rs60154123 | Chr1:210295654 | EUR | |
| 22 | MIA36, AIDA4 | MIA3-AS12 | rs17163363 | Chr1:222655362 | BP | EUR, HISP, JPN |
| 23 | AGT5, COG21 | rs699 | Chr1:230710048 | BP | EUR | |
| 24 | COLEC112 | RNASEH1-DT3 | rs56236159 | Chr2:3588888 | BC, BP, I, L | Multi |
| 25 | IAH13, ADAM173 | rs2715879 | Chr2:9375422 | EUR | ||
| 26 | OSR13, TTC322 | rs16986953 | Chr2:19742712 | EUR | ||
| 27 | APOB5, TDRD152 | rs13306206, rs515135 | Chr2:21019859-21063185 | BC, L, PSY | EUR, JPN | |
| 28 | ABCG810, ABCG56 | rs4245791, rs76866386 | Chr2:43847292-43848344 | BC, I, L, PSY | EUR | |
| 29 | PRKCE7, SRBD11 | rs582384 | Chr2:45669298 | BP | EUR | |
| 30 | BCL11A4, REL1 | MIR4432HG2 | rs243071 | Chr2:60391893 | EUR | |
| 31 | GFPT15, NFU13 | rs4346430 | Chr2:69441670 | Multi | ||
| 32 | GGCX7, VAMP86 | PARTICL2 | rs10176176 | Chr2:85534925 | BMI, L | EUR, JPN |
| 33 | IL1F105, PSD43 | rs6759676 | Chr2:113078771 | BP, I | EUR | |
| 34 | SAP1305, WDR331 | rs114192718 | Chr2:128028089 | EUR | ||
| 35 | ARHGAP155, KYNU1 | rs4662330 | Chr2:143428906 | BP | EUR | |
| 36 | ZEB212, GTDC13 | LINC014123, AC009951.42 | rs6740731, rs17408036, rs10928241 | Chr2:144513025-145073861 | BP, SM | EUR, HISP |
| 37 | ACVR2A4, MBD52 | rs35611688 | Chr2:147620292 | EUR | ||
| 38 | FIGN5, GRB141 | rs10930115 | Chr2:164073872 | BP, PSY, SM | EUR, JPN | |
| 39 | TFPI4, CALCRL3 | rs840616 | Chr2:187331742 | BMI, BP, L | EUR | |
| 40 | NBEAL17, CARF7 | rs148812085 | Chr2:203012510 | BMI, BP, L, PSY, SM | EUR, HISP | |
| 41 | FN17, ATIC2 | rs1250247 | Chr2:215434906 | BH, BMI, BP, L | EUR | |
| 42 | TNS15, TMBIM12 | rs2161967 | Chr2:217815806 | BP, L | EUR, HISP | |
| 43 | IRS15, NYAP22 | rs952227 | Chr2:226197364 | ALC, BMI, BP, D, L, SM | EUR | |
| 44 | PID15, DNER3 | rs4140748 | Chr2:229140789 | L | EUR, JPN | |
| 45 | GIGYF27, SNORC4 | rs283485 | Chr2:232780981 | BH, BMI | EUR | |
| 46 | COL6A34, RBM441 | rs11677932 | Chr2:237315312 | BP | EUR | |
| 47 | FGD55, NR2C21 | rs34991912 | Chr3:14884844 | BH, BP | EUR, JPN | |
| 48 | ZNF5897, CDC25A6 | AC134772.12 | rs7633770, rs7617773 | Chr3:46647072-48152025 | BH, BP | EUR |
| 49 | IP6K23, IHO13 | AC121247.12 | rs34759087 | Chr3:49124851 | BH, BMI, PSY | EUR |
| 50 | ITIH43, NISCH2 | rs77347777 | Chr3:52814191 | BMI, BP | EUR | |
| 51 | ITGB55, UMPS4 | rs17843797 | Chr3:124734175 | BMI | EUR | |
| 52 | DNAJC135, NPHP34 | rs10512861 | Chr3:132539117 | I, L | EUR | |
| 53 | PCCB7, PPP2R3A5 | NCK1-DT3 | rs34330586 | Chr3:136081567 | BMI, I, L | EUR |
| 54 | MRAS5, PIK3CB3 | rs185244 | Chr3:138374047 | BMI, BP, L | EUR | |
| 55 | ATP1B34, PLS12 | rs7622417 | Chr3:141907157 | BMI, L | Multi | |
| 56 | ARHGEF268, DHX367 | ARHGEF26-AS13 | rs357494 | Chr3:154219964 | BMI, BP, L | EUR, HISP |
| 57 | CCNL14, TIPARP4 | rs4266144 | Chr3:157134803 | BH, BMI | EUR | |
| 58 | MECOM2, SEC621 | rs16853198 | Chr3:169122391 | BP | Multi | |
| 59 | FNDC3B2, ECT21 | rs34229028 | Chr3:172399666 | EUR | ||
| 60 | rs73070813 | Chr3:187168807 | EUR | |||
| 61 | HGFAC2 | rs16844401 | Chr4:3447925 | BC, BH, L | EUR | |
| 62 | GNPDA21 | rs10938398 | Chr4:45184122 | BMI, I, L | Multi | |
| 63 | FIP1L12, LNX11 | rs17083333 | Chr4:53705899 | EUR | ||
| 64 | REST9, SPINK22 | rs781663 | Chr4:56915588 | BC, BH, I, L | EUR, JPN | |
| 65 | SHROOM35, CCDC1582 | rs12500824 | Chr4:76495474 | L, BP | EUR | |
| 66 | ANTXR22, PRDM81 | rs10857147 | Chr4:80259918 | L, BP | EUR, HISP | |
| 67 | RASGEF1B4, PRKG21 | rs36002015 | Chr4:81704567 | EUR | ||
| 68 | UNC5C2, BMPR1B2 | rs3775058 | Chr4:95196220 | EUR | ||
| 69 | MAD2L15, PDE5A4 | rs7678555 | Chr4:119988346 | BP | EUR | |
| 70 | PCDH182 | rs7439567 | Chr4:137543688 | BP | Multi | |
| 71 | ZNF8275, MMAA1 | rs13124853 | Chr4:145863622 | BP, L | EUR | |
| 72 | EDNRA14, ARHGAP103 | AC093908.12 | rs13120678, rs1878406, rs6841581 | Chr4:147352245-147480038 | BC, BP | EUR, JPN |
| 73 | GUCY1A110, MAP92 | RP11-588K22.22 | rs7440763, rs3796587 | Chr4:155512368-155716921 | ALC, BP, L | EUR, HISP, JPN |
| 74 | PALLD4, DDX60L1 | rs869396 | Chr4:168766849 | BP | EUR | |
| 75 | IRX13 | rs10041378 | Chr5:4094052 | JPN | ||
| 76 | SEMA5A7, TAS2R11 | SNHG185 | rs17263917 | Chr5:9552226 | BC | EUR |
| 77 | BASP12 | rs1032763 | Chr5:17118821 | Multi | ||
| 78 | ITGA15, PELO1 | rs4074793 | Chr5:52897294 | L, I | EUR, Multi | |
| 79 | ARL152, HSPB31 | AC027329.12 | rs7719168 | Chr5:53996560 | BP, L | EUR |
| 80 | MAP3K14, IL6ST2 | rs62362364, rs3936510 | Chr5:56145744-56565039 | BMI, BP, L, SM | EUR | |
| 81 | ANKDD1B7, HMGCR6 | rs12916 | Chr5:75360714 | BC, BMI, L, PSY | EUR | |
| 82 | LNPEP7, ERAP27 | AC009126.12 | rs27660 | Chr5:97012071 | BP, I, L, PSY | Multi |
| 83 | FER11, FBXL174 | LINC010233 | rs288173, rs112949822 | Chr5:108016593-108749489 | BH, BMI, BP, I | EUR, Multi |
| 84 | NREP1, WDR361 | rs256277 | Chr5:112071113 | Multi | ||
| 85 | DMXL12, TNFAIP81 | rs13169691 | Chr5:119112584 | EUR | ||
| 86 | SRFBP15, LOX3 | AC008568.12 | rs4345341 | Chr5:121943056 | BP | EUR |
| 87 | PRDM63, SNX241 | rs73796819 | Chr5:123084735 | Multi | ||
| 88 | FBN28, SLC27A62 | rs6883598 | Chr5:128590497 | BP | EUR | |
| 89 | IRF14, RAD504 | rs10477741, rs2706399 | Chr5:132459618-132532010 | L | EUR | |
| 90 | FGF16, SPRY42 | rs249760, rs3776307 | Chr5:142536127-143114600 | L | EUR | |
| 91 | SGCD4, MED72 | rs39929 | Chr5:156696640 | L, BP | Multi | |
| 92 | ADAM195, FNDC91 | rs11465228 | Chr5:157575687 | Multi | ||
| 93 | FOXC15, GMDS3 | rs72836800 | Chr6:1617092 | BH | EUR | |
| 94 | BLOC1S51, TXNDC51 | rs6597292 | Chr6:7975026 | Multi | ||
| 95 | PHACTR16, GFOD12 | rs9349379 | Chr6:12903725 | ALC, BC, BP, C | EUR, HISP, JPN | |
| 96 | HDGFL14, PRL1 | rs6909752 | Chr6:22612400 | AF | EUR | |
| 97 | HLA-A4, HLA-G3 | HLA-F-AS15, HLA-H3 | rs29266 | Chr6:29607502 | L | EUR |
| 98 | MICA4, PPP1R104 | rs139141104, rs9266631 | Chr6:31021244-31379121 | L | EUR, JPN | |
| 99 | CLIC13, DDAH23 | SNHG322, LOC1005075472 | rs3130683 | Chr6:31920590 | BH, BMI, BP, L, PSY | EUR |
| 100 | LEMD23, UQCC23 | rs62397561 | Chr6:33816328 | Multi | ||
| 101 | BLTP3A9, SNRPC8 | ILRUN-AS12 | rs9469899, rs17609940 | Chr6:34825347-35067023 | BH, BMI, L, PSY | EUR |
| 102 | CDKN1A5, ETV72 | DINOL2 | rs1321309 | Chr6:36670859 | BP, I, L | EUR |
| 103 | KCNK510, SAYSD11 | rs733701, rs10947789 | Chr6:39204086-39207146 | BP | EUR | |
| 104 | PEX63, PTK71 | rs1034246 | Chr6:43100632 | EUR | ||
| 105 | VEGFA4, MRPS18A1 | rs6905288, rs149372871 | Chr6:43791136-44080314 | ALC, BMI, BP, L, PSY | EUR, HISP, JPN | |
| 106 | RAB2310, PRIM26 | LOC1005061884, RP11-203B9.42 | rs11752218, rs9367716 | Chr6:57280764-57295774 | EUR | |
| 107 | CD1096 | CD109-AS12 | rs79717953, rs56171536 | Chr6:73695048-73706145 | JPN, Multi | |
| 108 | TENT5A4, IBTK1 | LINC0254`22 | rs9361867 | Chr6:81886242 | EUR | |
| 109 | UFL16, FHL55 | rs2983896 | Chr6:96581995 | BP | EUR | |
| 110 | CENPW8, TRMT112 | rs35510806 | Chr6:126357186 | BH, I, L, PSY | EUR | |
| 111 | TCF2113, SLC2A127 | TARID3 | rs6919211, rs2327426, rs2492304 | Chr6:133678730-134057013 | BH, BP | EUR, HISP, JPN |
| 112 | PLEKHG14, MTHFD1L2 | RAET1K2 | rs17080093 | Chr6:150676304 | BP | EUR |
| 113 | PLG29, LPA19 | LPAL22 | rs6932293, rs186696265, rs56195640, rs192425087, rs28867400, rs932631509, rs10455872, rs73596816, rs79018195, rs1998043, rs4252120, rs184278183 | Chr6:160114846-160756724 | BC, BMI, BP, C, L, PSY | BLK, EUR, HISP, JPN |
| 114 | MAD1L16, ELFN12 | rs62435159 | Chr7:1871645 | BMI | EUR | |
| 115 | DAGLB8, RAC17 | rs10951983 | Chr7:6406396 | I, L | EUR | |
| 116 | TMEM106B7, THSD7A2 | rs11509880 | Chr7:12222285 | L, PSY | EUR | |
| 117 | ETV14 | rs2073533 | Chr7:13990114 | EUR | ||
| 118 | TWIST15, HDAC92 | rs2107595 | Chr7:19009765 | BP | EUR, JPN | |
| 119 | MACC12, ITGB82 | rs10486389 | Chr7:20260793 | EUR | ||
| 120 | TBX208, DPY19L12 | DPY19L2P12 | rs2215614 | Chr7:35237481 | BH | EUR |
| 121 | CCM25, TBRG41 | rs2107732 | Chr7:45038379 | BP | EUR | |
| 122 | SEMA3C5 | rs917191 | Chr7:80941555 | BMI | Multi | |
| 123 | ZKSCAN16, AP4M15 | STAG3L5P2 | rs35146811 | Chr7:100123371 | BH, BP, L | EUR |
| 124 | COG59, BCAP298 | WBP1LP23, RPL7P322 | rs12112877, rs10953541 | Chr7:107300879-107604100 | L, PSY, SM | EUR |
| 125 | CFTR3, ASZ12 | rs13222797 | Chr7:117459992 | BMI, L | EUR | |
| 126 | ZC3HC15, KLHDC102 | rs11556924 | Chr7:130023656 | BC, BP, I, L, PSY, SM | EUR | |
| 127 | PARP122, HIPK21 | rs5887949 | Chr7:140060741 | BH | EUR | |
| 128 | NOS35, KCNH21 | rs3918226 | Chr7:150993088 | ALC, BC, BP, L, SM | EUR | |
| 129 | MSR13, CNOT71 | rs1480146 | Chr8:16272064 | Multi | ||
| 130 | NAT23, PCM13 | rs4646249 | Chr8:18402921 | L | EUR | |
| 131 | LPL14, CSGALNACT13 | rs894211, rs66778572, rs268 | Chr8:19956018-20058609 | ALC, BC, BH, BMI, BP, I, L, PSY, SM | BLK, EUR, HISP | |
| 132 | BMP15, PHYHIP2 | rs56408342 | Chr8:22190977 | EUR | ||
| 133 | DOCK56, KCTD92 | rs1510758 | Chr8:25204291 | EUR | ||
| 134 | PPP2R2A2 | rs6557894 | Chr8:26229397 | BH | EUR | |
| 135 | RP15 | rs9298506 | Chr8:54524964 | ALC, BH, I, L | Multi | |
| 136 | C8orf342 | rs2380472 | Chr8:68519476 | JPN | ||
| 137 | GEM6, CDH172 | rs34917849 | Chr8:94266079 | BP | EUR | |
| 138 | NDUFAF65, TP53INP11 | rs3019173 | Chr8:95008746 | Multi | ||
| 139 | rs10093110 | Chr8:105553186 | EUR | |||
| 140 | TRIB14, NSMCE21 | rs2001846 | Chr8:125466208 | ALC, BMI, BP, D, I, L | EUR, HISP | |
| 141 | NFIB2, MPDZ1 | LURAP1L-AS12 | rs10961206 | Chr9:13724052 | EUR | |
| 142 | ACER27, RPS61 | AL391834.12 | rs10811183 | Chr9:19436057 | L | EUR |
| 143 | CDKN2A13, CDKN2B11 | ERVFRD-36, MIR31HG4 | rs11523031, rs2891168, rs6475608, rs76959412 | Chr9:21843843-22101703 | BC, BP, C, L | BLK, EUR, HISP, JPN |
| 144 | WNK23, CARD192 | rs4617217 | Chr9:93226686 | Multi | ||
| 145 | ABCA12, NIPSNAP3B2 | rs35093463 | Chr9:104823957 | L | JPN | |
| 146 | KLF44, RAD23B1 | rs1967604 | Chr9:107768043 | BC, L | EUR | |
| 147 | SVEP14 | rs111245230 | Chr9:110407495 | BC, BP | EUR | |
| 148 | ASTN22 | rs7873551 | Chr9:116482848 | BH, PSY | Multi | |
| 149 | PHF195, C54 | rs41312891 | Chr9:120964472 | BH | EUR, Multi | |
| 150 | DAB2IP5, STOM1 | CUTALP2 | rs885150 | Chr9:121657894 | BC, BH, BMI, L | EUR |
| 151 | ABO6, CEL3 | rs651007 | Chr9:133278431 | BC, BP, D, I, L, PSY | BLK, EUR, HISP | |
| 152 | CAMK1D4, CDC1232 | rs17566555 | Chr10:12233948 | PSY | EUR | |
| 153 | ARHGAP214, GPR1581 | rs7077962 | Chr10:24765745 | Multi | ||
| 154 | JCAD7, SVIL1 | rs9337951 | Chr10:30028144 | BP, SM | EUR, HISP | |
| 155 | ZEB16, ARHGAP121 | ZEB1-AS15, PPIAP312 | rs161231 | Chr10:31466991 | L | Multi |
| 156 | CXCL1212, RASGEF1A2 | FRG1JP2 | rs1870634, rs494207, rs527052 | Chr10:43985363-44245808 | C | BLK, EUR, HISP |
| 157 | BICC17, TFAM1 | rs10740732 | Chr10:58593128 | BP | Multi | |
| 158 | TSPAN147, SH2D4B2 | LOC1019295742 | rs17680741 | Chr10:80491758 | EUR | |
| 159 | LIPA6, IFIT11 | rs1051338 | Chr10:89247603 | I, L | EUR, JPN | |
| 160 | PLCE112, SLC35G12 | rs1223583, rs55753709 | Chr10:93996743-94269413 | BMI, BP, L, PSY | EUR | |
| 161 | LOXL49, R3HCC1L3 | rs884811 | Chr10:98164006 | BH, BP | EUR | |
| 162 | CNNM28, BORCS75 | rs77787671 | Chr10:103016448 | BMI, BP, PSY | EUR, HISP, JPN | |
| 163 | STN13, SH3PXD2A2 | rs2067831 | Chr10:103883465 | BP, SM | EUR | |
| 164 | TCF7L25 | rs7903146 | Chr10:112998590 | BH, BMI, BP, D, L, SM | Multi | |
| 165 | C10orf823, GFRA11 | rs2218584 | Chr10:116649131 | L | Multi | |
| 166 | WDR111 | rs17101521 | Chr10:121156039 | BMI, BP, L | Multi | |
| 167 | HTRA16, TACC22 | rs2672592, rs4752700, rs17103659 | Chr10:122471234-122515087 | EUR, HISP | ||
| 168 | BET1L3, PSMD132 | rs73386640 | Chr11:203235 | JPN | ||
| 169 | TRIM54, TRIM221 | rs11601507 | Chr11:5679844 | BC, L | EUR | |
| 170 | DENND2B2, TRIM662 | LOC1027247842, TMEM9B-AS12 | rs56210063 | Chr11:8767618 | Multi | |
| 171 | SWAP708, TMEM41B5 | LOC4400282, SBF2-AS12 | rs4537761, rs360153 | Chr11:9301806-9740727 | BC, BP, D | EUR |
| 172 | CTR92 | MRVl12 | rs11042937 | Chr11:10723847 | EUR | |
| 173 | BMAL16, ARNTL2 | rs11316597 | Chr11:13274205 | BP, PSY | EUR | |
| 174 | BDNF2, LIN7C2 | LINC006782 | rs6265 | Chr11:27658369 | ALC, BMI, BP, I, L, PSY, SM | Multi |
| 175 | WT14, IMMP1L2 | rs7118294 | Chr11:32358975 | BH, BP, L | EUR | |
| 176 | HSD17B127, TTC171 | rs7116641 | Chr11:43675367 | BH, BMI, I | EUR | |
| 177 | SIPA18, RNASEH2C3 | rs2306363 | Chr11:65638129 | ALC, BMI, BP, L | EUR, JPN | |
| 178 | SERPINH117, MAP64 | AP001922.52 | rs590121, rs584961 | Chr11:75563105-75566583 | BH, BMI | EUR |
| 179 | ARHGAP4211, PGR1 | LOC1001283862 | rs633185, rs7947761 | Chr11:100722807-100753868 | ALC, BP | EUR |
| 180 | MMP132, MMP31 | rs1892971 | Chr11:102924877 | EUR | ||
| 181 | PDGFD5, DYNC2H11 | rs2839812 | Chr11:103802566 | BMI, BP | EUR, JPN | |
| 182 | CWF19L25, GUCY1A22 | rs7117780 | Chr11:107215397 | BP | EUR | |
| 183 | RDX5, FDX14 | rs10488763 | Chr11:110373636 | EUR, JPN | ||
| 184 | SIK27, CRYAB5 | rs11213945 | Chr11:111590279 | BP | Multi | |
| 185 | ZPR13, BUD132 | rs964184 | Chr11:116778201 | L | EUR, HISP | |
| 186 | VPS111, MCAM1 | rs1177562 | Chr11:119078621 | L | EUR | |
| 187 | TLCD53, ARHGEF122 | rs4938809 | Chr11:120493228 | EUR | ||
| 188 | UBASH3B6, JHY1 | rs57812455 | Chr11:122654259 | BC, BP, L | EUR | |
| 189 | ST3GAL45, TIRAP2 | GSEC3 | rs10790800 | Chr11:126392743 | L | EUR |
| 190 | C1S4, CHD42 | rs72447384 | Chr12:7071137 | I | EUR | |
| 191 | YBX36, OLR11 | rs2607903 | Chr12:10723974 | JPN | ||
| 192 | PDE3A4, AEBP22 | rs10841443 | Chr12:20067099 | BP | EUR | |
| 193 | ZCRB17, PPHLN17 | rs12820154 | Chr12:42389305 | BP | Multi | |
| 194 | ZNF6414, CCDC1842 | rs1859440 | Chr12:48033546 | BC, BP | EUR | |
| 195 | SMUG13, HOXC43 | FLJ128252, RP11-834C11.42 | rs1133773, rs75160195 | Chr12:54102435-54127810 | EUR, JPN | |
| 196 | COQ10A6, STAT24 | rs143405823 | Chr12:56329935 | BH, L, PSY | Multi | |
| 197 | LRP17, STAT65 | rs11172113 | Chr12:57133500 | BH, BMI, BP, L, PSY | EUR | |
| 198 | ATP2B15, GALNT43 | AC010201.22 | rs2681472 | Chr12:89615182 | ALC, BC, BMI, BP, PSY | EUR, JPN |
| 199 | FGD67, VEZT3 | rs11107903 | Chr12:95114195 | BC | EUR, JPN | |
| 200 | CORO1C4, SELPLG1 | rs1558802 | Chr12:108642472 | L, PSY | Multi | |
| 201 | ATXN24, SH2B32 | rs10774625, rs11066015 | Chr12:111472415-111730205 | ALC, BC, BMI, BP, I, L, PSY, SM | EUR, JPN | |
| 202 | TBX34, TBX51 | rs34606058 | Chr12:114915563 | BC, BH, BP | EUR, Multi | |
| 203 | KSR24, FBXW81 | rs11830157 | Chr12:117827636 | EUR | ||
| 204 | HNF1A6, C12orf435 | RNU4-22, HNF1A-AS12 | rs2244608 | Chr12:120979185 | ALC, BC, I, L, PSY | EUR |
| 205 | ZNF6646, CCDC9211 | rs7133378 | Chr12:123924955 | BMI, D, L, PSY, SM | EUR | |
| 206 | SCARB111, UBC2 | rs7485656, rs7296737 | Chr12:124831101-124852410 | I, L | BLK, EUR | |
| 207 | ZNF2682, ZNF1402 | AC131212.32 | rs36127550 | Chr12:133203723 | BMI, BP | Multi |
| 208 | FLT15, SLC46A32 | rs17086617, rs74412485 | Chr13:28388549-28468295 | BH, BP | EUR, JPN | |
| 209 | PDS5B8, N4BP2L25 | rs7991314 | Chr13:32551937 | BC, BH, BMI, SM | EUR | |
| 210 | DOCK93, SLC15A11 | rs8000794 | Chr13:98782556 | EUR | ||
| 211 | COL4A119, COL4A212 | rs11617955, rs3783113, rs11619113, rs4773141, rs9515203, rs7333991 | Chr13:110165755-110456089 | BP, C, L | BLK, EUR, JPN | |
| 212 | F102, MCF2L1 | rs4907571 | Chr13:112964182 | BP | EUR | |
| 213 | ARID4A2, ARMH42 | PSMA3-AS12 | rs2145598 | Chr14:58327283 | L | EUR |
| 214 | EIF2B27, MLH36 | rs10131894 | Chr14:74980176 | EUR | ||
| 215 | ITPK14, ATXN32 | rs1956028 | Chr14:93040852 | BC | Multi | |
| 216 | SERPINA14, DDX241 | rs112635299 | Chr14:94371805 | ALC, BH, BMI, BP, I, L, PSY | EUR | |
| 217 | HHIPL111, CYP46A15 | AL160313.12 | rs36033161, rs2895811 | Chr14:99657150-99667605 | BP | EUR, JPN |
| 218 | EXD12, KNSTRN2 | OIP5-AS13 | rs3959554 | Chr15:41151726 | BMI, BP | EUR |
| 219 | NEDD43, PIGB3 | AC018926.12 | rs11633313 | Chr15:55843511 | Multi | |
| 220 | LIPC5, ALDH1A25 | HSP90AB4P2 | rs588136 | Chr15:58438299 | L, PSY | EUR |
| 221 | TRIP45, RBPMS23 | rs6494488 | Chr15:64732005 | L | EUR | |
| 222 | SMAD37, SMAD63 | IQCH-AS12 | rs56062135 | Chr15:67163292 | EUR, HISP | |
| 223 | ADAMTS713, CTSH10 | AC090607.22 | rs7177201, rs7173743, rs112238647, rs7174727 | Chr15:78759363-78858944 | BH, BP,C, SM | EUR, HISP, JPN |
| 224 | CFAP1613, MESD2 | rs2683260 | Chr15:81093211 | EUR | ||
| 225 | AKAP133, KLHL253 | LOC1019296792, AC044860.12 | rs7177107 | Chr15:85580133 | BC | EUR |
| 226 | MFGE85, ABHD24 | rs1807214 | Chr15:89022026 | BP | EUR, JPN | |
| 227 | FES9, FURIN6 | rs7183988 | Chr15:90885359 | BP, L, PSY | BLK, EUR, JPN | |
| 228 | NR2F22 | rs17581137 | Chr15:95603185 | EUR | ||
| 229 | MYH115, NOMO12 | AC138969.12 | rs12691049 | Chr16:15815656 | AF, BP | EUR |
| 230 | FTO5, AKTIP1 | rs1421085 | Chr16:53767042 | ALC, BH, BMI, BP, D, I, L, PSY, SM | BLK, EUR | |
| 231 | CETP6, HERPUD13 | rs12446515 | Chr16:56953103 | AF, BMI, BP, C, L | EUR | |
| 232 | NOB13, NFAT52 | PDXDC2P3 | rs9302604 | Chr16:69542991 | BP, PSY, SM | EUR |
| 233 | DHODH7, DHX383 | rs12445401 | Chr16:72114520 | ALC, L, PSY | EUR | |
| 234 | CFDP16, TMEM170A5 | AC009054.22 | rs8046696 | Chr16:75408245 | ALC, BC, BH, BP, L, SM | EUR, JPN |
| 235 | CMIP8, PLCG26 | AC009148.12 | rs10493891, rs7189462 | Chr16:81477137-81874262 | BP, L | EUR |
| 236 | CDH134, OSGIN11 | rs7500448 | Chr16:83012185 | BP | EUR | |
| 237 | rs55880988 | Chr16:88479845 | L, I | Multi | ||
| 238 | SMG65, SRR3 | rs4790881 | Chr17:2165638 | L | EUR | |
| 239 | TOM1L27, DRC37 | CCDC144BP2, USP32P22 | rs12936927 | Chr17:17823651 | ALC, BMI, PSY, SM | EUR, HISP |
| 240 | SSH28, CORO65 | SH3GL1P22 | rs11080107 | Chr17:29611406 | BH, BMI | EUR |
| 241 | COPRS2, NF11 | rs76954792 | Chr17:31706495 | BP | EUR | |
| 242 | FBXL207, MED15 | rs1077715 | Chr17:39527295 | ALC, BC, L, SM | EUR | |
| 243 | CAVIN18, DHX586 | NBR24, AC099811.53 | rs12952244, rs8068844, rs9912587 | Chr17:42118063-43021069 | BMI, D, I, L | EUR, Multi |
| 244 | ARHGAP275, GOSR25 | LINC022102, NSFP12 | rs5026246, rs1706731, rs1881193, rs1881194, rs2532235, rs2696671, rs17608766 | Chr17:45495530-46935905 | AF, BP, PSY | EUR, HISP |
| 245 | SNF810, ATP5MC110 | SUMO2P174, LOC1053718142 | rs46522, rs4643373, rs5820757 | Chr17:48911235-49340581 | ALC, BH, BMI, BP, I, L, PSY | EUR |
| 246 | BCAS34, PPM1D2 | RPL12P382 | rs11655024 | Chr17:61155004 | BMI, BP | EUR, JPN |
| 247 | PECAM15, ERN12 | AC046185.32 | rs11079536 | Chr17:64315043 | BC, BP | EUR, JPN |
| 248 | WIPI16, BPTF3 | rs2909217 | Chr17:68467844 | L | EUR | |
| 249 | TRIM4710, UNC13D8 | ATF4P33 | rs2410859, rs78532451 | Chr17:75845204-75883011 | BP, L | EUR, Multi |
| 250 | rs112735431 | Chr17:80385145 | JPN | |||
| 251 | CTAGE13, GATA63 | rs9951447 | Chr18:22429728 | EUR, JPN | ||
| 252 | NPC14, RMC14 | rs1623003 | Chr18:23585199 | ALC, BP, L, SM | EUR | |
| 253 | rs4327120 | Chr18:38953012 | SM | Multi | ||
| 254 | LIPG5, SMAD74 | AC022919.12 | rs9945890, rs12965923 | Chr18:48989546-49687312 | L | EUR |
| 255 | LMAN16, MC4R5 | rs11663411, rs476828 | Chr18:59293278-60185354 | ALC, BH, BMI, BP, D, I, L, PSY, SM | EUR | |
| 256 | rs150651184 | Chr19:282148 | BLK | |||
| 257 | DOT1L6, PLEKHJ15 | AC006130.32 | rs191615952 | Chr19:2236098 | BH, BP | Multi |
| 258 | KDM4B5, TICAM12 | rs12185519 | Chr19:4967728 | BH, BMI, L | Multi | |
| 259 | ANGPTL46, MAP2K71 | rs116843064 | Chr19:8364439 | ALC, BMI, L | EUR | |
| 260 | LDLR18, SMARCA46 | ZNF433-AS14, AC008770.32 | rs55997232, rs10422256, rs72658867, rs34774090, rs167479 | Chr19:11077441-11416089 | ALC, BC, BP, L, PSY | BLK, EUR, HISP, JPN |
| 261 | CYP4F22, EPS15L12 | rs77420750 | Chr19:15868934 | EUR | ||
| 262 | KLF26, AP1M12 | rs10420373 | Chr19:16319866 | I | JPN | |
| 263 | MYO9B6, HAUS85 | rs7246865 | Chr19:17108295 | BC, BMI, BP | EUR | |
| 264 | MAP1S9, FCHO16 | rs10410487 | Chr19:17718799 | BC, BH, I | EUR | |
| 265 | ELL2, ISYNA12 | rs78030362 | Chr19:18464383 | BC, BMI, BP | EUR | |
| 266 | ZNF1007, ZNF4933 | rs8102320 | Chr19:21754083 | BP | Multi | |
| 267 | ZNF5078, DPY19L31 | rs12976411 | Chr19:32391114 | EUR | ||
| 268 | EIF3K2, ZFP302 | ZNF7815, LINC015352 | rs73025613 | Chr19:37843721 | Multi | |
| 269 | TGFB115, B9D26 | AC011462.53 | rs60315715, rs11466359, rs1800469 | Chr19:41284182-41354391 | BC, BP | EUR, HISP, JPN |
| 270 | APOE12, APOC18 | rs429358, rs7412, rs183657985 | Chr19:44908684-45239240 | BC, BH, BP, C, I, L | BLK, EUR, JPN | |
| 271 | RSPH6A4, DMWD4 | PPP5D1P2, GEMIN7-AS12 | rs8108474 | Chr19:45798221 | BH, BP, L | EUR |
| 272 | RRBP16, DSTN3 | rs1132274 | Chr20:17615510 | BH, L | EUR, Multi | |
| 273 | ITCH7, NCOA64 | FER1L42 | rs6088595 | Chr20:34770696 | BC, BH, BMI, D, I, L, SM | EUR |
| 274 | PROCR7, EDEM24 | NORAD2 | rs17406518 | Chr20:35211373 | BC | EUR |
| 275 | MAFB4 | rs2207132 | Chr20:40513876 | BMI, I, L | EUR | |
| 276 | PLCG16, TOP14 | rs6102343 | Chr20:41295639 | BMI, L | EUR | |
| 277 | PLTP3, CD403 | rs8124182 | Chr20:45980262 | L | EUR | |
| 278 | PREX15, ARFGEF22 | RN7SL197P2 | rs2008614 | Chr20:48816613 | BP | EUR |
| 279 | rs4809766 | Chr20:49942802 | EUR | |||
| 280 | GNAS3, ZNF8312 | APCDD1L-DT2, LINC017112 | rs6026739 | Chr20:59164414 | BH, BP | EUR |
| 281 | rs151206628 | Chr21:10592292 | HISP | |||
| 282 | NRIP13, HSPA131 | rs2064040 | Chr21:15212021 | Multi | ||
| 283 | MAP3K7CL5, BACH14 | LINC001893 | rs2832227 | Chr21:29160755 | EUR | |
| 284 | SLC5A36, MRPS63 | AP000318.22 | rs28451064, rs149487184 | Chr21:34221526-34233562 | BC, BMI, BP | EUR |
| 285 | ARVCF8, TANGO25 | DGCR112 | rs4819811, rs71313931 | Chr22:19621733-19972661 | BC, BP, I | EUR |
| 286 | SUSD27, GGT56 | POM121L9P3, GGTLC4P2 | rs12484557, rs5760309 | Chr22:24159893-24281863 | EUR | |
| 287 | OSM4, LIF3 | rs6006426 | Chr22:30273894 | EUR | ||
| 288 | FBXO71 | rs137525 | Chr22:32900153 | Multi | ||
| 289 | SCUBE13, TTLL122 | rs139012 | Chr22:43227966 | EUR | ||
| 290 | PNPLA32, PRR52 | rs738408 | Chr22:43928850 | ALC, BC, L | Multi | |
| 291 | TBL1X | rs5934659 | ChrX:9610064 | EUR | ||
| 292 | OPHN11, AR1 | rs1410127 | ChrX:68060539 | Multi | ||
| 293 | CYSLTR11, TAF9B1 | rs398484 | ChrX:78343972 | Multi | ||
| 294 | VDAC1P1 | rs2066280 | ChrX:80922131 | EUR | ||
| 295 | SETP4 | rs2342572 | ChrX:84814364 | Multi | ||
| 296 | CHRDL11, RGAG11 | rs7884019 | ChrX:110566261 | EUR | ||
| 297 | MAP7D31, FHL11 | rs5929743 | ChrX:136236818 | EUR | ||
| 298 | ARHGEF61 | RP11-308D16.21 | rs5975828 | ChrX:136904390 | EUR | |
| 299 | DNASE1L11, ATP6AP11 | rs147967693 | ChrX:154410919 | Multi |
| Locus ID | Genes (protein) | Genes (RNA) | Index rsIDs | Coordinates (hg38) | Traits | Ethnicity |
|---|---|---|---|---|---|---|
| 1 | SKI4, MORN13 | rs2843152 | Chr1:2314131 | L | EUR | |
| 2 | PRDM167, ACTRT22 | rs7413494, rs2493298 | Chr1:3069321-3409348 | BP | EUR | |
| 3 | DHDDS7, RPS6KA13 | DHDDS-AS12 | rs12046497 | Chr1:26521149 | BH, L | Multi |
| 4 | ARID1A7, KDF12 | LOC1019287282 | rs79598313 | Chr1:26958422 | ALC, BH, BP, I, L | EUR |
| 5 | FHL37, INPP5B6 | rs61776719 | Chr1:37995647 | BH, BMI, BP | EUR | |
| 6 | ZMYND125, FOXO64 | rs12047439, rs2152314 | Chr1:41343968-42480791 | L, BP | Multi | |
| 7 | PCSK916, USP244 | rs34232196, rs11591147, rs151193009, rs472495 | Chr1:55023869-55055640 | ALC, BC, L, PSY | EUR, JPN | |
| 8 | PLPP310, FGGY8 | LINC013584, AC093425.12 | rs11206803, rs56170783, rs71646019, rs12733512 | Chr1:56411837-59181306 | BH, BMI, BP, SM | EUR, HISP, JPN, Multi |
| 9 | PSRC18, CELSR26 | rs12740374, rs655246 | Chr1:109274968-109289661 | ALC, BC, BH, BP, I, L, PSY | BLK, EUR, HISP | |
| 10 | PHTF13, MAGI32 | rs1230666 | Chr1:113630788 | EUR | ||
| 11 | NGF8, TSPAN22 | rs11806316, rs61797068 | Chr1:115210861-115359893 | EUR | ||
| 12 | ECM16, MTMR115 | ADAMTSL4-AS12, AL356356.12 | rs67807996, rs11585169 | Chr1:150023307-150599561 | BH, BMI, BP, I, SM | EUR |
| 13 | TDRKH9, RIIAD13 | TDRKH-AS13 | rs11810571 | Chr1:151789832 | PSY | EUR |
| 14 | IL6R5, TPM32 | rs6686750 | Chr1:154447367 | I | EUR | |
| 15 | NME79, ATP1B15 | rs61806987 | Chr1:169345595 | BH | EUR | |
| 16 | KIAA00405, TNN1 | rs4650716 | Chr1:175159575 | EUR | ||
| 17 | DENND1B4, CRB12 | rs12022672 | Chr1:197618876 | SM | Multi | |
| 18 | DDX597, CAMSAP24 | DDX59-AS12 | rs6700559 | Chr1:200676945 | EUR | |
| 19 | LMOD17, IPO94 | IPO9-AS12 | rs2820315 | Chr1:201903136 | BH, BMI, BP, L, PSY | EUR |
| 20 | DSTYK3, NUAK23 | AC119673.22 | rs12048743 | Chr1:205145745 | L | Multi |
| 21 | HHAT7, SYT142 | RNU5A-8P2 | rs60154123 | Chr1:210295654 | EUR | |
| 22 | MIA36, AIDA4 | MIA3-AS12 | rs17163363 | Chr1:222655362 | BP | EUR, HISP, JPN |
| 23 | AGT5, COG21 | rs699 | Chr1:230710048 | BP | EUR | |
| 24 | COLEC112 | RNASEH1-DT3 | rs56236159 | Chr2:3588888 | BC, BP, I, L | Multi |
| 25 | IAH13, ADAM173 | rs2715879 | Chr2:9375422 | EUR | ||
| 26 | OSR13, TTC322 | rs16986953 | Chr2:19742712 | EUR | ||
| 27 | APOB5, TDRD152 | rs13306206, rs515135 | Chr2:21019859-21063185 | BC, L, PSY | EUR, JPN | |
| 28 | ABCG810, ABCG56 | rs4245791, rs76866386 | Chr2:43847292-43848344 | BC, I, L, PSY | EUR | |
| 29 | PRKCE7, SRBD11 | rs582384 | Chr2:45669298 | BP | EUR | |
| 30 | BCL11A4, REL1 | MIR4432HG2 | rs243071 | Chr2:60391893 | EUR | |
| 31 | GFPT15, NFU13 | rs4346430 | Chr2:69441670 | Multi | ||
| 32 | GGCX7, VAMP86 | PARTICL2 | rs10176176 | Chr2:85534925 | BMI, L | EUR, JPN |
| 33 | IL1F105, PSD43 | rs6759676 | Chr2:113078771 | BP, I | EUR | |
| 34 | SAP1305, WDR331 | rs114192718 | Chr2:128028089 | EUR | ||
| 35 | ARHGAP155, KYNU1 | rs4662330 | Chr2:143428906 | BP | EUR | |
| 36 | ZEB212, GTDC13 | LINC014123, AC009951.42 | rs6740731, rs17408036, rs10928241 | Chr2:144513025-145073861 | BP, SM | EUR, HISP |
| 37 | ACVR2A4, MBD52 | rs35611688 | Chr2:147620292 | EUR | ||
| 38 | FIGN5, GRB141 | rs10930115 | Chr2:164073872 | BP, PSY, SM | EUR, JPN | |
| 39 | TFPI4, CALCRL3 | rs840616 | Chr2:187331742 | BMI, BP, L | EUR | |
| 40 | NBEAL17, CARF7 | rs148812085 | Chr2:203012510 | BMI, BP, L, PSY, SM | EUR, HISP | |
| 41 | FN17, ATIC2 | rs1250247 | Chr2:215434906 | BH, BMI, BP, L | EUR | |
| 42 | TNS15, TMBIM12 | rs2161967 | Chr2:217815806 | BP, L | EUR, HISP | |
| 43 | IRS15, NYAP22 | rs952227 | Chr2:226197364 | ALC, BMI, BP, D, L, SM | EUR | |
| 44 | PID15, DNER3 | rs4140748 | Chr2:229140789 | L | EUR, JPN | |
| 45 | GIGYF27, SNORC4 | rs283485 | Chr2:232780981 | BH, BMI | EUR | |
| 46 | COL6A34, RBM441 | rs11677932 | Chr2:237315312 | BP | EUR | |
| 47 | FGD55, NR2C21 | rs34991912 | Chr3:14884844 | BH, BP | EUR, JPN | |
| 48 | ZNF5897, CDC25A6 | AC134772.12 | rs7633770, rs7617773 | Chr3:46647072-48152025 | BH, BP | EUR |
| 49 | IP6K23, IHO13 | AC121247.12 | rs34759087 | Chr3:49124851 | BH, BMI, PSY | EUR |
| 50 | ITIH43, NISCH2 | rs77347777 | Chr3:52814191 | BMI, BP | EUR | |
| 51 | ITGB55, UMPS4 | rs17843797 | Chr3:124734175 | BMI | EUR | |
| 52 | DNAJC135, NPHP34 | rs10512861 | Chr3:132539117 | I, L | EUR | |
| 53 | PCCB7, PPP2R3A5 | NCK1-DT3 | rs34330586 | Chr3:136081567 | BMI, I, L | EUR |
| 54 | MRAS5, PIK3CB3 | rs185244 | Chr3:138374047 | BMI, BP, L | EUR | |
| 55 | ATP1B34, PLS12 | rs7622417 | Chr3:141907157 | BMI, L | Multi | |
| 56 | ARHGEF268, DHX367 | ARHGEF26-AS13 | rs357494 | Chr3:154219964 | BMI, BP, L | EUR, HISP |
| 57 | CCNL14, TIPARP4 | rs4266144 | Chr3:157134803 | BH, BMI | EUR | |
| 58 | MECOM2, SEC621 | rs16853198 | Chr3:169122391 | BP | Multi | |
| 59 | FNDC3B2, ECT21 | rs34229028 | Chr3:172399666 | EUR | ||
| 60 | rs73070813 | Chr3:187168807 | EUR | |||
| 61 | HGFAC2 | rs16844401 | Chr4:3447925 | BC, BH, L | EUR | |
| 62 | GNPDA21 | rs10938398 | Chr4:45184122 | BMI, I, L | Multi | |
| 63 | FIP1L12, LNX11 | rs17083333 | Chr4:53705899 | EUR | ||
| 64 | REST9, SPINK22 | rs781663 | Chr4:56915588 | BC, BH, I, L | EUR, JPN | |
| 65 | SHROOM35, CCDC1582 | rs12500824 | Chr4:76495474 | L, BP | EUR | |
| 66 | ANTXR22, PRDM81 | rs10857147 | Chr4:80259918 | L, BP | EUR, HISP | |
| 67 | RASGEF1B4, PRKG21 | rs36002015 | Chr4:81704567 | EUR | ||
| 68 | UNC5C2, BMPR1B2 | rs3775058 | Chr4:95196220 | EUR | ||
| 69 | MAD2L15, PDE5A4 | rs7678555 | Chr4:119988346 | BP | EUR | |
| 70 | PCDH182 | rs7439567 | Chr4:137543688 | BP | Multi | |
| 71 | ZNF8275, MMAA1 | rs13124853 | Chr4:145863622 | BP, L | EUR | |
| 72 | EDNRA14, ARHGAP103 | AC093908.12 | rs13120678, rs1878406, rs6841581 | Chr4:147352245-147480038 | BC, BP | EUR, JPN |
| 73 | GUCY1A110, MAP92 | RP11-588K22.22 | rs7440763, rs3796587 | Chr4:155512368-155716921 | ALC, BP, L | EUR, HISP, JPN |
| 74 | PALLD4, DDX60L1 | rs869396 | Chr4:168766849 | BP | EUR | |
| 75 | IRX13 | rs10041378 | Chr5:4094052 | JPN | ||
| 76 | SEMA5A7, TAS2R11 | SNHG185 | rs17263917 | Chr5:9552226 | BC | EUR |
| 77 | BASP12 | rs1032763 | Chr5:17118821 | Multi | ||
| 78 | ITGA15, PELO1 | rs4074793 | Chr5:52897294 | L, I | EUR, Multi | |
| 79 | ARL152, HSPB31 | AC027329.12 | rs7719168 | Chr5:53996560 | BP, L | EUR |
| 80 | MAP3K14, IL6ST2 | rs62362364, rs3936510 | Chr5:56145744-56565039 | BMI, BP, L, SM | EUR | |
| 81 | ANKDD1B7, HMGCR6 | rs12916 | Chr5:75360714 | BC, BMI, L, PSY | EUR | |
| 82 | LNPEP7, ERAP27 | AC009126.12 | rs27660 | Chr5:97012071 | BP, I, L, PSY | Multi |
| 83 | FER11, FBXL174 | LINC010233 | rs288173, rs112949822 | Chr5:108016593-108749489 | BH, BMI, BP, I | EUR, Multi |
| 84 | NREP1, WDR361 | rs256277 | Chr5:112071113 | Multi | ||
| 85 | DMXL12, TNFAIP81 | rs13169691 | Chr5:119112584 | EUR | ||
| 86 | SRFBP15, LOX3 | AC008568.12 | rs4345341 | Chr5:121943056 | BP | EUR |
| 87 | PRDM63, SNX241 | rs73796819 | Chr5:123084735 | Multi | ||
| 88 | FBN28, SLC27A62 | rs6883598 | Chr5:128590497 | BP | EUR | |
| 89 | IRF14, RAD504 | rs10477741, rs2706399 | Chr5:132459618-132532010 | L | EUR | |
| 90 | FGF16, SPRY42 | rs249760, rs3776307 | Chr5:142536127-143114600 | L | EUR | |
| 91 | SGCD4, MED72 | rs39929 | Chr5:156696640 | L, BP | Multi | |
| 92 | ADAM195, FNDC91 | rs11465228 | Chr5:157575687 | Multi | ||
| 93 | FOXC15, GMDS3 | rs72836800 | Chr6:1617092 | BH | EUR | |
| 94 | BLOC1S51, TXNDC51 | rs6597292 | Chr6:7975026 | Multi | ||
| 95 | PHACTR16, GFOD12 | rs9349379 | Chr6:12903725 | ALC, BC, BP, C | EUR, HISP, JPN | |
| 96 | HDGFL14, PRL1 | rs6909752 | Chr6:22612400 | AF | EUR | |
| 97 | HLA-A4, HLA-G3 | HLA-F-AS15, HLA-H3 | rs29266 | Chr6:29607502 | L | EUR |
| 98 | MICA4, PPP1R104 | rs139141104, rs9266631 | Chr6:31021244-31379121 | L | EUR, JPN | |
| 99 | CLIC13, DDAH23 | SNHG322, LOC1005075472 | rs3130683 | Chr6:31920590 | BH, BMI, BP, L, PSY | EUR |
| 100 | LEMD23, UQCC23 | rs62397561 | Chr6:33816328 | Multi | ||
| 101 | BLTP3A9, SNRPC8 | ILRUN-AS12 | rs9469899, rs17609940 | Chr6:34825347-35067023 | BH, BMI, L, PSY | EUR |
| 102 | CDKN1A5, ETV72 | DINOL2 | rs1321309 | Chr6:36670859 | BP, I, L | EUR |
| 103 | KCNK510, SAYSD11 | rs733701, rs10947789 | Chr6:39204086-39207146 | BP | EUR | |
| 104 | PEX63, PTK71 | rs1034246 | Chr6:43100632 | EUR | ||
| 105 | VEGFA4, MRPS18A1 | rs6905288, rs149372871 | Chr6:43791136-44080314 | ALC, BMI, BP, L, PSY | EUR, HISP, JPN | |
| 106 | RAB2310, PRIM26 | LOC1005061884, RP11-203B9.42 | rs11752218, rs9367716 | Chr6:57280764-57295774 | EUR | |
| 107 | CD1096 | CD109-AS12 | rs79717953, rs56171536 | Chr6:73695048-73706145 | JPN, Multi | |
| 108 | TENT5A4, IBTK1 | LINC0254`22 | rs9361867 | Chr6:81886242 | EUR | |
| 109 | UFL16, FHL55 | rs2983896 | Chr6:96581995 | BP | EUR | |
| 110 | CENPW8, TRMT112 | rs35510806 | Chr6:126357186 | BH, I, L, PSY | EUR | |
| 111 | TCF2113, SLC2A127 | TARID3 | rs6919211, rs2327426, rs2492304 | Chr6:133678730-134057013 | BH, BP | EUR, HISP, JPN |
| 112 | PLEKHG14, MTHFD1L2 | RAET1K2 | rs17080093 | Chr6:150676304 | BP | EUR |
| 113 | PLG29, LPA19 | LPAL22 | rs6932293, rs186696265, rs56195640, rs192425087, rs28867400, rs932631509, rs10455872, rs73596816, rs79018195, rs1998043, rs4252120, rs184278183 | Chr6:160114846-160756724 | BC, BMI, BP, C, L, PSY | BLK, EUR, HISP, JPN |
| 114 | MAD1L16, ELFN12 | rs62435159 | Chr7:1871645 | BMI | EUR | |
| 115 | DAGLB8, RAC17 | rs10951983 | Chr7:6406396 | I, L | EUR | |
| 116 | TMEM106B7, THSD7A2 | rs11509880 | Chr7:12222285 | L, PSY | EUR | |
| 117 | ETV14 | rs2073533 | Chr7:13990114 | EUR | ||
| 118 | TWIST15, HDAC92 | rs2107595 | Chr7:19009765 | BP | EUR, JPN | |
| 119 | MACC12, ITGB82 | rs10486389 | Chr7:20260793 | EUR | ||
| 120 | TBX208, DPY19L12 | DPY19L2P12 | rs2215614 | Chr7:35237481 | BH | EUR |
| 121 | CCM25, TBRG41 | rs2107732 | Chr7:45038379 | BP | EUR | |
| 122 | SEMA3C5 | rs917191 | Chr7:80941555 | BMI | Multi | |
| 123 | ZKSCAN16, AP4M15 | STAG3L5P2 | rs35146811 | Chr7:100123371 | BH, BP, L | EUR |
| 124 | COG59, BCAP298 | WBP1LP23, RPL7P322 | rs12112877, rs10953541 | Chr7:107300879-107604100 | L, PSY, SM | EUR |
| 125 | CFTR3, ASZ12 | rs13222797 | Chr7:117459992 | BMI, L | EUR | |
| 126 | ZC3HC15, KLHDC102 | rs11556924 | Chr7:130023656 | BC, BP, I, L, PSY, SM | EUR | |
| 127 | PARP122, HIPK21 | rs5887949 | Chr7:140060741 | BH | EUR | |
| 128 | NOS35, KCNH21 | rs3918226 | Chr7:150993088 | ALC, BC, BP, L, SM | EUR | |
| 129 | MSR13, CNOT71 | rs1480146 | Chr8:16272064 | Multi | ||
| 130 | NAT23, PCM13 | rs4646249 | Chr8:18402921 | L | EUR | |
| 131 | LPL14, CSGALNACT13 | rs894211, rs66778572, rs268 | Chr8:19956018-20058609 | ALC, BC, BH, BMI, BP, I, L, PSY, SM | BLK, EUR, HISP | |
| 132 | BMP15, PHYHIP2 | rs56408342 | Chr8:22190977 | EUR | ||
| 133 | DOCK56, KCTD92 | rs1510758 | Chr8:25204291 | EUR | ||
| 134 | PPP2R2A2 | rs6557894 | Chr8:26229397 | BH | EUR | |
| 135 | RP15 | rs9298506 | Chr8:54524964 | ALC, BH, I, L | Multi | |
| 136 | C8orf342 | rs2380472 | Chr8:68519476 | JPN | ||
| 137 | GEM6, CDH172 | rs34917849 | Chr8:94266079 | BP | EUR | |
| 138 | NDUFAF65, TP53INP11 | rs3019173 | Chr8:95008746 | Multi | ||
| 139 | rs10093110 | Chr8:105553186 | EUR | |||
| 140 | TRIB14, NSMCE21 | rs2001846 | Chr8:125466208 | ALC, BMI, BP, D, I, L | EUR, HISP | |
| 141 | NFIB2, MPDZ1 | LURAP1L-AS12 | rs10961206 | Chr9:13724052 | EUR | |
| 142 | ACER27, RPS61 | AL391834.12 | rs10811183 | Chr9:19436057 | L | EUR |
| 143 | CDKN2A13, CDKN2B11 | ERVFRD-36, MIR31HG4 | rs11523031, rs2891168, rs6475608, rs76959412 | Chr9:21843843-22101703 | BC, BP, C, L | BLK, EUR, HISP, JPN |
| 144 | WNK23, CARD192 | rs4617217 | Chr9:93226686 | Multi | ||
| 145 | ABCA12, NIPSNAP3B2 | rs35093463 | Chr9:104823957 | L | JPN | |
| 146 | KLF44, RAD23B1 | rs1967604 | Chr9:107768043 | BC, L | EUR | |
| 147 | SVEP14 | rs111245230 | Chr9:110407495 | BC, BP | EUR | |
| 148 | ASTN22 | rs7873551 | Chr9:116482848 | BH, PSY | Multi | |
| 149 | PHF195, C54 | rs41312891 | Chr9:120964472 | BH | EUR, Multi | |
| 150 | DAB2IP5, STOM1 | CUTALP2 | rs885150 | Chr9:121657894 | BC, BH, BMI, L | EUR |
| 151 | ABO6, CEL3 | rs651007 | Chr9:133278431 | BC, BP, D, I, L, PSY | BLK, EUR, HISP | |
| 152 | CAMK1D4, CDC1232 | rs17566555 | Chr10:12233948 | PSY | EUR | |
| 153 | ARHGAP214, GPR1581 | rs7077962 | Chr10:24765745 | Multi | ||
| 154 | JCAD7, SVIL1 | rs9337951 | Chr10:30028144 | BP, SM | EUR, HISP | |
| 155 | ZEB16, ARHGAP121 | ZEB1-AS15, PPIAP312 | rs161231 | Chr10:31466991 | L | Multi |
| 156 | CXCL1212, RASGEF1A2 | FRG1JP2 | rs1870634, rs494207, rs527052 | Chr10:43985363-44245808 | C | BLK, EUR, HISP |
| 157 | BICC17, TFAM1 | rs10740732 | Chr10:58593128 | BP | Multi | |
| 158 | TSPAN147, SH2D4B2 | LOC1019295742 | rs17680741 | Chr10:80491758 | EUR | |
| 159 | LIPA6, IFIT11 | rs1051338 | Chr10:89247603 | I, L | EUR, JPN | |
| 160 | PLCE112, SLC35G12 | rs1223583, rs55753709 | Chr10:93996743-94269413 | BMI, BP, L, PSY | EUR | |
| 161 | LOXL49, R3HCC1L3 | rs884811 | Chr10:98164006 | BH, BP | EUR | |
| 162 | CNNM28, BORCS75 | rs77787671 | Chr10:103016448 | BMI, BP, PSY | EUR, HISP, JPN | |
| 163 | STN13, SH3PXD2A2 | rs2067831 | Chr10:103883465 | BP, SM | EUR | |
| 164 | TCF7L25 | rs7903146 | Chr10:112998590 | BH, BMI, BP, D, L, SM | Multi | |
| 165 | C10orf823, GFRA11 | rs2218584 | Chr10:116649131 | L | Multi | |
| 166 | WDR111 | rs17101521 | Chr10:121156039 | BMI, BP, L | Multi | |
| 167 | HTRA16, TACC22 | rs2672592, rs4752700, rs17103659 | Chr10:122471234-122515087 | EUR, HISP | ||
| 168 | BET1L3, PSMD132 | rs73386640 | Chr11:203235 | JPN | ||
| 169 | TRIM54, TRIM221 | rs11601507 | Chr11:5679844 | BC, L | EUR | |
| 170 | DENND2B2, TRIM662 | LOC1027247842, TMEM9B-AS12 | rs56210063 | Chr11:8767618 | Multi | |
| 171 | SWAP708, TMEM41B5 | LOC4400282, SBF2-AS12 | rs4537761, rs360153 | Chr11:9301806-9740727 | BC, BP, D | EUR |
| 172 | CTR92 | MRVl12 | rs11042937 | Chr11:10723847 | EUR | |
| 173 | BMAL16, ARNTL2 | rs11316597 | Chr11:13274205 | BP, PSY | EUR | |
| 174 | BDNF2, LIN7C2 | LINC006782 | rs6265 | Chr11:27658369 | ALC, BMI, BP, I, L, PSY, SM | Multi |
| 175 | WT14, IMMP1L2 | rs7118294 | Chr11:32358975 | BH, BP, L | EUR | |
| 176 | HSD17B127, TTC171 | rs7116641 | Chr11:43675367 | BH, BMI, I | EUR | |
| 177 | SIPA18, RNASEH2C3 | rs2306363 | Chr11:65638129 | ALC, BMI, BP, L | EUR, JPN | |
| 178 | SERPINH117, MAP64 | AP001922.52 | rs590121, rs584961 | Chr11:75563105-75566583 | BH, BMI | EUR |
| 179 | ARHGAP4211, PGR1 | LOC1001283862 | rs633185, rs7947761 | Chr11:100722807-100753868 | ALC, BP | EUR |
| 180 | MMP132, MMP31 | rs1892971 | Chr11:102924877 | EUR | ||
| 181 | PDGFD5, DYNC2H11 | rs2839812 | Chr11:103802566 | BMI, BP | EUR, JPN | |
| 182 | CWF19L25, GUCY1A22 | rs7117780 | Chr11:107215397 | BP | EUR | |
| 183 | RDX5, FDX14 | rs10488763 | Chr11:110373636 | EUR, JPN | ||
| 184 | SIK27, CRYAB5 | rs11213945 | Chr11:111590279 | BP | Multi | |
| 185 | ZPR13, BUD132 | rs964184 | Chr11:116778201 | L | EUR, HISP | |
| 186 | VPS111, MCAM1 | rs1177562 | Chr11:119078621 | L | EUR | |
| 187 | TLCD53, ARHGEF122 | rs4938809 | Chr11:120493228 | EUR | ||
| 188 | UBASH3B6, JHY1 | rs57812455 | Chr11:122654259 | BC, BP, L | EUR | |
| 189 | ST3GAL45, TIRAP2 | GSEC3 | rs10790800 | Chr11:126392743 | L | EUR |
| 190 | C1S4, CHD42 | rs72447384 | Chr12:7071137 | I | EUR | |
| 191 | YBX36, OLR11 | rs2607903 | Chr12:10723974 | JPN | ||
| 192 | PDE3A4, AEBP22 | rs10841443 | Chr12:20067099 | BP | EUR | |
| 193 | ZCRB17, PPHLN17 | rs12820154 | Chr12:42389305 | BP | Multi | |
| 194 | ZNF6414, CCDC1842 | rs1859440 | Chr12:48033546 | BC, BP | EUR | |
| 195 | SMUG13, HOXC43 | FLJ128252, RP11-834C11.42 | rs1133773, rs75160195 | Chr12:54102435-54127810 | EUR, JPN | |
| 196 | COQ10A6, STAT24 | rs143405823 | Chr12:56329935 | BH, L, PSY | Multi | |
| 197 | LRP17, STAT65 | rs11172113 | Chr12:57133500 | BH, BMI, BP, L, PSY | EUR | |
| 198 | ATP2B15, GALNT43 | AC010201.22 | rs2681472 | Chr12:89615182 | ALC, BC, BMI, BP, PSY | EUR, JPN |
| 199 | FGD67, VEZT3 | rs11107903 | Chr12:95114195 | BC | EUR, JPN | |
| 200 | CORO1C4, SELPLG1 | rs1558802 | Chr12:108642472 | L, PSY | Multi | |
| 201 | ATXN24, SH2B32 | rs10774625, rs11066015 | Chr12:111472415-111730205 | ALC, BC, BMI, BP, I, L, PSY, SM | EUR, JPN | |
| 202 | TBX34, TBX51 | rs34606058 | Chr12:114915563 | BC, BH, BP | EUR, Multi | |
| 203 | KSR24, FBXW81 | rs11830157 | Chr12:117827636 | EUR | ||
| 204 | HNF1A6, C12orf435 | RNU4-22, HNF1A-AS12 | rs2244608 | Chr12:120979185 | ALC, BC, I, L, PSY | EUR |
| 205 | ZNF6646, CCDC9211 | rs7133378 | Chr12:123924955 | BMI, D, L, PSY, SM | EUR | |
| 206 | SCARB111, UBC2 | rs7485656, rs7296737 | Chr12:124831101-124852410 | I, L | BLK, EUR | |
| 207 | ZNF2682, ZNF1402 | AC131212.32 | rs36127550 | Chr12:133203723 | BMI, BP | Multi |
| 208 | FLT15, SLC46A32 | rs17086617, rs74412485 | Chr13:28388549-28468295 | BH, BP | EUR, JPN | |
| 209 | PDS5B8, N4BP2L25 | rs7991314 | Chr13:32551937 | BC, BH, BMI, SM | EUR | |
| 210 | DOCK93, SLC15A11 | rs8000794 | Chr13:98782556 | EUR | ||
| 211 | COL4A119, COL4A212 | rs11617955, rs3783113, rs11619113, rs4773141, rs9515203, rs7333991 | Chr13:110165755-110456089 | BP, C, L | BLK, EUR, JPN | |
| 212 | F102, MCF2L1 | rs4907571 | Chr13:112964182 | BP | EUR | |
| 213 | ARID4A2, ARMH42 | PSMA3-AS12 | rs2145598 | Chr14:58327283 | L | EUR |
| 214 | EIF2B27, MLH36 | rs10131894 | Chr14:74980176 | EUR | ||
| 215 | ITPK14, ATXN32 | rs1956028 | Chr14:93040852 | BC | Multi | |
| 216 | SERPINA14, DDX241 | rs112635299 | Chr14:94371805 | ALC, BH, BMI, BP, I, L, PSY | EUR | |
| 217 | HHIPL111, CYP46A15 | AL160313.12 | rs36033161, rs2895811 | Chr14:99657150-99667605 | BP | EUR, JPN |
| 218 | EXD12, KNSTRN2 | OIP5-AS13 | rs3959554 | Chr15:41151726 | BMI, BP | EUR |
| 219 | NEDD43, PIGB3 | AC018926.12 | rs11633313 | Chr15:55843511 | Multi | |
| 220 | LIPC5, ALDH1A25 | HSP90AB4P2 | rs588136 | Chr15:58438299 | L, PSY | EUR |
| 221 | TRIP45, RBPMS23 | rs6494488 | Chr15:64732005 | L | EUR | |
| 222 | SMAD37, SMAD63 | IQCH-AS12 | rs56062135 | Chr15:67163292 | EUR, HISP | |
| 223 | ADAMTS713, CTSH10 | AC090607.22 | rs7177201, rs7173743, rs112238647, rs7174727 | Chr15:78759363-78858944 | BH, BP,C, SM | EUR, HISP, JPN |
| 224 | CFAP1613, MESD2 | rs2683260 | Chr15:81093211 | EUR | ||
| 225 | AKAP133, KLHL253 | LOC1019296792, AC044860.12 | rs7177107 | Chr15:85580133 | BC | EUR |
| 226 | MFGE85, ABHD24 | rs1807214 | Chr15:89022026 | BP | EUR, JPN | |
| 227 | FES9, FURIN6 | rs7183988 | Chr15:90885359 | BP, L, PSY | BLK, EUR, JPN | |
| 228 | NR2F22 | rs17581137 | Chr15:95603185 | EUR | ||
| 229 | MYH115, NOMO12 | AC138969.12 | rs12691049 | Chr16:15815656 | AF, BP | EUR |
| 230 | FTO5, AKTIP1 | rs1421085 | Chr16:53767042 | ALC, BH, BMI, BP, D, I, L, PSY, SM | BLK, EUR | |
| 231 | CETP6, HERPUD13 | rs12446515 | Chr16:56953103 | AF, BMI, BP, C, L | EUR | |
| 232 | NOB13, NFAT52 | PDXDC2P3 | rs9302604 | Chr16:69542991 | BP, PSY, SM | EUR |
| 233 | DHODH7, DHX383 | rs12445401 | Chr16:72114520 | ALC, L, PSY | EUR | |
| 234 | CFDP16, TMEM170A5 | AC009054.22 | rs8046696 | Chr16:75408245 | ALC, BC, BH, BP, L, SM | EUR, JPN |
| 235 | CMIP8, PLCG26 | AC009148.12 | rs10493891, rs7189462 | Chr16:81477137-81874262 | BP, L | EUR |
| 236 | CDH134, OSGIN11 | rs7500448 | Chr16:83012185 | BP | EUR | |
| 237 | rs55880988 | Chr16:88479845 | L, I | Multi | ||
| 238 | SMG65, SRR3 | rs4790881 | Chr17:2165638 | L | EUR | |
| 239 | TOM1L27, DRC37 | CCDC144BP2, USP32P22 | rs12936927 | Chr17:17823651 | ALC, BMI, PSY, SM | EUR, HISP |
| 240 | SSH28, CORO65 | SH3GL1P22 | rs11080107 | Chr17:29611406 | BH, BMI | EUR |
| 241 | COPRS2, NF11 | rs76954792 | Chr17:31706495 | BP | EUR | |
| 242 | FBXL207, MED15 | rs1077715 | Chr17:39527295 | ALC, BC, L, SM | EUR | |
| 243 | CAVIN18, DHX586 | NBR24, AC099811.53 | rs12952244, rs8068844, rs9912587 | Chr17:42118063-43021069 | BMI, D, I, L | EUR, Multi |
| 244 | ARHGAP275, GOSR25 | LINC022102, NSFP12 | rs5026246, rs1706731, rs1881193, rs1881194, rs2532235, rs2696671, rs17608766 | Chr17:45495530-46935905 | AF, BP, PSY | EUR, HISP |
| 245 | SNF810, ATP5MC110 | SUMO2P174, LOC1053718142 | rs46522, rs4643373, rs5820757 | Chr17:48911235-49340581 | ALC, BH, BMI, BP, I, L, PSY | EUR |
| 246 | BCAS34, PPM1D2 | RPL12P382 | rs11655024 | Chr17:61155004 | BMI, BP | EUR, JPN |
| 247 | PECAM15, ERN12 | AC046185.32 | rs11079536 | Chr17:64315043 | BC, BP | EUR, JPN |
| 248 | WIPI16, BPTF3 | rs2909217 | Chr17:68467844 | L | EUR | |
| 249 | TRIM4710, UNC13D8 | ATF4P33 | rs2410859, rs78532451 | Chr17:75845204-75883011 | BP, L | EUR, Multi |
| 250 | rs112735431 | Chr17:80385145 | JPN | |||
| 251 | CTAGE13, GATA63 | rs9951447 | Chr18:22429728 | EUR, JPN | ||
| 252 | NPC14, RMC14 | rs1623003 | Chr18:23585199 | ALC, BP, L, SM | EUR | |
| 253 | rs4327120 | Chr18:38953012 | SM | Multi | ||
| 254 | LIPG5, SMAD74 | AC022919.12 | rs9945890, rs12965923 | Chr18:48989546-49687312 | L | EUR |
| 255 | LMAN16, MC4R5 | rs11663411, rs476828 | Chr18:59293278-60185354 | ALC, BH, BMI, BP, D, I, L, PSY, SM | EUR | |
| 256 | rs150651184 | Chr19:282148 | BLK | |||
| 257 | DOT1L6, PLEKHJ15 | AC006130.32 | rs191615952 | Chr19:2236098 | BH, BP | Multi |
| 258 | KDM4B5, TICAM12 | rs12185519 | Chr19:4967728 | BH, BMI, L | Multi | |
| 259 | ANGPTL46, MAP2K71 | rs116843064 | Chr19:8364439 | ALC, BMI, L | EUR | |
| 260 | LDLR18, SMARCA46 | ZNF433-AS14, AC008770.32 | rs55997232, rs10422256, rs72658867, rs34774090, rs167479 | Chr19:11077441-11416089 | ALC, BC, BP, L, PSY | BLK, EUR, HISP, JPN |
| 261 | CYP4F22, EPS15L12 | rs77420750 | Chr19:15868934 | EUR | ||
| 262 | KLF26, AP1M12 | rs10420373 | Chr19:16319866 | I | JPN | |
| 263 | MYO9B6, HAUS85 | rs7246865 | Chr19:17108295 | BC, BMI, BP | EUR | |
| 264 | MAP1S9, FCHO16 | rs10410487 | Chr19:17718799 | BC, BH, I | EUR | |
| 265 | ELL2, ISYNA12 | rs78030362 | Chr19:18464383 | BC, BMI, BP | EUR | |
| 266 | ZNF1007, ZNF4933 | rs8102320 | Chr19:21754083 | BP | Multi | |
| 267 | ZNF5078, DPY19L31 | rs12976411 | Chr19:32391114 | EUR | ||
| 268 | EIF3K2, ZFP302 | ZNF7815, LINC015352 | rs73025613 | Chr19:37843721 | Multi | |
| 269 | TGFB115, B9D26 | AC011462.53 | rs60315715, rs11466359, rs1800469 | Chr19:41284182-41354391 | BC, BP | EUR, HISP, JPN |
| 270 | APOE12, APOC18 | rs429358, rs7412, rs183657985 | Chr19:44908684-45239240 | BC, BH, BP, C, I, L | BLK, EUR, JPN | |
| 271 | RSPH6A4, DMWD4 | PPP5D1P2, GEMIN7-AS12 | rs8108474 | Chr19:45798221 | BH, BP, L | EUR |
| 272 | RRBP16, DSTN3 | rs1132274 | Chr20:17615510 | BH, L | EUR, Multi | |
| 273 | ITCH7, NCOA64 | FER1L42 | rs6088595 | Chr20:34770696 | BC, BH, BMI, D, I, L, SM | EUR |
| 274 | PROCR7, EDEM24 | NORAD2 | rs17406518 | Chr20:35211373 | BC | EUR |
| 275 | MAFB4 | rs2207132 | Chr20:40513876 | BMI, I, L | EUR | |
| 276 | PLCG16, TOP14 | rs6102343 | Chr20:41295639 | BMI, L | EUR | |
| 277 | PLTP3, CD403 | rs8124182 | Chr20:45980262 | L | EUR | |
| 278 | PREX15, ARFGEF22 | RN7SL197P2 | rs2008614 | Chr20:48816613 | BP | EUR |
| 279 | rs4809766 | Chr20:49942802 | EUR | |||
| 280 | GNAS3, ZNF8312 | APCDD1L-DT2, LINC017112 | rs6026739 | Chr20:59164414 | BH, BP | EUR |
| 281 | rs151206628 | Chr21:10592292 | HISP | |||
| 282 | NRIP13, HSPA131 | rs2064040 | Chr21:15212021 | Multi | ||
| 283 | MAP3K7CL5, BACH14 | LINC001893 | rs2832227 | Chr21:29160755 | EUR | |
| 284 | SLC5A36, MRPS63 | AP000318.22 | rs28451064, rs149487184 | Chr21:34221526-34233562 | BC, BMI, BP | EUR |
| 285 | ARVCF8, TANGO25 | DGCR112 | rs4819811, rs71313931 | Chr22:19621733-19972661 | BC, BP, I | EUR |
| 286 | SUSD27, GGT56 | POM121L9P3, GGTLC4P2 | rs12484557, rs5760309 | Chr22:24159893-24281863 | EUR | |
| 287 | OSM4, LIF3 | rs6006426 | Chr22:30273894 | EUR | ||
| 288 | FBXO71 | rs137525 | Chr22:32900153 | Multi | ||
| 289 | SCUBE13, TTLL122 | rs139012 | Chr22:43227966 | EUR | ||
| 290 | PNPLA32, PRR52 | rs738408 | Chr22:43928850 | ALC, BC, L | Multi | |
| 291 | TBL1X | rs5934659 | ChrX:9610064 | EUR | ||
| 292 | OPHN11, AR1 | rs1410127 | ChrX:68060539 | Multi | ||
| 293 | CYSLTR11, TAF9B1 | rs398484 | ChrX:78343972 | Multi | ||
| 294 | VDAC1P1 | rs2066280 | ChrX:80922131 | EUR | ||
| 295 | SETP4 | rs2342572 | ChrX:84814364 | Multi | ||
| 296 | CHRDL11, RGAG11 | rs7884019 | ChrX:110566261 | EUR | ||
| 297 | MAP7D31, FHL11 | rs5929743 | ChrX:136236818 | EUR | ||
| 298 | ARHGEF61 | RP11-308D16.21 | rs5975828 | ChrX:136904390 | EUR | |
| 299 | DNASE1L11, ATP6AP11 | rs147967693 | ChrX:154410919 | Multi |
In total, 393 independent CAD associations were collapsed into 290 autosomal and 9 X-linked genetic loci (locus ID) based on the proximal or most likely causal gene(s), including protein and RNA coding genes. The internal confidence score (cs) for causality is displayed as a superscript number [max. score = 29 for plasminogen (PLG)]. The column ‘index rsIDs' includes independent CAD associations within one gene region. Additional associations for CAD risk factors are shown in column ‘Traits’, including lipids (L), body mass index (BMI), body height (BH), blood pressure (BP), inflammation (I), calcification (C), blood coagulation (BC), diabetes (D), alcohol intake (ALC), smoking (SM), and psychological factors (e.g. sleep) (PSY). The last column shows the main ethnic groups for which CAD risk variants were identified (EUR = European, JPN = Japanese, HISP = Hispanic, BLK = black/African American, and Multi = mixed cohort).
2. Genetic architecture of CAD
2.1 Deciphering the molecular mechanisms of CAD risk variant
Determining the genetic architecture of CAD has proven to be a significant step in understanding disease aetiology and developing new treatment strategies for at-risk individuals. However, deciphering the precise mechanisms of disease-associated variants is always challenging and requires an integrative approach using bioinformatics tools and in vitro models. Figure 2 provides an overview of variant consequences (from Ensembl release 11018) for all 393 index variants and ∼18 500 additional variants in high linkage disequilibrium (LD) (r2 > 0.6). Most CAD-associated variants are in intronic regions and about 57% of these intronic variants reside in cis-regulatory elements (CREs) [e.g. transcription factor (TF) binding sites], indicating that a high proportion of these variants are indeed regulatory and modulate gene expression in a context-specific manner. It is believed that not only one single variant within one locus is linked to disease risk, but rather orchestrated effects of numerous variants can contribute to changes in transcript and protein levels.19–21 In theory, every step of the gene expression machinery can be affected by disease-associated variants, including genome organization (e.g. chromatin state), mRNA processing (e.g. transport or alternative splicing), and protein biosynthesis (e.g. protein modifications) (Figure 2). Examples of molecular mechanisms of CAD risk variants and affected genes (e.g. APOB, SH2B3, LPP, SORT1, FES, FURIN, and GUCY1A1) can be found in these excellent reviews16,17 and studies.22–24 Further examples (EDN1, PLPP3, HDAC9, LMOD1, SMAD3, LIPA, etc.) and a description of experimental methods can be found in this review by Rodriguez et al.17 and in Table 2.
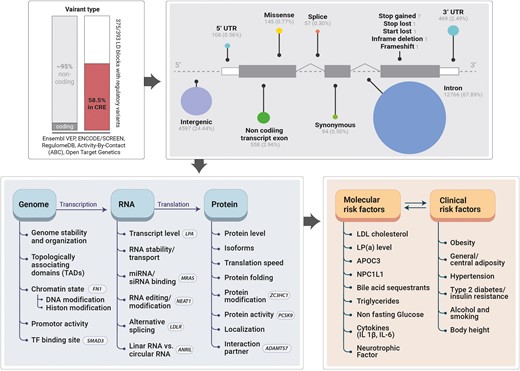
Overview of possible consequences of CAD risk variants. Variant types (top right): about 95% of all ∼18 500 variants in LD with 393 proxy CAD-associated variants (r2 > 0.6) are in the non-coding region. About 60% of all CAD-associated variants in LD have regulatory potential as they either overlap with CREs from ENCODE/SCREEN,25 with VEP enhancer elements (Ensembl Variant Effect Predictor18) rank high in the RegulomeDB (ranking ≤ 2a or probability score > 0.8),26 or have a high score in other data sets (ABC score > 0.04 or Open Targets Genetics score > 0.7). Variant consequences (top right): most variants are in introns and about 56.7% of these intronic variants may show regulatory potential by altering enhancer activity or by impacting mRNA splicing (splice variants). Variant consequences are from the Ensembl VEP.18 Variant mechanisms (bottom left): CAD risk variant can affect gene expression (transcription or translation) as well as protein function. Examples of molecular mechanisms of genes in grey boxes are described in Table 2. CAD risk factors (bottom right): the combination of risk variants can ultimately impact CAD causing molecular risk factors such as LDL cholesterol, which are often associated with clinical risk factors such as obesity. The complete list of causal traits, biomarkers, and diseases derived from Mendelian randomization studies can be found in a review article by Chen and Schunkert.27
Exemplary list of variant consequences and detection methods
| Level | Regulatory feature | Analysis | Description | Example of CAD-associated genes |
|---|---|---|---|---|
| Genome | Chromatin state | ATAC-seq or DNase-seq | Both sequencing techniques allow for the detection of accessible chromatin, a prerequisite for active regulation of gene expression. Variants that affect chromatin accessibility are called caQTLs. | The caQTL variant rs2692224 located downstream of the FN1 gene was identified using ATAC-seq in human coronary artery SMCs28 |
| Enhancer/TF binding sites | ChIP-seq, CRISPR-Cas perturbation | Chromatin immunoprecipitation (ChIP) is combined with DNA-seq. DNA and proteins (histones or TFs) are crosslinked. Fragmented protein-bound DNA is captured using antibodies and sequenced. Variants that alter TF binding are called bQTLs. CREs can be confirmed using CRISPR-Cas perturbation experiments.17 | By combining DNase-seq and histone ChIP-seq data, Turner et al. identified a protective T allele of rs17293632 that disrupts an activator protein-1 site in a SMAD3 intron enhancer, resulting in reduced SMAD3 expression.29 Toropainen et al. also identified this variant in ECs.30 | |
| Chromatin looping | Hi-C-seq | Hi-C-seq combines DNA proximity ligation with high-throughput sequencing in a genome-wide fashion to identify interacting regulatory features such as enhancers and promotors. Variants that affect chromatin looping are called clQTLs. | The caQTL variant rs2692224 in the FN1 locus has also been described to alter chromatin looping in human coronary artery SMCs28 | |
| RNA | RNA level | eQTL, reporter assays | Variants that impact transcript level or gene expression (e) are called eQTLs. To detect eQTLs, genotyping and RNA sequencing are combined followed by association tests. Allele specific activity can be confirmed by reporter assay such as MPRA or STARR-seq.17 | About 85% of CAD loci contain eQTLs (see Supplementary material online, Table S1). The SMAD3 variant rs17293632 is one example in which the eQTL can be explained by the modulation of a TF binding site. However, eQTLs are often cell type and context-specific, i.e. only detectable after adding a certain stimulus. |
| UTR variants | mRNA stability/miRNA binding, etc. | UTR variants can have a variety of functional consequences such as altering the secondary structure, stability, location, translation, or binding of microRNAs or RNA-binding proteins. There are bioinformatics tools to predict mRNA stability or binding of microRNAs (e.g. https://mirdb.org/). | Two CAD-associated variants (rs9818870 and rs40593) are located in the 3′-UTR of the MRAS gene, adjacent to a cluster of miRNA binding sites, thereby most likely affecting mRNA stability and biosynthesis of the MRAS protein31,32 | |
| RNA editing | edQTL | Only a few disease-associated variants have been reported to directly affect RNA modifications or RNA editing, e.g. adenosine-to-inosine conversion. Although it has been shown that variants that affect RNA editing (so-called edQTLs) can contribute to the genetic risk of common inflammatory diseases,33 comprehensive datasets of edQTLs are missing. | Recently, 81 variants associated with CAD or acute MI were identified to affect RNA modifications.34 Four of these variants were genome-wide significant for CAD or acute MI (rs3739998 in JCAD, rs148172130 in RPL14, rs12190287 in TCF21, and rs186643756 in PVT1). | |
| RNA splicing | sQTL | Alternative splicing is critical in atherosclerosis.35 Like eQTL analyses, genotyping and isoform-sensitive RNA sequencing are required to detect splice (s)QTLs that can impact alternative splicing (e.g. exon skipping) and/or isoform usage. | The exon variant rs688 leads to alternative splicing of LDLR mRNA, rapid degradation of the LDLR isoform, and increased LDL cholesterol levels. Rare intronic LDLR variants were also identified to affect splicing in familial hypercholesterolemia patients.36,37 | |
| Circular RNA | circRNAs-seq | Circular (circ)RNAs, products of back-splicing of precursor mRNAs and covalent ligation of the 3′ and 5′ end, are abundantly expressed with high evolutionary conservation, show high tissue specificity, and exert their regulatory functions by miRNA sponging, protein sponging, or direct interaction with proteins of the transcriptional machinery.38 Detection methods for circRNAs can be found in these reviews.39,40 | CircRNAs have been linked to various cardiovascular diseases38,41 (e.g. circANRIL, circSATB2, circRUSC2, or circCHFR).38,42 The circular version of the antisense non-coding RNA at the INK4 locus (circANRIL) transcribed on the 9p21 locus is more abundant in carriers of the CAD-protective haplotype. The protective effect is thought to be due to circANRIL-mediated disturbed maturation of precursor rRNA, thereby inducing nucleolar stress and apoptosis in vascular SMCs and macrophages.16,43 | |
| Protein | Protein level | pQTL | Variants that impact protein level are called pQTLs, which are identified through association tests of genotypes and protein abundance in certain tissues or cell types. | A dose–response relationship has been described for lipoprotein(a), with protective CAD variants lowering lipoprotein(a) level, whereas higher levels of lipoprotein(a) were found in individuals carrying CAD-causal variants12 |
| Protein function | Enzyme activity | Missense variants that change the amino sequence of a protein can alter protein function in many ways. Therefore, enzyme specific functional assays must be carefully designed to confirm the impact of a missense variant. However, a variety of bioinformatics prediction tools are available (e.g. LoFtool44 or AlphaMissense.45 | At least 20 rare loss-of-function PCSK9 variants46 and 1 common missense variant in PCSK9 have been reported.7 The consequences of these variants can range from degradation of PCSK9, compromised intracellular transport, aberrant autocatalysis, or reduced affinity for the LDLR on the liver surface which is normally targeted through PCSK9 to lysosomal degradation after internalization of the LDLR-LDL complex. | |
| Protein modification | Mass spectrometry (MS), Western blot analysis (WB) | Missense variants that alter post-translational modifications have been reported frequently. MS is widely used to detect post-translational modification such as phosphorylation. WB can confirm such findings but is less sensitive. | A coding variant rs11556924-T (p.Arg363His) of ZC3HC1 (involved in cell cycle) results in enhanced phosphorylation at serin 395 of the ZC3HC1 protein, thereby reducing its activity.47 Of note, synonymous variants can have functional effects on mRNA stability and may contribute to co-translational misfolding or changes in co-translational modifications such as glycosylation.48 | |
| Interaction partner | Crosslinking mass spectrometry (clMS), co-immunoprecipitation (Co-IP), fluorescence resonance energy transfer (FRET), yeast two-hybrid screening (Y2H) | Missense variants can affect the binding affinity to binding partners with multiple consequences, depending on the nature of binding partners (e.g. proteinases or substrates). To detect protein–protein interactions, several methods are available such as high-throughput clMS49 as well as targeted approaches such as co-IP, FRET, Y2H, and others.50 | Due to its size, the ADAMTS7 protein has many interaction partners, such as activators (e.g. furin51,52), inhibitors (e.g. PRGN48 or A2M53) and substrates (e.g. TSP-1,54 TIMP1,55 SVEP1,56 or COMP57). The protective minor allele rs3825807-G (p.Ser214Pro) hampers furin-dependent cleavage of the pro-domain of ADAMTS7, resulting in lower level of active ADAMST7 |
| Level | Regulatory feature | Analysis | Description | Example of CAD-associated genes |
|---|---|---|---|---|
| Genome | Chromatin state | ATAC-seq or DNase-seq | Both sequencing techniques allow for the detection of accessible chromatin, a prerequisite for active regulation of gene expression. Variants that affect chromatin accessibility are called caQTLs. | The caQTL variant rs2692224 located downstream of the FN1 gene was identified using ATAC-seq in human coronary artery SMCs28 |
| Enhancer/TF binding sites | ChIP-seq, CRISPR-Cas perturbation | Chromatin immunoprecipitation (ChIP) is combined with DNA-seq. DNA and proteins (histones or TFs) are crosslinked. Fragmented protein-bound DNA is captured using antibodies and sequenced. Variants that alter TF binding are called bQTLs. CREs can be confirmed using CRISPR-Cas perturbation experiments.17 | By combining DNase-seq and histone ChIP-seq data, Turner et al. identified a protective T allele of rs17293632 that disrupts an activator protein-1 site in a SMAD3 intron enhancer, resulting in reduced SMAD3 expression.29 Toropainen et al. also identified this variant in ECs.30 | |
| Chromatin looping | Hi-C-seq | Hi-C-seq combines DNA proximity ligation with high-throughput sequencing in a genome-wide fashion to identify interacting regulatory features such as enhancers and promotors. Variants that affect chromatin looping are called clQTLs. | The caQTL variant rs2692224 in the FN1 locus has also been described to alter chromatin looping in human coronary artery SMCs28 | |
| RNA | RNA level | eQTL, reporter assays | Variants that impact transcript level or gene expression (e) are called eQTLs. To detect eQTLs, genotyping and RNA sequencing are combined followed by association tests. Allele specific activity can be confirmed by reporter assay such as MPRA or STARR-seq.17 | About 85% of CAD loci contain eQTLs (see Supplementary material online, Table S1). The SMAD3 variant rs17293632 is one example in which the eQTL can be explained by the modulation of a TF binding site. However, eQTLs are often cell type and context-specific, i.e. only detectable after adding a certain stimulus. |
| UTR variants | mRNA stability/miRNA binding, etc. | UTR variants can have a variety of functional consequences such as altering the secondary structure, stability, location, translation, or binding of microRNAs or RNA-binding proteins. There are bioinformatics tools to predict mRNA stability or binding of microRNAs (e.g. https://mirdb.org/). | Two CAD-associated variants (rs9818870 and rs40593) are located in the 3′-UTR of the MRAS gene, adjacent to a cluster of miRNA binding sites, thereby most likely affecting mRNA stability and biosynthesis of the MRAS protein31,32 | |
| RNA editing | edQTL | Only a few disease-associated variants have been reported to directly affect RNA modifications or RNA editing, e.g. adenosine-to-inosine conversion. Although it has been shown that variants that affect RNA editing (so-called edQTLs) can contribute to the genetic risk of common inflammatory diseases,33 comprehensive datasets of edQTLs are missing. | Recently, 81 variants associated with CAD or acute MI were identified to affect RNA modifications.34 Four of these variants were genome-wide significant for CAD or acute MI (rs3739998 in JCAD, rs148172130 in RPL14, rs12190287 in TCF21, and rs186643756 in PVT1). | |
| RNA splicing | sQTL | Alternative splicing is critical in atherosclerosis.35 Like eQTL analyses, genotyping and isoform-sensitive RNA sequencing are required to detect splice (s)QTLs that can impact alternative splicing (e.g. exon skipping) and/or isoform usage. | The exon variant rs688 leads to alternative splicing of LDLR mRNA, rapid degradation of the LDLR isoform, and increased LDL cholesterol levels. Rare intronic LDLR variants were also identified to affect splicing in familial hypercholesterolemia patients.36,37 | |
| Circular RNA | circRNAs-seq | Circular (circ)RNAs, products of back-splicing of precursor mRNAs and covalent ligation of the 3′ and 5′ end, are abundantly expressed with high evolutionary conservation, show high tissue specificity, and exert their regulatory functions by miRNA sponging, protein sponging, or direct interaction with proteins of the transcriptional machinery.38 Detection methods for circRNAs can be found in these reviews.39,40 | CircRNAs have been linked to various cardiovascular diseases38,41 (e.g. circANRIL, circSATB2, circRUSC2, or circCHFR).38,42 The circular version of the antisense non-coding RNA at the INK4 locus (circANRIL) transcribed on the 9p21 locus is more abundant in carriers of the CAD-protective haplotype. The protective effect is thought to be due to circANRIL-mediated disturbed maturation of precursor rRNA, thereby inducing nucleolar stress and apoptosis in vascular SMCs and macrophages.16,43 | |
| Protein | Protein level | pQTL | Variants that impact protein level are called pQTLs, which are identified through association tests of genotypes and protein abundance in certain tissues or cell types. | A dose–response relationship has been described for lipoprotein(a), with protective CAD variants lowering lipoprotein(a) level, whereas higher levels of lipoprotein(a) were found in individuals carrying CAD-causal variants12 |
| Protein function | Enzyme activity | Missense variants that change the amino sequence of a protein can alter protein function in many ways. Therefore, enzyme specific functional assays must be carefully designed to confirm the impact of a missense variant. However, a variety of bioinformatics prediction tools are available (e.g. LoFtool44 or AlphaMissense.45 | At least 20 rare loss-of-function PCSK9 variants46 and 1 common missense variant in PCSK9 have been reported.7 The consequences of these variants can range from degradation of PCSK9, compromised intracellular transport, aberrant autocatalysis, or reduced affinity for the LDLR on the liver surface which is normally targeted through PCSK9 to lysosomal degradation after internalization of the LDLR-LDL complex. | |
| Protein modification | Mass spectrometry (MS), Western blot analysis (WB) | Missense variants that alter post-translational modifications have been reported frequently. MS is widely used to detect post-translational modification such as phosphorylation. WB can confirm such findings but is less sensitive. | A coding variant rs11556924-T (p.Arg363His) of ZC3HC1 (involved in cell cycle) results in enhanced phosphorylation at serin 395 of the ZC3HC1 protein, thereby reducing its activity.47 Of note, synonymous variants can have functional effects on mRNA stability and may contribute to co-translational misfolding or changes in co-translational modifications such as glycosylation.48 | |
| Interaction partner | Crosslinking mass spectrometry (clMS), co-immunoprecipitation (Co-IP), fluorescence resonance energy transfer (FRET), yeast two-hybrid screening (Y2H) | Missense variants can affect the binding affinity to binding partners with multiple consequences, depending on the nature of binding partners (e.g. proteinases or substrates). To detect protein–protein interactions, several methods are available such as high-throughput clMS49 as well as targeted approaches such as co-IP, FRET, Y2H, and others.50 | Due to its size, the ADAMTS7 protein has many interaction partners, such as activators (e.g. furin51,52), inhibitors (e.g. PRGN48 or A2M53) and substrates (e.g. TSP-1,54 TIMP1,55 SVEP1,56 or COMP57). The protective minor allele rs3825807-G (p.Ser214Pro) hampers furin-dependent cleavage of the pro-domain of ADAMTS7, resulting in lower level of active ADAMST7 |
Abbreviations: ADAMTS7, a disintegrin and metalloproteinase with thrombospondin motifs 7; ATAC-seq, assay for transposase-accessible chromatin with high-throughput sequencing; A2M, alpha-2-macroglobulin; ChIP-seq, chromatin immunoprecipitation sequencing; CRISPR, clustered regularly interspaced short palindromic repeats; COMP, cartilage oligomeric matrix protein; DNase-seq, Dnase I hypersensitive sites sequencing; ECs, endothelial cells; FN1, fibronectin 1; LDLR, low density lipoprotein receptor; MPRA, massively parallel reporter assay; MRAS, muscle RAS oncogene homologue; PCSK9, proprotein convertase subtilisin/kexin type 9; PRGN, progranulin; QTL, quantitative trait loci; rRNA, ribosomal RNA; SMAD3, SMAD family member 3; SMCs, smooth muscle cells; STARR-seq, self-transcribing active regulatory region sequencing; SVEP1, Sushi, von Willebrand factor type A, EGF and pentraxin domain-containing protein 1; TIMP1, tissue inhibitor of metalloproteinases 1; TSP-1, thrombospondin-1; UTR, untranslated region.
Exemplary list of variant consequences and detection methods
| Level | Regulatory feature | Analysis | Description | Example of CAD-associated genes |
|---|---|---|---|---|
| Genome | Chromatin state | ATAC-seq or DNase-seq | Both sequencing techniques allow for the detection of accessible chromatin, a prerequisite for active regulation of gene expression. Variants that affect chromatin accessibility are called caQTLs. | The caQTL variant rs2692224 located downstream of the FN1 gene was identified using ATAC-seq in human coronary artery SMCs28 |
| Enhancer/TF binding sites | ChIP-seq, CRISPR-Cas perturbation | Chromatin immunoprecipitation (ChIP) is combined with DNA-seq. DNA and proteins (histones or TFs) are crosslinked. Fragmented protein-bound DNA is captured using antibodies and sequenced. Variants that alter TF binding are called bQTLs. CREs can be confirmed using CRISPR-Cas perturbation experiments.17 | By combining DNase-seq and histone ChIP-seq data, Turner et al. identified a protective T allele of rs17293632 that disrupts an activator protein-1 site in a SMAD3 intron enhancer, resulting in reduced SMAD3 expression.29 Toropainen et al. also identified this variant in ECs.30 | |
| Chromatin looping | Hi-C-seq | Hi-C-seq combines DNA proximity ligation with high-throughput sequencing in a genome-wide fashion to identify interacting regulatory features such as enhancers and promotors. Variants that affect chromatin looping are called clQTLs. | The caQTL variant rs2692224 in the FN1 locus has also been described to alter chromatin looping in human coronary artery SMCs28 | |
| RNA | RNA level | eQTL, reporter assays | Variants that impact transcript level or gene expression (e) are called eQTLs. To detect eQTLs, genotyping and RNA sequencing are combined followed by association tests. Allele specific activity can be confirmed by reporter assay such as MPRA or STARR-seq.17 | About 85% of CAD loci contain eQTLs (see Supplementary material online, Table S1). The SMAD3 variant rs17293632 is one example in which the eQTL can be explained by the modulation of a TF binding site. However, eQTLs are often cell type and context-specific, i.e. only detectable after adding a certain stimulus. |
| UTR variants | mRNA stability/miRNA binding, etc. | UTR variants can have a variety of functional consequences such as altering the secondary structure, stability, location, translation, or binding of microRNAs or RNA-binding proteins. There are bioinformatics tools to predict mRNA stability or binding of microRNAs (e.g. https://mirdb.org/). | Two CAD-associated variants (rs9818870 and rs40593) are located in the 3′-UTR of the MRAS gene, adjacent to a cluster of miRNA binding sites, thereby most likely affecting mRNA stability and biosynthesis of the MRAS protein31,32 | |
| RNA editing | edQTL | Only a few disease-associated variants have been reported to directly affect RNA modifications or RNA editing, e.g. adenosine-to-inosine conversion. Although it has been shown that variants that affect RNA editing (so-called edQTLs) can contribute to the genetic risk of common inflammatory diseases,33 comprehensive datasets of edQTLs are missing. | Recently, 81 variants associated with CAD or acute MI were identified to affect RNA modifications.34 Four of these variants were genome-wide significant for CAD or acute MI (rs3739998 in JCAD, rs148172130 in RPL14, rs12190287 in TCF21, and rs186643756 in PVT1). | |
| RNA splicing | sQTL | Alternative splicing is critical in atherosclerosis.35 Like eQTL analyses, genotyping and isoform-sensitive RNA sequencing are required to detect splice (s)QTLs that can impact alternative splicing (e.g. exon skipping) and/or isoform usage. | The exon variant rs688 leads to alternative splicing of LDLR mRNA, rapid degradation of the LDLR isoform, and increased LDL cholesterol levels. Rare intronic LDLR variants were also identified to affect splicing in familial hypercholesterolemia patients.36,37 | |
| Circular RNA | circRNAs-seq | Circular (circ)RNAs, products of back-splicing of precursor mRNAs and covalent ligation of the 3′ and 5′ end, are abundantly expressed with high evolutionary conservation, show high tissue specificity, and exert their regulatory functions by miRNA sponging, protein sponging, or direct interaction with proteins of the transcriptional machinery.38 Detection methods for circRNAs can be found in these reviews.39,40 | CircRNAs have been linked to various cardiovascular diseases38,41 (e.g. circANRIL, circSATB2, circRUSC2, or circCHFR).38,42 The circular version of the antisense non-coding RNA at the INK4 locus (circANRIL) transcribed on the 9p21 locus is more abundant in carriers of the CAD-protective haplotype. The protective effect is thought to be due to circANRIL-mediated disturbed maturation of precursor rRNA, thereby inducing nucleolar stress and apoptosis in vascular SMCs and macrophages.16,43 | |
| Protein | Protein level | pQTL | Variants that impact protein level are called pQTLs, which are identified through association tests of genotypes and protein abundance in certain tissues or cell types. | A dose–response relationship has been described for lipoprotein(a), with protective CAD variants lowering lipoprotein(a) level, whereas higher levels of lipoprotein(a) were found in individuals carrying CAD-causal variants12 |
| Protein function | Enzyme activity | Missense variants that change the amino sequence of a protein can alter protein function in many ways. Therefore, enzyme specific functional assays must be carefully designed to confirm the impact of a missense variant. However, a variety of bioinformatics prediction tools are available (e.g. LoFtool44 or AlphaMissense.45 | At least 20 rare loss-of-function PCSK9 variants46 and 1 common missense variant in PCSK9 have been reported.7 The consequences of these variants can range from degradation of PCSK9, compromised intracellular transport, aberrant autocatalysis, or reduced affinity for the LDLR on the liver surface which is normally targeted through PCSK9 to lysosomal degradation after internalization of the LDLR-LDL complex. | |
| Protein modification | Mass spectrometry (MS), Western blot analysis (WB) | Missense variants that alter post-translational modifications have been reported frequently. MS is widely used to detect post-translational modification such as phosphorylation. WB can confirm such findings but is less sensitive. | A coding variant rs11556924-T (p.Arg363His) of ZC3HC1 (involved in cell cycle) results in enhanced phosphorylation at serin 395 of the ZC3HC1 protein, thereby reducing its activity.47 Of note, synonymous variants can have functional effects on mRNA stability and may contribute to co-translational misfolding or changes in co-translational modifications such as glycosylation.48 | |
| Interaction partner | Crosslinking mass spectrometry (clMS), co-immunoprecipitation (Co-IP), fluorescence resonance energy transfer (FRET), yeast two-hybrid screening (Y2H) | Missense variants can affect the binding affinity to binding partners with multiple consequences, depending on the nature of binding partners (e.g. proteinases or substrates). To detect protein–protein interactions, several methods are available such as high-throughput clMS49 as well as targeted approaches such as co-IP, FRET, Y2H, and others.50 | Due to its size, the ADAMTS7 protein has many interaction partners, such as activators (e.g. furin51,52), inhibitors (e.g. PRGN48 or A2M53) and substrates (e.g. TSP-1,54 TIMP1,55 SVEP1,56 or COMP57). The protective minor allele rs3825807-G (p.Ser214Pro) hampers furin-dependent cleavage of the pro-domain of ADAMTS7, resulting in lower level of active ADAMST7 |
| Level | Regulatory feature | Analysis | Description | Example of CAD-associated genes |
|---|---|---|---|---|
| Genome | Chromatin state | ATAC-seq or DNase-seq | Both sequencing techniques allow for the detection of accessible chromatin, a prerequisite for active regulation of gene expression. Variants that affect chromatin accessibility are called caQTLs. | The caQTL variant rs2692224 located downstream of the FN1 gene was identified using ATAC-seq in human coronary artery SMCs28 |
| Enhancer/TF binding sites | ChIP-seq, CRISPR-Cas perturbation | Chromatin immunoprecipitation (ChIP) is combined with DNA-seq. DNA and proteins (histones or TFs) are crosslinked. Fragmented protein-bound DNA is captured using antibodies and sequenced. Variants that alter TF binding are called bQTLs. CREs can be confirmed using CRISPR-Cas perturbation experiments.17 | By combining DNase-seq and histone ChIP-seq data, Turner et al. identified a protective T allele of rs17293632 that disrupts an activator protein-1 site in a SMAD3 intron enhancer, resulting in reduced SMAD3 expression.29 Toropainen et al. also identified this variant in ECs.30 | |
| Chromatin looping | Hi-C-seq | Hi-C-seq combines DNA proximity ligation with high-throughput sequencing in a genome-wide fashion to identify interacting regulatory features such as enhancers and promotors. Variants that affect chromatin looping are called clQTLs. | The caQTL variant rs2692224 in the FN1 locus has also been described to alter chromatin looping in human coronary artery SMCs28 | |
| RNA | RNA level | eQTL, reporter assays | Variants that impact transcript level or gene expression (e) are called eQTLs. To detect eQTLs, genotyping and RNA sequencing are combined followed by association tests. Allele specific activity can be confirmed by reporter assay such as MPRA or STARR-seq.17 | About 85% of CAD loci contain eQTLs (see Supplementary material online, Table S1). The SMAD3 variant rs17293632 is one example in which the eQTL can be explained by the modulation of a TF binding site. However, eQTLs are often cell type and context-specific, i.e. only detectable after adding a certain stimulus. |
| UTR variants | mRNA stability/miRNA binding, etc. | UTR variants can have a variety of functional consequences such as altering the secondary structure, stability, location, translation, or binding of microRNAs or RNA-binding proteins. There are bioinformatics tools to predict mRNA stability or binding of microRNAs (e.g. https://mirdb.org/). | Two CAD-associated variants (rs9818870 and rs40593) are located in the 3′-UTR of the MRAS gene, adjacent to a cluster of miRNA binding sites, thereby most likely affecting mRNA stability and biosynthesis of the MRAS protein31,32 | |
| RNA editing | edQTL | Only a few disease-associated variants have been reported to directly affect RNA modifications or RNA editing, e.g. adenosine-to-inosine conversion. Although it has been shown that variants that affect RNA editing (so-called edQTLs) can contribute to the genetic risk of common inflammatory diseases,33 comprehensive datasets of edQTLs are missing. | Recently, 81 variants associated with CAD or acute MI were identified to affect RNA modifications.34 Four of these variants were genome-wide significant for CAD or acute MI (rs3739998 in JCAD, rs148172130 in RPL14, rs12190287 in TCF21, and rs186643756 in PVT1). | |
| RNA splicing | sQTL | Alternative splicing is critical in atherosclerosis.35 Like eQTL analyses, genotyping and isoform-sensitive RNA sequencing are required to detect splice (s)QTLs that can impact alternative splicing (e.g. exon skipping) and/or isoform usage. | The exon variant rs688 leads to alternative splicing of LDLR mRNA, rapid degradation of the LDLR isoform, and increased LDL cholesterol levels. Rare intronic LDLR variants were also identified to affect splicing in familial hypercholesterolemia patients.36,37 | |
| Circular RNA | circRNAs-seq | Circular (circ)RNAs, products of back-splicing of precursor mRNAs and covalent ligation of the 3′ and 5′ end, are abundantly expressed with high evolutionary conservation, show high tissue specificity, and exert their regulatory functions by miRNA sponging, protein sponging, or direct interaction with proteins of the transcriptional machinery.38 Detection methods for circRNAs can be found in these reviews.39,40 | CircRNAs have been linked to various cardiovascular diseases38,41 (e.g. circANRIL, circSATB2, circRUSC2, or circCHFR).38,42 The circular version of the antisense non-coding RNA at the INK4 locus (circANRIL) transcribed on the 9p21 locus is more abundant in carriers of the CAD-protective haplotype. The protective effect is thought to be due to circANRIL-mediated disturbed maturation of precursor rRNA, thereby inducing nucleolar stress and apoptosis in vascular SMCs and macrophages.16,43 | |
| Protein | Protein level | pQTL | Variants that impact protein level are called pQTLs, which are identified through association tests of genotypes and protein abundance in certain tissues or cell types. | A dose–response relationship has been described for lipoprotein(a), with protective CAD variants lowering lipoprotein(a) level, whereas higher levels of lipoprotein(a) were found in individuals carrying CAD-causal variants12 |
| Protein function | Enzyme activity | Missense variants that change the amino sequence of a protein can alter protein function in many ways. Therefore, enzyme specific functional assays must be carefully designed to confirm the impact of a missense variant. However, a variety of bioinformatics prediction tools are available (e.g. LoFtool44 or AlphaMissense.45 | At least 20 rare loss-of-function PCSK9 variants46 and 1 common missense variant in PCSK9 have been reported.7 The consequences of these variants can range from degradation of PCSK9, compromised intracellular transport, aberrant autocatalysis, or reduced affinity for the LDLR on the liver surface which is normally targeted through PCSK9 to lysosomal degradation after internalization of the LDLR-LDL complex. | |
| Protein modification | Mass spectrometry (MS), Western blot analysis (WB) | Missense variants that alter post-translational modifications have been reported frequently. MS is widely used to detect post-translational modification such as phosphorylation. WB can confirm such findings but is less sensitive. | A coding variant rs11556924-T (p.Arg363His) of ZC3HC1 (involved in cell cycle) results in enhanced phosphorylation at serin 395 of the ZC3HC1 protein, thereby reducing its activity.47 Of note, synonymous variants can have functional effects on mRNA stability and may contribute to co-translational misfolding or changes in co-translational modifications such as glycosylation.48 | |
| Interaction partner | Crosslinking mass spectrometry (clMS), co-immunoprecipitation (Co-IP), fluorescence resonance energy transfer (FRET), yeast two-hybrid screening (Y2H) | Missense variants can affect the binding affinity to binding partners with multiple consequences, depending on the nature of binding partners (e.g. proteinases or substrates). To detect protein–protein interactions, several methods are available such as high-throughput clMS49 as well as targeted approaches such as co-IP, FRET, Y2H, and others.50 | Due to its size, the ADAMTS7 protein has many interaction partners, such as activators (e.g. furin51,52), inhibitors (e.g. PRGN48 or A2M53) and substrates (e.g. TSP-1,54 TIMP1,55 SVEP1,56 or COMP57). The protective minor allele rs3825807-G (p.Ser214Pro) hampers furin-dependent cleavage of the pro-domain of ADAMTS7, resulting in lower level of active ADAMST7 |
Abbreviations: ADAMTS7, a disintegrin and metalloproteinase with thrombospondin motifs 7; ATAC-seq, assay for transposase-accessible chromatin with high-throughput sequencing; A2M, alpha-2-macroglobulin; ChIP-seq, chromatin immunoprecipitation sequencing; CRISPR, clustered regularly interspaced short palindromic repeats; COMP, cartilage oligomeric matrix protein; DNase-seq, Dnase I hypersensitive sites sequencing; ECs, endothelial cells; FN1, fibronectin 1; LDLR, low density lipoprotein receptor; MPRA, massively parallel reporter assay; MRAS, muscle RAS oncogene homologue; PCSK9, proprotein convertase subtilisin/kexin type 9; PRGN, progranulin; QTL, quantitative trait loci; rRNA, ribosomal RNA; SMAD3, SMAD family member 3; SMCs, smooth muscle cells; STARR-seq, self-transcribing active regulatory region sequencing; SVEP1, Sushi, von Willebrand factor type A, EGF and pentraxin domain-containing protein 1; TIMP1, tissue inhibitor of metalloproteinases 1; TSP-1, thrombospondin-1; UTR, untranslated region.
2.2 Prioritizing causal variants and genes at CAD GWAS loci
In the last decade, the number of identified variants associated with CAD has increased dramatically due to larger GWAS; however, the effect sizes of these additional variants are rather small (i.e. odds ratio < 1.0527), making it even more challenging to confirm the causality of disease-driving variants and corresponding genes. If CAD variants are protein coding, linking causal genes is often straightforward. A total of 104 CAD risk loci with 164 protein-altering variants, representing sentinel variants or proxy variants (r2 ≥ 0.6), have been identified so far (see Supplementary material online, Table S1). However, about 95% of CAD variants are in non-coding regions that moderately affect gene expression.58 Many of the CAD loci are broad and contain multiple proximal genes as well as co-inherited variants in high LD. In addition, many variants show cell type specificity, and their regulatory potential are often only detectable in a context-specific manner, for example in response to a certain stimulus,30,59,60 making it even more difficult to determine which specific genetic factors are causally linked to CAD.
To tackle this problem, scientists have developed various approaches for prioritizing candidate genes and variants in GWAS loci.61–68 One approach is to use functional annotations to identify variants that are likely to have a regulatory influence on gene expression or protein function as they are located in gene promoters, enhancers, or coding regions. Successful examples in which the functional activity of regulatory risk variants has been confirmed in vitro can be found here7,28,69–71 (and above).
Functional annotations are often combined with statistical approaches, such as fine mapping19,21,72 and colocalization with molecular traits73–75 to underpin causality of variants and genes. Fine mapping identifies causal variants within a genomic region, while colocalization examines whether multiple traits or diseases share genetic variants. Multivariate colocalization includes fine mapping as part of its analysis. These statistical methods are affected by various parameters such as thresholds (e.g. P-value or false discovery rate) that are set for prior probabilities, and differences in haplotype structures of the populations from which GWAS data are collected. For instance, colocalization analysis is simple when the lead variants for GWAS and molecular quantitative trait loci (QTL) studies are the same or in high LD in both populations. While colocalization or fine-mapping analyses from a multi-ethnic population can identify common variants that have disease relevance for all ethnic groups, some ancestry-specific variants may still be missed due to differences in haplotype structures.
Another approach is to use gene-based tests that prioritize genes with a high burden of genetic variation, such as rare or common variants with large effect sizes. Multiple SNP-to-gene pipelines are now available that integrate a variety of datasets and perform machine learning to provide a probability score that displays the likelihood that a variant is regulatory active and affects a certain gene. Table 3 summarizes frequently used SNP-to-gene pipelines that provide variant scores via file transfer protocol (FTP) server, application programming interfaces (API) or web interfaces as well as Python/R tools.
Overview of exemplary SNP-to-gene pipelines
| Pipeline | Description | Available | References |
|---|---|---|---|
| Open Targets Genetics | The ‘locus-to-gene’ (L2G) model derives features to prioritize likely causal genes at each GWAS locus based on genetic and functional genomics features | FTP, API, web browser | 66 |
| RegulomeDB | A database that provides functional context to variants and serves as a tool to prioritize functionally important single nucleotide variants located within non-coding regions | FTP, web browser | 26 |
| FUMA GWAS | The SNP2GENE function takes GWAS summary statistics as an input and provides extensive functional annotation for all SNPs in genomic areas identified by lead SNPs | Web browser | 78 |
| Functional genome wide association analysis (FGWAS) | FGWAS consists of three components, a multivariate varying coefficient model, a global sure independence screening procedure, and a test procedure to discover important genetic variants and gene-environmental interactions | Python Package | 79 |
| MAGMA | A tool for gene analysis and generalized gene-set analysis of GWAS data. The gene analysis is based on a multiple regression model to provide better statistical performance. | Shell tool | 80 |
| DEPICT | An integrative tool that employs predicted gene functions to systematically prioritize the most likely causal genes at associated loci, highlight enriched pathways, and identify tissues/cell types where genes from associated loci are highly expressed | Python package | 81 |
| PoPs | Polygenic priority score (PoPs) is a gene prioritization method that leverages genome-wide signal from GWAS summary statistics and incorporates data from an extensive set of public bulk and single-cell expression datasets, curated biological pathways, and predicted protein–protein interactions | Python script | 82 |
| Effector index | Machine learning of commonly used genomic features, including open chromatin maps, eQTL, and chromatin conformation data to predict target genes at GWAS loci | R/shell tool | 83 |
| Summary-data-based Mendelian randomization (SMR) | SMR test for pleiotropic association between the expression level of a gene and a complex trait of interest using summary-level data from GWAS and eQTL studies | R script | 75 |
| ABC score | In combination with CRISPRi-FlowFISH the ABC model provide a systematic approach to map and predict which enhancers regulate which genes | FTP, web browser | 76 |
| GeneHancer | Genome-wide integration of enhancers and target genes in GeneCards | Web browser | 84 |
| DeepSEA | DeepSEA is a deep learning framework that predicts genomic variant effects on a wide range of regulatory features with single nucleotide sensitivity: TFs binding, DNase I hypersensitive sites, and histone marks in multiple human cell types | Web browser | 76 |
| Pipeline | Description | Available | References |
|---|---|---|---|
| Open Targets Genetics | The ‘locus-to-gene’ (L2G) model derives features to prioritize likely causal genes at each GWAS locus based on genetic and functional genomics features | FTP, API, web browser | 66 |
| RegulomeDB | A database that provides functional context to variants and serves as a tool to prioritize functionally important single nucleotide variants located within non-coding regions | FTP, web browser | 26 |
| FUMA GWAS | The SNP2GENE function takes GWAS summary statistics as an input and provides extensive functional annotation for all SNPs in genomic areas identified by lead SNPs | Web browser | 78 |
| Functional genome wide association analysis (FGWAS) | FGWAS consists of three components, a multivariate varying coefficient model, a global sure independence screening procedure, and a test procedure to discover important genetic variants and gene-environmental interactions | Python Package | 79 |
| MAGMA | A tool for gene analysis and generalized gene-set analysis of GWAS data. The gene analysis is based on a multiple regression model to provide better statistical performance. | Shell tool | 80 |
| DEPICT | An integrative tool that employs predicted gene functions to systematically prioritize the most likely causal genes at associated loci, highlight enriched pathways, and identify tissues/cell types where genes from associated loci are highly expressed | Python package | 81 |
| PoPs | Polygenic priority score (PoPs) is a gene prioritization method that leverages genome-wide signal from GWAS summary statistics and incorporates data from an extensive set of public bulk and single-cell expression datasets, curated biological pathways, and predicted protein–protein interactions | Python script | 82 |
| Effector index | Machine learning of commonly used genomic features, including open chromatin maps, eQTL, and chromatin conformation data to predict target genes at GWAS loci | R/shell tool | 83 |
| Summary-data-based Mendelian randomization (SMR) | SMR test for pleiotropic association between the expression level of a gene and a complex trait of interest using summary-level data from GWAS and eQTL studies | R script | 75 |
| ABC score | In combination with CRISPRi-FlowFISH the ABC model provide a systematic approach to map and predict which enhancers regulate which genes | FTP, web browser | 76 |
| GeneHancer | Genome-wide integration of enhancers and target genes in GeneCards | Web browser | 84 |
| DeepSEA | DeepSEA is a deep learning framework that predicts genomic variant effects on a wide range of regulatory features with single nucleotide sensitivity: TFs binding, DNase I hypersensitive sites, and histone marks in multiple human cell types | Web browser | 76 |
Overview of exemplary SNP-to-gene pipelines
| Pipeline | Description | Available | References |
|---|---|---|---|
| Open Targets Genetics | The ‘locus-to-gene’ (L2G) model derives features to prioritize likely causal genes at each GWAS locus based on genetic and functional genomics features | FTP, API, web browser | 66 |
| RegulomeDB | A database that provides functional context to variants and serves as a tool to prioritize functionally important single nucleotide variants located within non-coding regions | FTP, web browser | 26 |
| FUMA GWAS | The SNP2GENE function takes GWAS summary statistics as an input and provides extensive functional annotation for all SNPs in genomic areas identified by lead SNPs | Web browser | 78 |
| Functional genome wide association analysis (FGWAS) | FGWAS consists of three components, a multivariate varying coefficient model, a global sure independence screening procedure, and a test procedure to discover important genetic variants and gene-environmental interactions | Python Package | 79 |
| MAGMA | A tool for gene analysis and generalized gene-set analysis of GWAS data. The gene analysis is based on a multiple regression model to provide better statistical performance. | Shell tool | 80 |
| DEPICT | An integrative tool that employs predicted gene functions to systematically prioritize the most likely causal genes at associated loci, highlight enriched pathways, and identify tissues/cell types where genes from associated loci are highly expressed | Python package | 81 |
| PoPs | Polygenic priority score (PoPs) is a gene prioritization method that leverages genome-wide signal from GWAS summary statistics and incorporates data from an extensive set of public bulk and single-cell expression datasets, curated biological pathways, and predicted protein–protein interactions | Python script | 82 |
| Effector index | Machine learning of commonly used genomic features, including open chromatin maps, eQTL, and chromatin conformation data to predict target genes at GWAS loci | R/shell tool | 83 |
| Summary-data-based Mendelian randomization (SMR) | SMR test for pleiotropic association between the expression level of a gene and a complex trait of interest using summary-level data from GWAS and eQTL studies | R script | 75 |
| ABC score | In combination with CRISPRi-FlowFISH the ABC model provide a systematic approach to map and predict which enhancers regulate which genes | FTP, web browser | 76 |
| GeneHancer | Genome-wide integration of enhancers and target genes in GeneCards | Web browser | 84 |
| DeepSEA | DeepSEA is a deep learning framework that predicts genomic variant effects on a wide range of regulatory features with single nucleotide sensitivity: TFs binding, DNase I hypersensitive sites, and histone marks in multiple human cell types | Web browser | 76 |
| Pipeline | Description | Available | References |
|---|---|---|---|
| Open Targets Genetics | The ‘locus-to-gene’ (L2G) model derives features to prioritize likely causal genes at each GWAS locus based on genetic and functional genomics features | FTP, API, web browser | 66 |
| RegulomeDB | A database that provides functional context to variants and serves as a tool to prioritize functionally important single nucleotide variants located within non-coding regions | FTP, web browser | 26 |
| FUMA GWAS | The SNP2GENE function takes GWAS summary statistics as an input and provides extensive functional annotation for all SNPs in genomic areas identified by lead SNPs | Web browser | 78 |
| Functional genome wide association analysis (FGWAS) | FGWAS consists of three components, a multivariate varying coefficient model, a global sure independence screening procedure, and a test procedure to discover important genetic variants and gene-environmental interactions | Python Package | 79 |
| MAGMA | A tool for gene analysis and generalized gene-set analysis of GWAS data. The gene analysis is based on a multiple regression model to provide better statistical performance. | Shell tool | 80 |
| DEPICT | An integrative tool that employs predicted gene functions to systematically prioritize the most likely causal genes at associated loci, highlight enriched pathways, and identify tissues/cell types where genes from associated loci are highly expressed | Python package | 81 |
| PoPs | Polygenic priority score (PoPs) is a gene prioritization method that leverages genome-wide signal from GWAS summary statistics and incorporates data from an extensive set of public bulk and single-cell expression datasets, curated biological pathways, and predicted protein–protein interactions | Python script | 82 |
| Effector index | Machine learning of commonly used genomic features, including open chromatin maps, eQTL, and chromatin conformation data to predict target genes at GWAS loci | R/shell tool | 83 |
| Summary-data-based Mendelian randomization (SMR) | SMR test for pleiotropic association between the expression level of a gene and a complex trait of interest using summary-level data from GWAS and eQTL studies | R script | 75 |
| ABC score | In combination with CRISPRi-FlowFISH the ABC model provide a systematic approach to map and predict which enhancers regulate which genes | FTP, web browser | 76 |
| GeneHancer | Genome-wide integration of enhancers and target genes in GeneCards | Web browser | 84 |
| DeepSEA | DeepSEA is a deep learning framework that predicts genomic variant effects on a wide range of regulatory features with single nucleotide sensitivity: TFs binding, DNase I hypersensitive sites, and histone marks in multiple human cell types | Web browser | 76 |
In this review, we provide a comprehensive list of putative causal CAD risk genes with a cs for each gene of a CAD GWAS locus within a 1 Mb window. This score was created by adding the following weights: n = 2 if a gene has been previously assigned to be causal,7,10 n = 2 if a gene is linked to variant with an activity-by-contact (ABC) score above 0.15,76 n = 2 if a gene has a locus-to-gene score derived from the Open Targets pipeline above 0.4,66 n = 1 if a gene is a cis-eQTL-affected gene (eGene) (see Qtlizer database77), and n = 1 if a gene is a smooth muscle cell (SMC)-specific eGene (quiescent or proliferative state)62 (see Supplementary material online, Table S1, for more details).
As expected for atherosclerosis and reviewed here,11 many high-scored genes regulate lipid metabolism, including APOE, LDLR, lipoprotein(a) (LPA), LPL, PCSK9, and SCARB1 (Figure 3A). Causative gene candidates with high confidence were identified in approximately 30% and risk gene candidates in 57% of all the GWAS CAD loci (Figure 3B). Of the 244 autosomal CAD susceptibility loci, only 30 loci have no reliable data supporting the association between the variant and its corresponding genes, whereas at least 15 CAD loci harbour two or more causative genes (Figure 3C and D).
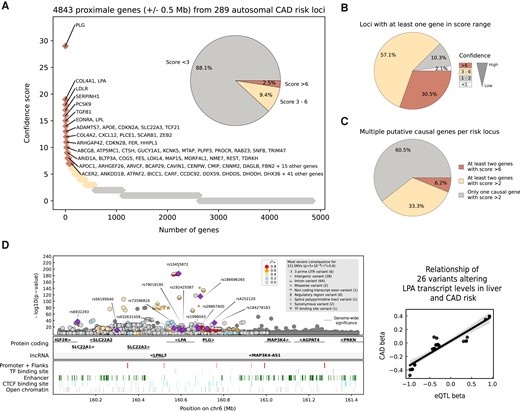
Characterization of CAD risk loci. (A) The cs for each CAD risk gene proximal to a CAD index variant (±0.5 Mb) was calculated using following weights: causal in recent SNP-to-gene analyses = 27,10; V2G score from Opentargets Genetics = 2; ABC score = 285; eQTLs from Qtlizer database = 177; SMC-specific eQTLs = 1.62 A score above 10 is possible if two or more independent CAD associations were detected for a given gene. (B) The causative gene has been identified with very high confidence for around one-third of CAD risk loci. (C) Approximately 40% of susceptibility loci may harbour more than one causative gene. (D) The regional association of the LPA/PLG locus demonstrates the complexity of a GWAS signal. Independent index variants from different CAD GWASs are shown in purple. Genetic features and most severe consequences are from Ensembl (release 110). eQTL affecting the expression level of LPA in liver correlate with CAD risk. The eQTL effect sizes (beta values) are from the Qtlizer database77 and CAD GWAS summary statistics are from Ref. 7.
The LPA locus illustrates the complexity of a GWAS signal as it contains multiple independent CAD risk variants (r2 < 0.4) that modulate more than two most likely causal genes. The variant-to-gene relationships for LPA, PLG, SLC22A1, and SLC22A3 are supported by several metrices, including polygenic priority score,7,82 Open Targets Genetic locus-to-gene score, and expression quantitative trait loci (eQTL) data (see Supplementary material online, Table S1). In addition, several independent variants in this locus are associated with LPA level (see GWAS Catalog and12 for more details). Moreover, it is well established that LPA and PLG are atherogenic factors due to their involvement in the fibrinolytic system and foam cell formation,86,87 which is also reviewed here.88
2.3 Identifying the tissues and cell types relevant to CAD
SNP- or gene-based tissue-specific enrichment analyses have proven useful to identify the cell types and tissues most relevant to CAD (Figure 4). For example, two groups have independently found that CAD SNPs are predominantly enriched in open chromatin regions of liver, adipose tissue, and blood vessels/artery tissue based on Dnase I hypersensitive sites6 or ChromHMM-derived chromatin states.7 In concordance, cis-eQTL enrichment revealed that a high proportion of CAD risk variants regulate transcriptional activity in arterial tissue.7 Zheng et al. used eight different computational methods to predict CAD potential causal genes and found that these genes were significantly overexpressed in adipose and liver tissues.89 These findings were confirmed by a large-scale genome-wide multi-ethnic association study90 using gene-property analyses (MAGMA and DEPICT) and others studies demonstrated that metabolic tissues (e.g. liver/adipose tissue) and vascular tissues (e.g. atherosclerotic arterial wall) are interconnected via cross-tissue regulatory gene networks.91–93
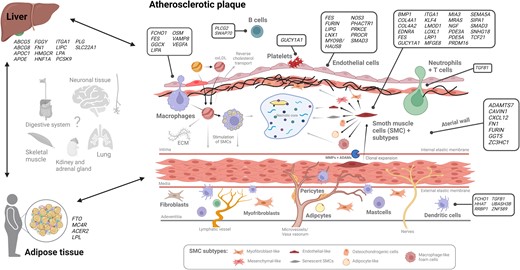
Relevant tissues and cell types in atherosclerosis. Liver and adipose tissue, which are critical for lipid metabolism, appear to be the most important atherogenic non-arterial tissues. Variants that are active in other tissue types also contribute to atherosclerosis, but the mechanisms are less well described. In the vessel wall, many lesional cell types can promote atherosclerosis, including dysfunctional ECs and infiltrated macrophages that uptake oxidized LDL particles (oxLDL). These oxLDL particles can also accumulate in the vessel wall by interacting with the extracellular matrix (ECM). The inflammatory environment triggers SMCs to remodel the arterial ECM through matrix metalloproteinases (MMPs) and ADAM family members (e.g. ADAMTS7), and to proliferate and migrate from the tunica media into the tunica intima. SMCs can adopt several subtypes (e.g. myofibroblast-like cells) in response to different stimuli (e.g. growth factors). CAD risk genes (cs >5) were allocated based on epigenetic enrichment of associated variants, cell-specific eQTLs, or tissue/cell type enriched gene expression (see Supplementary material online, Table S3). This figure was created with BioRender.com.
Lipid-associated CAD risk factors such as high low density lipoprotein (LDL) cholesterol can be readily treated using statins or PCSK9 modulation. However, many CAD risk loci have predicted roles in the arterial wall (e.g. ADAMTS7, GUCY1A1, NOS3, or MRAS)94,95 which has therefore become novel therapeutic target. The involvement of many cell types in atherosclerotic lesions, such as macrophages (MP), monocytes, lymphocytes, endothelial cells (ECs), SMCs, mast cells, and neutrophils, highlights the difficulty in identifying cell-specific genetic determinants of CAD risk.96,97 ECs and SMCs may be the most important, as single-cell epigenomics of five major lesional cell types (EC, SMC, MP, T cells, and B cells) revealed that CAD-associated genetic variants are enriched in EC- and SMC-specific open chromatin.65 In line with that, Tcheandjieu et al. highlighted the relevance of ECs, SMCs, and stromal–fibroblasts in the pathogenesis of CAD using MAGMA gene-property analyses of single-cell RNA datasets from mice and humans.90
However, the situation in atherosclerotic lesions is even more complex in terms of cell identity. Recent lineage tracing mouse models and single-cell RNA sequencing data98,99 demonstrated that lesional cell types are not homogenous, and there is considerable phenotypic evolution and cellular diversity within these cell types in atherosclerotic lesions. For example, up to seven distinct SMC clusters with atheroprotective or atherogenic properties are now known, which can adopt myofibroblast-, osteogenic-, or macrophage-like phenotypes.100,101 Initial efforts in this direction are made by some studies,102,103 but more are required to determine which cell types and subtypes are the exact drivers of the disease in which the CAD GWAS loci act.
2.4 Pathophysiological pathways in CAD
The number of common CAD risk alleles a person carries is in hundreds (∼190 sentinel risk variants in the UK Biobank population) without increasing the CAD risk dramatically.104 It is assumed that changes in transcriptional activity of hundreds of genes can be safely tolerated. The addition of just a few extra variants (e.g. 10–20 risk variants), however, results in sharp risk increments, as their effects multiply with each other and with modifiable risk factors such as smoking, ultimately leading to the destabilization of gene regulatory networks and biological pathways across various tissues.104 Researchers have therefore begun to investigate CAD risk genes within a systems biology framework. By leveraging Gene Ontology (GO) annotations for 517 identified causal gene candidates, originating from 299 CAD loci and surpassing a cs threshold of two (refer to Table 1 and Figure 3), we developed a comprehensive map of CAD-associated pathophysiological pathways (Figure 5). In addition to previous articles that reviewed the genetics and biological signalling pathways of CAD in the light of GWAS,10,11,13,15,17,27 this present review extends the list of biological processes that may also contribute to the progression of CAD, including neuronal activity or autophagy. For example, there is evidence that autophagy is closely associated with the progression of atherosclerosis and that enhancing autophagosome activity by inhibiting the mTOR signalling pathway with rapamycin105 or intermittent fasting106 can provide therapeutic benefits to patients at risk.107,108
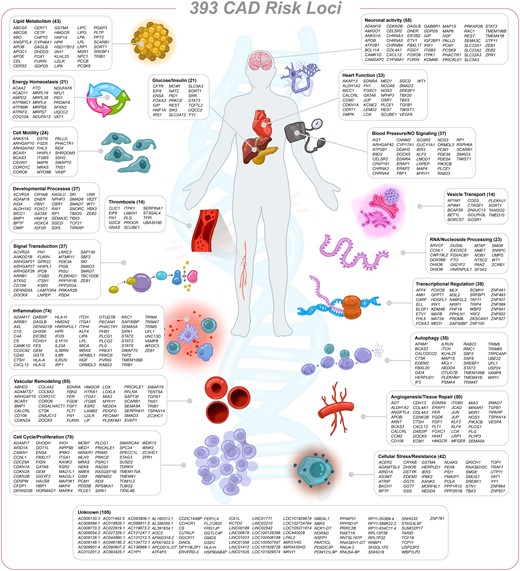
CAD-causal genes mapped to pathophysiological pathways in atherosclerosis. Genes with a cs >2 were assigned to biological processes using the GO annotations and assignments by a recent review.27 The number of genes is displayed in brackets. For simplification, some GO terms have been combined into one category, e.g. angiogenesis, wound healing, and tissue repair to angiogenesis/tissue repair or ECM organization was assigned to vascular remodelling. Of note, many genes can be involved in multiple biological processes. For example, the ubiquitously expressed endoprotease furin is assigned to lipid metabolism, neuronal activity, vascular remodelling, and signal transduction.
2.5 Clinical implications of CAD GWAS findings
2.5.1 Polygenic risk scores for risk prediction
Polygenic risk scores (PRS) use genetic information to predict an individual's risk of developing CAD.109 GWAS allows the comparison of SNPs between individuals with CAD and those without CAD to generate SNP-level association statistics, which are used to compute the PRS. The PRS is trained and validated in an independent cohort of individuals before its application in downstream research and clinical analysis.109–111
CAD PRS can be used for genetic risk prediction and stratification to guide clinical decision making. Individuals in the highest decile of a PRS have almost four-fold higher likelihood of prevalent CAD,7,112 which is similar to that posed by the presence of monogenic mutations for familial hypercholesterolemia. A high PRS in patients presenting with early-onset as well as late-onset MI112 is also independently associated with a higher risk of all-cause mortality.113 Studies have shown that PRS for CAD is close to the risk discrimination for individual clinical risk factors.114 By combining genetic information with classic risk factors, such as sex, age, and lifestyle factors, clinicians can identify individuals at increased CAD risk and tailor prevention strategies accordingly. High PRS may rationalize the selective use of cardiac imaging in asymptomatic individuals for screening and optimizing the timing of preventive therapy. Coronary artery calcium (CAC) score has shown to be strong predictor of intermediate-term CAD risk.115 A high PRS for CAD has been shown to be associated with CAC in younger adults as well as those with traditional risk factors,116 indicating that exposure to CAD risk alleles predisposes to subclinical and clinical atherosclerosis.116
Calculated early in life, PRS for CAD has the potential to provide a strong platform for primary prevention117 and justify the early use of lipid-lowering therapy. PRS determination can help set a baseline lifetime risk trajectory,118 even before the onset of risk factors, that can be utilized to estimate absolute long-term risk.119 In patients already on lipid-lowering therapy, those with high PRS for CAD derived the largest relative and absolute clinical benefit from statin therapy,116,120 which can be enhanced with a PCSK9 monoclonal antibody therapy.121,122 It has also been shown that the knowledge of a high PRS for CAD may enhance motivation to initiate or adhere to risk-reducing intervention.123
There are, however, potential limitations and challenges associated with using PRS, including the lack of approved guidelines, leading to inconsistencies among studies and misinterpretation of results.
Only recently, a protocol for PRS analysis with detailed guidelines has been published to discuss standard quality control and best practices.110
As the predominant focus of the GWAS has been on individuals of European ancestry, the PRS predictions do not work well in non-Europeans. Improving GWAS methods for accommodating diverse populations and using multi-ancestry statistics for constructing the PRS appears to be the way forward.
Lastly, challenges also exist related to ethical considerations such as patient autonomy and privacy. There is potential risk of genetic information and PRS being used against patients in areas such as insurance claims and determining healthcare insurance premium costs.109
2.5.2 Sex matters in CAD
Although CAD is ubiquitous in our society, important sex-related changes in its epidemiology have only recently been evaluated.124–127 CAD affects both men and women and is influenced by a combination of genetic, hormonal, and environmental factors, which result in a unique risk profile and symptom presentation for each sex. The molecular mechanisms that underlie these influences, however, have not been thoroughly investigated and are not well understood.
Many studies on the genetics of CAD are, however, limited by the lack of inclusion of sex chromosomes in the GWAS.128
The X and Y chromosomes contain protein coding genes that govern numerous biological mechanisms. Considering these genes becomes essential when exploring sex differences.
The uneven expression of X-linked genes between women (XX) and men (XY) requires the inactivation of one X chromosome, a process that is tissue- and cell-specific. Approximately 15% of X-linked genes, however, evade this inactivation, leading to a doubling of the gene dosage for these specific genes.129,130 These analytical challenges, especially by the X chromosome, require special computational tools and were excluded in most GWAS to simplify analysis. Out of the 53 studies with available dbGAP genotype data, only 31 included analysis of the X chromosome.128 Notably, none of these included GWAS for CAD until recently by Tcheandjieu et al.90
Research studies on the genetics of CAD were also limited by the inadequate consideration of sex as a variable in targeted genetic analyses and insufficient examination of hormone-gene interactions. Women are generally underrepresented in the preclinical, clinical cardiovascular research131–134 and in CAD GWAS.135,136 Women GWAS suffers from weaker statistical power and smaller effect sizes.7 There is also an element of misclassification of women in CAD due to the difference in the presentation of CAD in men and women. In contrast to men, who typically exhibit obstructive CAD, women often manifest non-obstructive CAD. This difference in presentation can lead to misclassification, particularly in studies that use obstructive CAD as an outcome, as women may be incorrectly categorized as controls.137,138
Recently, a GWAS meta-analysis was conducted using 181 522 cases of CAD (32% women vs. 68% men) and 1 165 690 controls (53% women vs. 47% men).7
A sex stratified GWAS involving a smaller subset of studies, with 77 080 cases (30% women vs. 70% men) and 550 952 controls (56% women vs. 44% men), identified 10 variants out of 241 conditionally independent associations with CAD that exhibited a differential effect between sexes.
Among the variants examined, only rs7696877 (MYOZ2) had a more pronounced effect in women than in men. Nine variants, including rs9521672 (COL4A1), rs4977574 (CDKN2B-AS1), rs12740374 (CELSR2), rs11225975 (MIR4693), rs112422902 (PVRL2), rs149722146 (EDNRA), rs186399184 (ICA1L), rs28451064 (NCRNA00310), and rs190352900 (RAPH1), had a greater impact on men. In the same line, Tcheandjieu et al. used 243 392 cases and 849 686 controls (<10% women) and identified 9 loci associated with CAD that are located on chromosome X (see Table 1). With the overrepresentation of men in this GWAS, this study signifies X-chromosomal contribution to CAD.90
Dungan et al. performed a sex stratified GWAS analysis on incident mortality in patients with CAD (25.4% women vs. 74.6% men).139 This study revealed eight variants that were shown to confer increased mortality risk in men: rs2076780 (GREM2/RGS7), rs11252040 (LOC105376360), rs17103766 (BRMS1L/LINC00609), rs2062640 (UNC13C/LOC105370829), rs2835913 (KCNJ6), rs4776247 (UNC13C/LOC105370829), rs12150051 (LINC00670), and rs9932462 (EMP2/TEKT5); while 15 variants showed sex-specific risk in women: rs8133010 (PDE9A), rs12145981 (LPGAT1), rs10494195 (LOC101929099), rs12145981 (LOC91548/LPGAT1), rs17591646 (SLC9A9), rs8021816 (PRKD1), rs26445 (LOC102546299/LINC01947), rs7217169 (RAP1GAP2), rs9388813 (TMEM200A/SMLR1), rs10923243 (VTCN1/LINC01525), rs1751291 (LINC00703/MANCR), rs10768256 (C11orf74/LINC02760), rs7320901 (LINC00457/NBEA), rs17051660 (LINC00457/NBEA), and rs9599764 (LINC00457/NBEA).
Although the genetic variants identified through sex-specific analysis show involvement in endothelial dysfunction, lipid metabolism, vascular remodelling, and thrombosis, the genetic underpinning of these sex differences in CAD is not completely understood. Large sex stratified GWAS, with appropriate study design, adequate sample size, and equal participation of men and women, can be critical in translating these genetic signals into specific disease pathways.
2.5.3 GWAS need more ethnic diversity
Despite the success of GWAS in identifying genetic variants associated with CAD, the majority of these studies from the CARDIoGRAMplusC4D 1M+ Hearts project (https://www.phpc.cam.ac.uk/ceu/1m-hearts-study/), UK Biobank,140 and Million Veteran Program (MVP)8 have primarily focused on people of European descent.
In 2007, it was estimated that the GWAS samples for CAD lacked non-European ancestry. By 2022, the percentage had, however, increased to 29%, with most of the increase attributed to people of East Asian origin rather than people from Africa, South Asia, Latin America, South America, the Middle East, and the Pacific Islands.14 One of the advantages of CAD GWAS for non-Europeans is that if certain genetic variants are uncommon or absent, it may reveal significant effects on the disease that may not be apparent in European studies.141,142 However, it is important to note that the associations between genotype and phenotype discovered in Europeans may not be equally relevant or significant in non-European populations.14 In particular, the accuracy of PRS relies on the genetic background of the population.143,144
Thanks to genotyping technologies, analytic methods, and more attention to underrepresented populations, by 2012, we started to see GWAS for CAD in other ethnic groups, such as Han Chinese145 and Japanese.146 In 2020, another CAD GWAS was conducted on the Japanese population, which revealed eight novel loci.9
To overcome the limitation of ethnicity, Tcheandjieu et al. conducted the first, large-scale, multi-population GWAS using data from White, Black, and Hispanic cohorts from the MVP. This analysis identified 95 novel loci and detected eight loci of genome-wide significance exclusively in Black or Hispanic individuals.90 Interestingly, this multi-population GWAS demonstrated that two common haplotypes at the 9p21 locus contributed to risk stratification across all populations except those of African origin, where these haplotypes are virtually absent. This observation even after a two-stage meta-analysis involves >27 000 and >12 100 CAD cases.90 In addition, Lettre et al. reported that the index SNP rs6475606 in 9p21 was also insignificant (P = 6.4 × 10−4) in a meta-analysis of multiple African American cohorts.147
Conducting GWAS on diverse ethnicities can address health disparities disproportionately affecting certain ethnic groups. For example, African Americans and Hispanics are at a higher risk of developing CAD than other ethnic groups.148,149 We can identify potential genetic factors contributing to this increased risk, which may ultimately lead to the development of more effective, personalized, and targeted interventions.
2.5.4 GWAS guided drug development
Since preclinical compounds are twice as likely to succeed if there is genetic evidence, findings from GWAS can inform drug discovery.150 GWAS have proven useful in identifying disease-associated genes and pathways to guide drug development10–12 (Figure 6A).
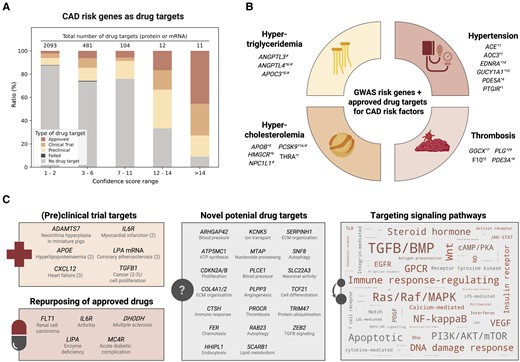
CAD risk genes as potential therapeutic target. (A) Classification of risk genes as potential drug targets for CAD derived from the therapeutic target database.151 The higher the cs, which represents the level of evidence of causality, the more likely it is to identify a drug target that interacts with drugs approved by the U.S. Food and Drug Administration (FDA). (B) Approved targets encoded by genes that impact CAD risk and CAD-associated pathomechanisms such as hypercholesterolemia. cs are shown in superscript (*). Some drug targets were also identified in large-scale array-based and deep-sequencing projects (#).10 (C) Exemplary drug targets that can be (re)used for new therapeutic strategies. Some drug targets encoded by CAD risk gene (cs ≥ 5) are already tested in animal models or clinical trials (phase in parentheses) for CAD-related indications, others interact with FDA-approved drugs prescribed today for other diseases (shown in grey), or were not yet recognized as clinical drug targets (middle panel, cs ≥ 10). According to the Drug-Gene Interaction Database (https://www.dgidb.org/), almost all these genes are druggable or clinically actionable (except RAB23, SNF8, and TCF21). A fourth strategy may involve targeting multiple components of CAD-relevant signalling pathways, e.g. through antisense RNAs. The larger the font size, the more CAD risk candidate genes are part of the signalling pathway. The word cloud was created from GO annotations of CAD risk genes (cs ≥ 3) using the python package wordcloud (pypi.org/project/wordcloud/). Details of drug targets can be found in Supplementary material online, Tables S4 and S5.
Lowering the blood LDL cholesterol level is considered an effective method for primary prevention of arteriosclerosis.152 Unsurprisingly, the use of statins, a class of drugs also known as 3-hydroxy-3-methylglutaryl-CoA reductase inhibitors, increased by ∼25% worldwide from 2015 to 2020.153 Inspired by the LDL-lowering nature of loss-of-function variants of PCSK9,46 inhibition of PCSK9 became a promising therapeutic strategy for aggressive LDL-lowering in high-risk patients with atherosclerotic cardiovascular disease using antibodies, vaccines, siRNAs, or gene editing,121,122 which is associated with a safety profile superior to that of other classes of LDL-lowering drugs.46 Moreover, gain-of-function mutations in PCSK9, which cause familial hypercholesterolemia and CAD,154–157 serve as one of the best examples of how genetic insights can be used as a powerful tool to develop new drug targets. Other examples of therapeutic targets (e.g. PDE5A for hypertension) that deep-sequencing and/or GWAS have identified are given in Figure 6B. Guided by GWAS outcomes, other therapeutic targets could also be clinically relevant in the near future, including inflammation152 or intimal hyperplasia.158
Progress is being made in the treatment of in-stent restenosis, which involves migration and proliferation of vascular SMCs. Several drug targets genetically associated with CAD are serving as targets in clinical trials [e.g. transforming growth factor B1 (TGFB1)/TGFB1 receptor159] or are about to enter clinical trials (e.g. ADAMTS7160) (Figure 6C and Supplementary material online, Table S4). Since lower ADAMST7 activity has been linked to less SMC-mediated neointima formation, lower burden of vessel calcification, and more stable plaques,52,55 inhibiting ADAMTS7 is considered a novel non-lipid therapy against cardiovascular diseases. GWAS findings can also inform repurposing of approved drugs. One example is IL6R which is causally linked to CAD and other common vascular diseases.13 The human monoclonal antibody tocilizumab (DrugBank ID: DB06273), which targets the IL6R and is approved for treatment of rheumatoid arthritis, has been investigated in a clinical trial in patients with MI (ClinicalTrials.gov: NCT01491074, phase II). Another IL6 antagonist, ziltivekimab, has been evaluated in patients at high atherosclerotic risk with moderate to severe chronic kidney disease and high-sensitivity C-reactive protein (CRP) of at least 2 mg/L (ClinicalTrials.gov: NCT03926117, phase II). The subcutaneous administration of ziltivekimab markedly reduced high-sensitivity CRP by up to ∼90% and led to the reduction of other pro-inflammatory biomarkers in dose-dependent manner.161 Finally, recent CAD GWAS meta-analyses revealed most likely causal genes that are not currently recognized as drug targets, but are part of signalling pathways, modulation of which may be beneficial in reducing CAD risk, e.g. TGFB signalling pathway (Figure 6C).
3. Future directions and expectations
Hundreds of genetic variants associated with CAD have been identified through GWAS3,6–9,90,146 but distinguishing between causative genetic variants and those merely associated with a disease is a major challenge. Advances in statistical genetics and causal inference methods are expected to improve our ability to identify the causal genetic variants that contribute to CAD. Integrating data from multiple omics technologies, including epigenomics, transcriptomics, proteomics, metabolomics, and the microbiome, can provide a more comprehensive understanding of the biological mechanisms underlying genetic associations with CAD. Integration of large-scale clinical data, such as electronic health records, can provide insights into the clinical features and outcomes of CAD, which can help refine our understanding of the genetic basis of the disease and improve treatment strategies.
In summary, CAD GWAS have provided important insights into the genetic architecture of CAD and will likely bring many exciting developments that can lead to more personalized and effective therapies for the disease and improve our ability to prevent, diagnose, and treat CAD.
Supplementary material
Supplementary material is available at Cardiovascular Research online. The data underlying this article are available in the article and/or can be downloaded from the sources listed in Table 3.
Acknowledgement
This work is dedicated to the memory of Jeanette Erdmann, who passed away on 9 July 2023, before the publication of this paper. She was a remarkable researcher and a compassionate mentor who had a significant impact on the research field. Her dedication, professionalism, and distinctive approach will always be remembered and treasured.
Funding
R.A. was supported by an Postdoctoral Fellowship by American Heart Association 18POST33990046, the University of Eastern Finland (Researcher Fellowship), the Finnish Foundation for Cardiovascular Research, German Centre for Cardiovascular Research (DZHK, FKZ81Z0700108, FKZ81X2700133), the Junior Investigator Award from Leducq Foundation (18CVD02, PlaqOmics, R.A., and T.R.), the Junior Research Cardiovascular Diseases Grant of the CORONA Foundation (Corona-Stiftung) (S0199/10097/2023 to R.A.), and German Research Foundation (DFG) (528455242 and 515637292 to R.A.).
Data availability
All data are incorporated into the article and its online supplementary material.
References
Author notes
Conflict of interest: None declared.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}