





Abstract
Pelecanimimus polyodon was discovered in 1993 in the Spanish Barremian fossil site of Las Hoyas, being the first ornithomimosaur described from Europe. So far, there has been no detailed description of the holotype of Pelecanimimus, which is composed of the anterior-half of an articulated skeleton. Here we report a new, detailed, revised and more accurate osteological description of its postcranial skeleton, comparing this new data to information about Ornithomimosauria from the last three decades. This osteological and phylogenetic analysis of Pelecanimimus shows several ornithomimosaur synapomorphies and a unique combination of characters that emend its original diagnosis. Pelecanimimus diverged early in Ornithomimosauria and reveals an enlargement trend of the manus, shared with derived ornithomimosaurians, due to a long metacarpal I and elongated distal phalanges. This evolutionary novelty, and other synapomorphies, has led to the definition of a new clade, Macrocheiriformes, including Pelecanimimus and more derived ornithomimosaurs. Pelecanimimus has the only ossified sternal plates among ornithomimosaurs and the first evidence of uncinate processes in a non-maniraptoran theropod, indicating a convergent appearance of these structures in Coelurosauria. The character combination in an early-diverging ornithomimosaur like Pelecanimimus found in this analysis provides a key step in the evolution of the manus and pectoral girdle in Ornithomimosauria.
INTRODUCTION
Ornithomimosauria was defined by Sereno (1998) as the stem-based taxon that includes all maniraptoriforms closer to Ornithomimus Marsh, 1890 than to Neornithes. In recent phylogenies, Ornithomimosauria is one of the most basal clades in Coelurosauria, more derived than Tyrannosauroidea, and the sister-group to Maniraptora (e.g. Choiniere et al., 2010, 2012; Lee et al., 2014; Chinzorig et al., 2018).
Ornithomimosaurs are widely distributed throughout the world, especially abundant in Cretaceous beds of Asia and North America (Makovicky et al., 2004). However, the European record is poor, with ornithomimosaur fossils recovered only from England, France and Spain. The English record includes fragmentary Early Cretaceous material described as ValdoraptorOlshevsky, 1991, now considered a nomen dubium (Allain et al., 2014). In south-western France, the Berriasian Angeac-Charente bonebed (Rozada et al., 2018) has yielded several bones recognized as ornithomimosaurs, preliminarily described by Allain et al. (2014) and still under study. The first known European ornithomimosaur was Pelecanimimus polyodonPérez-Moreno et al. 1994 from the Las Hoyas fossil site, a Konservat-lagerstätte from the Barremian (Early Cretaceous) of Cuenca (Spain).
Pelecanimimus polyodon was discovered in 1993, being the first non-avian dinosaur found from the site (Pérez-Moreno et al., 1994). Unitil now, there has not been a published detailed description of the holotype of PelecanimimusPérez-Moreno et al., 1994, except for an unpublished doctoral thesis (Pérez-Moreno, 2004). The holotype (and only known specimen) is the anterior-half of an articulated skeleton (MCCM-LH 7777). The postcranial skeleton of Pelecanimimus preserves all cervical vertebrae, including the proatlas, atlas and axis, two anteriormost dorsal vertebrae and three posterior dorsal vertebrae, ribs, sternal plates, almost complete pectoral girdle and both almost complete forelimbs (Fig. 1). During the last 27 years since its discovery, the known diversity of ornithomimosaurs has increased with many new discoveries made worldwide (e.g. Klerk et al., 2000; Makovicky et al., 2010; Chinzorig et al., 2018). New detailed descriptions of historic material have also been provided (e.g. Kobayashi & Barsbold, 2005a, b; Lee et al., 2014; Claessens & Loewen, 2016). Among the vast ornithomimosaurian record, the axial and appendicular skeletons are altogether well represented. Taxa such as Archaeornithomimus Russel, 1972 (Gilmore, 1933; Smith & Galton, 1990), DeinocheirusOsmólska & Roniewicz, 1970 (Lee et al., 2014), Gallimimus bullatusOsmόlska et al., 1972, HarpymimusBarsbold & Perle, 1984 (Kobayashi & Barsbold, 2005a), Ornithomimus edmontonicusSternberg, 1933, SinornithomimusKobayashi & Lü, 2003 or StruthiomimusLambe, 1902 (Nicholls & Russell, 1981, 1985) have an almost complete and well-preserved axial skeleton, pectoral girdles and forelimbs. On the other hand, the atlas–axis complex is poorly represented in the fossil record, as only Deinocheirus (Lee et al., 2014), Gallimimus bullatusOsmólska et al., 1972, GarudimimusBarsbold, 1981 (Kobayashi & Barsbold, 2005b), Sinornithomimus (Kobayashi & Lü, 2003) and Struthiomimus (Lambe, 1902) preserve them complete or partially complete. Regarding other ornithomimosaurs, BeishanlongMakovicky et al., 2010, despite lacking most cervical and all dorsal vertebrae, has a well-preserved and almost complete pectoral girdle and forelimb. Although the holotype of ShenzhousaurusJi et al., 2003 is an almost complete skeleton, the forelimb and cervical region are practically missing (Ji et al., 2003).

Postcranial skeleton of Pelecanimimuspolyodon (MCCCM-LH 7777). A, right side. B, left side. Scale bars: 50 mm.
Pérez-Moreno et al. (1994) described Pelecanimimus polyodon (MCCM-LH 7777) as an ornithomimosaur with the following two diagnostic features in the postcranial skeleton: (1) ulna and radius are tightly adhered distally and (2) a metacarpal ratio of 0.81/1/0.98. Here, we carry out the first detailed osteological description of the postcranial skeleton of the ornithomimosaur Pelecanimimus polyodon, highlighting the features relevant to its phylogenetic affinities, as well as the evolutionary history of other ornithomimosaurs. The postcranial elements of this specimen are compared with all taxa mentioned above, especially with those that have been personally studied. The original diagnostic features proposed by Pérez-Moreno et al. (1994) are reviewed and discussed, and a new revised diagnosis, based on the postcranial elements of Pelecanimimus, is proposed, enabling a better understanding of the early evolution of the postcranial skeleton of ornithomimosaurs.
MATERIAL AND METHODS
Measurements and comparison
Bones were measured first-hand on the examined specimen. All measurements (in mm) are given in Tables 1–4.
Measurements of pectoral girdle elements of Pelecanimimus MCCM-LH 7777 (in mm). Abbreviations: dp, depth; tw, transversal width; *incomplete bones; **sum of measurements of all the fragments of the bone
| Right Scapula | Left Scapula | Right Coracoid | Left Coracoid | |
|---|---|---|---|---|
| Length | 130* | 103.5** | 65 | 50 |
| tw | - | - | 10 | - |
| Proximal dp | 15 | 21 | - | 60 |
| Midshaft dp | 17 | 17 | - | - |
| Distal dp | 24 | 12,5* | - | 20 |
| Right Scapula | Left Scapula | Right Coracoid | Left Coracoid | |
|---|---|---|---|---|
| Length | 130* | 103.5** | 65 | 50 |
| tw | - | - | 10 | - |
| Proximal dp | 15 | 21 | - | 60 |
| Midshaft dp | 17 | 17 | - | - |
| Distal dp | 24 | 12,5* | - | 20 |
Measurements of pectoral girdle elements of Pelecanimimus MCCM-LH 7777 (in mm). Abbreviations: dp, depth; tw, transversal width; *incomplete bones; **sum of measurements of all the fragments of the bone
| Right Scapula | Left Scapula | Right Coracoid | Left Coracoid | |
|---|---|---|---|---|
| Length | 130* | 103.5** | 65 | 50 |
| tw | - | - | 10 | - |
| Proximal dp | 15 | 21 | - | 60 |
| Midshaft dp | 17 | 17 | - | - |
| Distal dp | 24 | 12,5* | - | 20 |
| Right Scapula | Left Scapula | Right Coracoid | Left Coracoid | |
|---|---|---|---|---|
| Length | 130* | 103.5** | 65 | 50 |
| tw | - | - | 10 | - |
| Proximal dp | 15 | 21 | - | 60 |
| Midshaft dp | 17 | 17 | - | - |
| Distal dp | 24 | 12,5* | - | 20 |
Measurements of forelimb elements of Pelecanimimus MCCM-LH 7777 (in mm). Abbreviations: dp, depth; tw, transversal width; *incomplete bones; **sum of measurements of all the fragments of the bone
| Right Humerus | Left Humerus | Right Ulna | Left Ulna | Right Radius | Left Radius | |
|---|---|---|---|---|---|---|
| Length | 180** | 192** | 155** | 157** | 150** | 144** |
| Proximal tw | 30 | 27.5 | 16 | 13 | - | 14 |
| Midshaft tw | 19 | 14 | 9 | 10 | 6 | 4 |
| Distal tw | 30 | - | 11 | 15 | 7 | 10 |
| Proximal dp | - | - | - | - | - | - |
| Midshaft dp | - | 13 | - | 4 | - | 6 |
| Distal dp | 22 | 12* | - | 6 | - | 2 |
| Right Humerus | Left Humerus | Right Ulna | Left Ulna | Right Radius | Left Radius | |
|---|---|---|---|---|---|---|
| Length | 180** | 192** | 155** | 157** | 150** | 144** |
| Proximal tw | 30 | 27.5 | 16 | 13 | - | 14 |
| Midshaft tw | 19 | 14 | 9 | 10 | 6 | 4 |
| Distal tw | 30 | - | 11 | 15 | 7 | 10 |
| Proximal dp | - | - | - | - | - | - |
| Midshaft dp | - | 13 | - | 4 | - | 6 |
| Distal dp | 22 | 12* | - | 6 | - | 2 |
Measurements of forelimb elements of Pelecanimimus MCCM-LH 7777 (in mm). Abbreviations: dp, depth; tw, transversal width; *incomplete bones; **sum of measurements of all the fragments of the bone
| Right Humerus | Left Humerus | Right Ulna | Left Ulna | Right Radius | Left Radius | |
|---|---|---|---|---|---|---|
| Length | 180** | 192** | 155** | 157** | 150** | 144** |
| Proximal tw | 30 | 27.5 | 16 | 13 | - | 14 |
| Midshaft tw | 19 | 14 | 9 | 10 | 6 | 4 |
| Distal tw | 30 | - | 11 | 15 | 7 | 10 |
| Proximal dp | - | - | - | - | - | - |
| Midshaft dp | - | 13 | - | 4 | - | 6 |
| Distal dp | 22 | 12* | - | 6 | - | 2 |
| Right Humerus | Left Humerus | Right Ulna | Left Ulna | Right Radius | Left Radius | |
|---|---|---|---|---|---|---|
| Length | 180** | 192** | 155** | 157** | 150** | 144** |
| Proximal tw | 30 | 27.5 | 16 | 13 | - | 14 |
| Midshaft tw | 19 | 14 | 9 | 10 | 6 | 4 |
| Distal tw | 30 | - | 11 | 15 | 7 | 10 |
| Proximal dp | - | - | - | - | - | - |
| Midshaft dp | - | 13 | - | 4 | - | 6 |
| Distal dp | 22 | 12* | - | 6 | - | 2 |
Measurements of metacarpals of Pelecanimimus MCCM-LH 7777 (in mm). Abbreviations: dp, depth; Mc, metacarpal; L, left; R, right; tw, transversal width. *incomplete bones
| R McI | R McII | R McII | L McI | L McII | L McII | |
|---|---|---|---|---|---|---|
| Length | 48 | 59 | 58 | 44* | 44* | 45* |
| Proximal tw | 12 | 13 | 4 | 10.6 | 13 | 4.5 |
| Midshaft tw | 8 | 7 | 2 | 8 | 8 | 4.7 |
| Distal tw | 11 | 10 | 8 | - | - | - |
| Proximal dp | 3 | - | - | 9 | - | 8 |
| Midshaft dp | 2 | - | - | 5 | 4.8 | 5.5 |
| Distal dp | 6 | - | - | - | - | - |
| R McI | R McII | R McII | L McI | L McII | L McII | |
|---|---|---|---|---|---|---|
| Length | 48 | 59 | 58 | 44* | 44* | 45* |
| Proximal tw | 12 | 13 | 4 | 10.6 | 13 | 4.5 |
| Midshaft tw | 8 | 7 | 2 | 8 | 8 | 4.7 |
| Distal tw | 11 | 10 | 8 | - | - | - |
| Proximal dp | 3 | - | - | 9 | - | 8 |
| Midshaft dp | 2 | - | - | 5 | 4.8 | 5.5 |
| Distal dp | 6 | - | - | - | - | - |
Measurements of metacarpals of Pelecanimimus MCCM-LH 7777 (in mm). Abbreviations: dp, depth; Mc, metacarpal; L, left; R, right; tw, transversal width. *incomplete bones
| R McI | R McII | R McII | L McI | L McII | L McII | |
|---|---|---|---|---|---|---|
| Length | 48 | 59 | 58 | 44* | 44* | 45* |
| Proximal tw | 12 | 13 | 4 | 10.6 | 13 | 4.5 |
| Midshaft tw | 8 | 7 | 2 | 8 | 8 | 4.7 |
| Distal tw | 11 | 10 | 8 | - | - | - |
| Proximal dp | 3 | - | - | 9 | - | 8 |
| Midshaft dp | 2 | - | - | 5 | 4.8 | 5.5 |
| Distal dp | 6 | - | - | - | - | - |
| R McI | R McII | R McII | L McI | L McII | L McII | |
|---|---|---|---|---|---|---|
| Length | 48 | 59 | 58 | 44* | 44* | 45* |
| Proximal tw | 12 | 13 | 4 | 10.6 | 13 | 4.5 |
| Midshaft tw | 8 | 7 | 2 | 8 | 8 | 4.7 |
| Distal tw | 11 | 10 | 8 | - | - | - |
| Proximal dp | 3 | - | - | 9 | - | 8 |
| Midshaft dp | 2 | - | - | 5 | 4.8 | 5.5 |
| Distal dp | 6 | - | - | - | - | - |
Measurements of digit elements of Pelecanimimus MCCM-LH 7777 (in mm). Abbreviations: dp, depth; Ph, phalanx; tw, transversal width
| Right Ph. | Ph I-1 | Ph I-2 | Ph II-1 | Ph II-2 | Ph II-3 | Ph III-1 | Ph III-2 | Ph III-3 | Ph III-4 |
|---|---|---|---|---|---|---|---|---|---|
| Length | 66 | 31 | 23 | 54 | 38 | 15 | 16 | 42 | - |
| Proximal tw | 8 | 2 | - | - | - | 9 | 8 | 9 | - |
| Midshaft tw | 6 | 2 | - | 5 | - | 6 | 6 | 5 | - |
| Distal tw | 8 | 2 | 7 | 3 | - | 8 | 8 | - | - |
| Proximal dp | 13 | 12 | 10 | 15 | - | 9 | 8 | - | - |
| Midshaft dp | 7 | 9 | 8 | 7 | - | 7 | 5 | - | - |
| Distal dp | 9 | 4 | 9 | - | - | 8 | 7 | - | - |
| Left Ph. | Ph I-1 | Ph I-2 | Ph II-1 | Ph II-2 | Ph II-3 | Ph III-1 | Ph III-2 | Ph III-3 | Ph III-4 |
| Length | 60 | 33 | - | 54 | 40 | - | - | 42 | 45 |
| Proximal tw | - | 3 | - | 7 | 5 | - | - | 9 | 6 |
| Midshaft tw | - | 4 | - | 5 | 5 | - | - | 5 | 5 |
| Distal tw | - | 2 | - | 6 | 3 | - | - | 8 | 2 |
| Proximal dp | 10 | - | - | 10 | 17 | - | - | - | 12 |
| Midshaft dp | 8 | 5 | - | 7 | 7 | - | - | 7 | 6 |
| Distal dp | 11 | 2 | - | - | 2 | - | - | - | 2 |
| Right Ph. | Ph I-1 | Ph I-2 | Ph II-1 | Ph II-2 | Ph II-3 | Ph III-1 | Ph III-2 | Ph III-3 | Ph III-4 |
|---|---|---|---|---|---|---|---|---|---|
| Length | 66 | 31 | 23 | 54 | 38 | 15 | 16 | 42 | - |
| Proximal tw | 8 | 2 | - | - | - | 9 | 8 | 9 | - |
| Midshaft tw | 6 | 2 | - | 5 | - | 6 | 6 | 5 | - |
| Distal tw | 8 | 2 | 7 | 3 | - | 8 | 8 | - | - |
| Proximal dp | 13 | 12 | 10 | 15 | - | 9 | 8 | - | - |
| Midshaft dp | 7 | 9 | 8 | 7 | - | 7 | 5 | - | - |
| Distal dp | 9 | 4 | 9 | - | - | 8 | 7 | - | - |
| Left Ph. | Ph I-1 | Ph I-2 | Ph II-1 | Ph II-2 | Ph II-3 | Ph III-1 | Ph III-2 | Ph III-3 | Ph III-4 |
| Length | 60 | 33 | - | 54 | 40 | - | - | 42 | 45 |
| Proximal tw | - | 3 | - | 7 | 5 | - | - | 9 | 6 |
| Midshaft tw | - | 4 | - | 5 | 5 | - | - | 5 | 5 |
| Distal tw | - | 2 | - | 6 | 3 | - | - | 8 | 2 |
| Proximal dp | 10 | - | - | 10 | 17 | - | - | - | 12 |
| Midshaft dp | 8 | 5 | - | 7 | 7 | - | - | 7 | 6 |
| Distal dp | 11 | 2 | - | - | 2 | - | - | - | 2 |
Measurements of digit elements of Pelecanimimus MCCM-LH 7777 (in mm). Abbreviations: dp, depth; Ph, phalanx; tw, transversal width
| Right Ph. | Ph I-1 | Ph I-2 | Ph II-1 | Ph II-2 | Ph II-3 | Ph III-1 | Ph III-2 | Ph III-3 | Ph III-4 |
|---|---|---|---|---|---|---|---|---|---|
| Length | 66 | 31 | 23 | 54 | 38 | 15 | 16 | 42 | - |
| Proximal tw | 8 | 2 | - | - | - | 9 | 8 | 9 | - |
| Midshaft tw | 6 | 2 | - | 5 | - | 6 | 6 | 5 | - |
| Distal tw | 8 | 2 | 7 | 3 | - | 8 | 8 | - | - |
| Proximal dp | 13 | 12 | 10 | 15 | - | 9 | 8 | - | - |
| Midshaft dp | 7 | 9 | 8 | 7 | - | 7 | 5 | - | - |
| Distal dp | 9 | 4 | 9 | - | - | 8 | 7 | - | - |
| Left Ph. | Ph I-1 | Ph I-2 | Ph II-1 | Ph II-2 | Ph II-3 | Ph III-1 | Ph III-2 | Ph III-3 | Ph III-4 |
| Length | 60 | 33 | - | 54 | 40 | - | - | 42 | 45 |
| Proximal tw | - | 3 | - | 7 | 5 | - | - | 9 | 6 |
| Midshaft tw | - | 4 | - | 5 | 5 | - | - | 5 | 5 |
| Distal tw | - | 2 | - | 6 | 3 | - | - | 8 | 2 |
| Proximal dp | 10 | - | - | 10 | 17 | - | - | - | 12 |
| Midshaft dp | 8 | 5 | - | 7 | 7 | - | - | 7 | 6 |
| Distal dp | 11 | 2 | - | - | 2 | - | - | - | 2 |
| Right Ph. | Ph I-1 | Ph I-2 | Ph II-1 | Ph II-2 | Ph II-3 | Ph III-1 | Ph III-2 | Ph III-3 | Ph III-4 |
|---|---|---|---|---|---|---|---|---|---|
| Length | 66 | 31 | 23 | 54 | 38 | 15 | 16 | 42 | - |
| Proximal tw | 8 | 2 | - | - | - | 9 | 8 | 9 | - |
| Midshaft tw | 6 | 2 | - | 5 | - | 6 | 6 | 5 | - |
| Distal tw | 8 | 2 | 7 | 3 | - | 8 | 8 | - | - |
| Proximal dp | 13 | 12 | 10 | 15 | - | 9 | 8 | - | - |
| Midshaft dp | 7 | 9 | 8 | 7 | - | 7 | 5 | - | - |
| Distal dp | 9 | 4 | 9 | - | - | 8 | 7 | - | - |
| Left Ph. | Ph I-1 | Ph I-2 | Ph II-1 | Ph II-2 | Ph II-3 | Ph III-1 | Ph III-2 | Ph III-3 | Ph III-4 |
| Length | 60 | 33 | - | 54 | 40 | - | - | 42 | 45 |
| Proximal tw | - | 3 | - | 7 | 5 | - | - | 9 | 6 |
| Midshaft tw | - | 4 | - | 5 | 5 | - | - | 5 | 5 |
| Distal tw | - | 2 | - | 6 | 3 | - | - | 8 | 2 |
| Proximal dp | 10 | - | - | 10 | 17 | - | - | - | 12 |
| Midshaft dp | 8 | 5 | - | 7 | 7 | - | - | 7 | 6 |
| Distal dp | 11 | 2 | - | - | 2 | - | - | - | 2 |
Pelecanimimus was compared with other ornithomimosaurian taxa that were examined directly by one or more of the authors and from literature review. Anserimimus planinychusBarsbold, 1988 (MPC-D 100/300); Deinocheirus mirificusOsmólska & Roniewicz, 1970 (MPC-D 100/127/128/18); Gallimimus bullatus (MPC-D 100/300); ‘Gallimimus mongoliensis’ (Kobayashi & Barsbold, 2006) (cast FPDM-V-7216); Garudimimus brevipesBarsbold, 1981 (MCP-D 100/13); Harpymimus okladnikoviBarsbold & Perle, 1984 (MPC-D 100/29); Kinnareemimus khonkaenensisBuffetaut et al., 2009 (PW5A-123); Shenzhousaurus orientalisJi et al., 2003 (NGMC 97-4-002) and Sinornithomimus dongiKobayashi & Lü, 2003 (IVPP-V11797-9; cast FPDM-V-7217) were all inspected first-hand.
Phylogenetic analysis
We recoded the postcranial characters of Pelecanimimus polyodon from the matrix of Lee et al. (2014) [modified from Choiniere et al. (2012)], one of the most recent matrices specific to Ornithomimosauria (see Supporting Information, Appendix S1). The matrix uses 568 characters, with eight ordered multistate characters (ch. 84, 100, 227, 231, 326, 329, 361 and 471) and 96 OTUs. No character or taxa were excluded or modified from this original matrix. The matrix was edited using MESQUITE 3.61 (Maddison & Maddison, 2019). A heuristic tree search was performed in TNT 1.5 (Goloboff & Catalano, 2016) in order to find the most parsimonious trees (MPTs). The heuristic tree search used the New Technology algorithms: sectorial searches, ratchet, tree-drifting and tree fusing, using the default settings for all of them. These algorithms were applied to the recovered set of trees using the driven search with a stabilization of the consensus twice with a factor of 75. Subsequently, the results were further analysed with the tree bisection reconnection (TBR) branch-swapping algorithm. The MPTs retrieved were examined under the strict consensus. Consistency index (CI) and retention index (RI) were obtained using the ‘stats.run’ script of TNT. The branch support was tested using the methodology proposed by Goloboff et al. (2008) to calculate Bremer support values and the resampling methods bootstrap and jackknife (under default settings). The modified characters in Pelecanimimus have been mapped using the ‘character common mapping setting’ in TNT.
Three-dimensional reconstruction
The postcranial slab and counterslab of the holotype of Pelecanimimus were scanned by digital photogrammetry, using the protocol described by Mallison & Wings (2014). Pictures were taken with a regular 18–55 mm lens and some detail shots were taken with a 65 mm macro lens. The pictures were processed in Agisoft Photoscan 1.3, producing models of each slab ranging from 2 million to 5 million polygons. Only the slab and counterslab, including atlas and axis, were scanned using a CT-scan TXS320-ACTIS (TESCO Co., Yokohama, Japan) at Fukui Prefectural Dinosaur Museum (Katsuyama, Japan), under a voltage of 217 kV, current of 153 µA. For the slab, CT images have 1821 slices with an interslice spacing of 0.1 mm and the image size is 816 × 323 pixels. For the counterslab, CT images have 2231 slices with an interslice spacing of 0.1 mm and the image size of 988 × 354 pixels. The segmentation, visualization and analysis of the CT images were prepared using AMIRA (v.2019.3, Mercury Computer Systems, San Diego, CA, USA). To produce restored bones in three dimensions, the right and left halves of each bone were digitally separated from their respective slabs in ZBrush 4R6 using mask lasso and trim lasso brushes. Then, both halves were manually aligned using the symmetrical plane of fracture, filling missing parts using move and clay tube brushes. Digital infilling of missing parts was the only digital restoration, as no retrodeformation was applied to any bone. Restored bones include proatlases, atlas, axis, cervical vertebra 5, posterior dorsal vertebra, right sternal plate with some attached sternal ribs, left hand, forearm and humerus (this last, as a combination of broken pieces from left and right humeri scattered on both slabs) (see Supporting Information, Appendix S2). All restored virtual bones were colour-coded, differentiating fossil bone from digitally filled missing parts (Fig. 2).
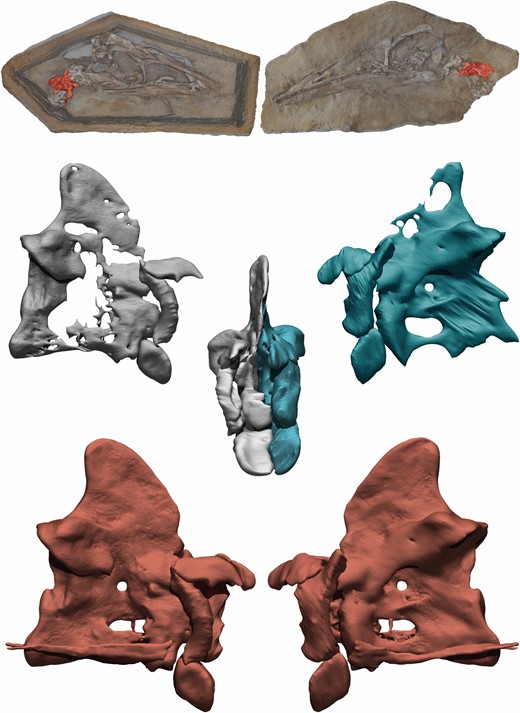
3D reconstruction process of the axis of Pelecanimimus. Original CT scan of right side in grey, original CT scan of left side in blue, complete and restored axis in red.
Intitutional abbreviations
AMNH, American Museum of Natural History, New York, USA; FPDM, Fukui Prefectural Dinosaur Museum, Fukui, Japan; IVPP, Institute of Vertebrate Paleontology and Paleoanthropology, Beijing, China; MCCM, Museo de las Ciencias de Castilla-La Mancha, Cuenca, Spain (now MUPA, Museo de Paleontología de Castilla-La Mancha); MPC-D, Paleontological Center of Mongolia, Ulaan Bataar, Mongolia; NGMC, National Geological Museum of China, Pekin, China; RTMP, Royal Tyrrell Museum of Palaeontology, Alberta, Canada; ZIN PH, Paleoherpetological Collection, Zoological Institute, Russian Academy of Sciences, Saint Petersburg, Russia.
Systematic palaentology
Theropoda Marsh, 1881
Coelurosauria von Huene, 1914
Ornithomimosauria Barsbold, 1983
Macrocheiriformes clade nov.
Definition:
Ornithomimosaur clade, including the most recent common ancestor of Ornithomimus and Pelecanimimus and all its descendants (rank free, node-based).
Etymology:
This rank-free clade name is derived from the Greek μάκρος (makros), long, χειρία, (cheiria), hand and φόρμες (formes), forms, in reference to the enlargement of the manus (metacarpal I and distal phalanges) of this taxon.
Pelecanimimus polyodon Pérez-Moreno et al., 1994
Type specimen:
MCCM-LH 7777 (holotype).
Described material:
The anterior-half part of the postcranial skeleton of the holotype of Pelecanimimus polyodon MCCM-LH 7777, including a complete pectoral girdle (including both ossified sternal plates), both broken but complete forelimbs and manus, all cervical vertebrae (including atlas and axis), three anteriormost dorsal vertebrae and three posterior dorsal vertebrae, cervical, sternal and dorsal ribs and uncinate processes. The specimen is housed in the Museo de Paleontología de Castilla-La Mancha (MUPA, formerly MCCM).
Occurrence:
Las Hoyas fossil site, Calizas de La Huérguina Formation, Upper Barremian (Lower Cretaceous); La Cierva township, Cuenca, Spain.
Characters to be included in the revised diagnosis based on the postcranial skeleton:
Ornithomimosaurian theropod with the following unique combination of postcranial characters within Ornithomimosauria (autapomorphies among Ornithomimosauria with asterisk): (1) neural spine of the axis dorsally tall, with a posterodorsally tapered, posteriorly projected and rounded apex*; (2) fan-shaped neural spine on posteriormost dorsal vertebrae in lateral view*; (3) evenly developed distal condyles in metacarpal I; and (4) poor divergence between metacarpal II and metacarpal I.
Description and comparison
Axial skeleton
Cervical vertebrae:
Ten cervical vertebrae are preserved. Both main fossil slabs include the 3rd to 10th cervical vertebrae. The atlas, axis and the anterior-half of the 3rd vertebra are in articulation with the skull in separate slabs. Overall, the right side of the axial skeleton is better preserved than the more damaged left one. All cervical vertebrae have amphiplatyan articular faces, like in other coelurosaurians (Holtz, 1998). Despite the generally worse preservation, cervical ribs are better preserved on their left side, in articulation with each vertebra, with parapophyses hidden. From 6th to 10th cervical vertebrae, the centra are described based on their right side.
The two proatlases (Figs 3, 4) are well preserved in both slabs. They are triangular in lateral view and anteroposteriorly elongated. Medially, the proatlas is concave. The right proatlas is articulated with the exoccipital anteriorly and with the atlantal arch posteriorly. Aside from Pelecanimimus, proatlases had been reported before only in Sinornithomimus (Kobayashi & Lü, 2003) and Struthiomimus (Makovicky et al., 2004) within Ornithomimosauria, where both proatlases have the same orientation and shape. Compared with other coelurosaurian proatlases, Pelecanimimus proatlases lack the marked tripartite arrangement seen in those of the dromaeosaurid Tsaagan mangasNorell et al., 2006, being more similar to those observed in the troodontid SinovenatorXu et al., 2002 (Yin et al., 2018), although these were located dorsoventrally oriented in the posterior end of the skull.
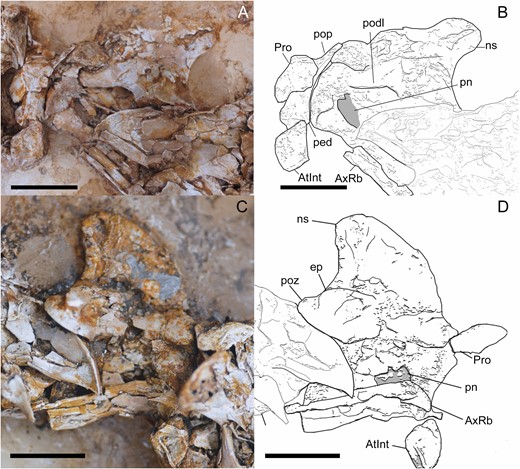
Atlas and axis of Pelecanimimus (MCCM-LH 7777), photo (A), drawing (B). A, B, left side. C, D, right side. Abbreviations: AtInt, atlas intercentrum; AxRb, axial rib; ep, epipophysis; ns, neural spine; ped, pedunculum; pn, pneumatic foramina; podl, postzygodiapophyseal lamina; pop, posterior process; poz, postzygapophysis; Pro, proatlas. Scale bars: 10 mm.
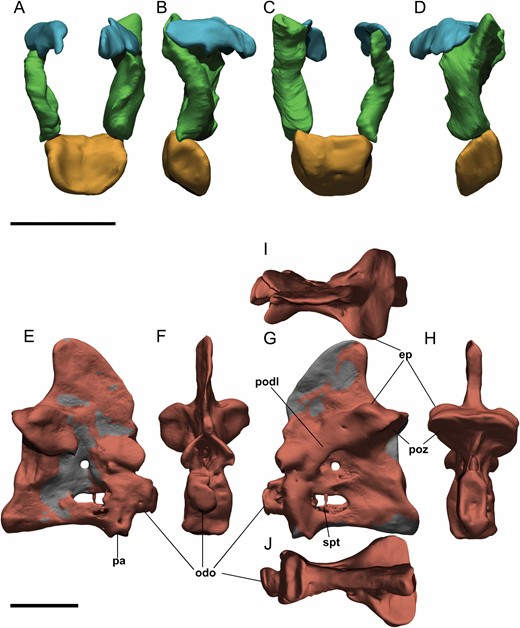
Digital atlas and axis of Pelecanimimus (MCCM-LH 7777). A–D, atlas in anterior (A), lateral right (B), posterior (C) and lateral left (D) views. E–J, axis in lateral right (E), anterior (F), lateral left (G), posterior (H), dorsal (I) and ventral (J) views. Abbreviations: ep, epipophysis; pn, pneumatic foramina; odo, odontoid; pa, parapophysis; podl, postzygodiapophyseal lamina; poz, postzygapophysis. Scale bars: 10 mm.
The atlantal intercentrum (Figs 3, 4) is complete, articulated with the occipital condyle and disarticulated from the axial centrum. Both atlantal neural arches are preserved and not fused with the intercentrum, as in Sinornithomimus (Kobayashi & Lü, 2003) and Garudimimus (MPC-D 100/13), troodontids (Yin et al., 2018), dromaeosaurids and Aves (Norell & Makovicky, 2004), although both elements are also disarticulated in the basal theropod HerrerasaurusReig, 1963 (Sereno & Novas, 1994). The atlantal intercentrum is U-shaped in anterior view due to the concave anterior surface for articulation with the occipital condyle, which is slightly anterodorsally oriented. It is triangular in lateral view, and has a circular depression for the articulation of the non-preserved atlantal ribs, which are especially prominent on the right side. Posteriorly, a ridge divides the intercentrum into two surfaces of articulation. The posterodorsally oriented surface is for the articulation with the odontoid and the posteroventral one is for the articulation with the axis intercentrum. Dorsally, the anteroventrally inclined surface articulates with the atlantal arches.
As already mentioned, both neural arches of the atlas are preserved (Fig. 4). They are triradiate in lateral view, although the right one is incomplete anteriorly. The basal pedicle for the articulation with the intercentrum is anteroposteriorly wide but shorter than the posterior process, as in Gallimimus bullatus and Sinornithomimus (Kobayashi & Lü, 2003) and unlike Garudimimus (MPC-D 100/13), which has a pedicle longer than the posterior process. The postzygapophyses are long, stout and posterodorsally projected, with a concave and ventrolaterally oriented articular surface. The left neural arch preserves a strong developed and triangular dorsal process, which is not preserved on the right arch. In the anterior projection, the anterior processes are tab-like and medially developed and curved.
The axis is also complete and preserved in its anatomical position, articulated with the 3rd cervical vertebra (Figs 3, 4). The odontoid is articulated to the axis on its right side. The neural spine of the axis is tall compared with those of the following vertebrae. It is triangular and transversally compressed as a thin lamina. This neural spine has a posterodorsally tapered, posteriorly projected and rounded apex. The neural spine is distinct from the less dorsoventrally tall, more anteroposteriorly long and dorsally straigther apex in other ornithomimosaurs, such as Garudimimus (MPC-D 100/13; Kobayashi & Barsbold, 2005b) and Sinornithomimus (FPDM-V-7217; Kobayashi & Lü, 2003). The posterior articular surface is tilted anteriorly in lateral view like in others ornithomimosaurs (Kobayashi & Barsbold, 2005b). However, the anterior surface is almost vertically oriented. The centrum is anteroposteriorly short and dorsoventrally tall like Garudimimus (MPC-D 100/13), but unlike the long and slender one of Gallimimus (MPC-D 100/11; Osmólska et al., 1972) and Ornithomimus sp. (Makovicky, 1995; Kobayashi & Barsbold, 2005b). The centrum has pneumatic foramina at its midlength, which are anteroposteriorly elongated and oval, like those observed also in Gallimimus (MPC-D 100/11). In contrast, Garudimimus lacks pneumatic foramina in its axis (MPC-D 100/13; Kobayashi & Barsbold, 2005b). Dorsal to this pneumatic foramen, an anteroposteriorly directed and thin ridge/septum is developed beyond the middle of the centrum. The postzygapophyses of the axis are more rounded than those of the following vertebrae, and are scarcely projected posteriorly like in Harpymimus (MPC-D 100/29) and unlike the most posteriorly projected postzygapophyses in Garudimimus (MPC-D 100/13). The epipophyses are located above the postzygapophyses, posteriorly oriented and, although poorly developed, they are more developed than those of the following cervical vertebrae. Derived ornithomimosaurs, such as Sinornithomimus, Gallimimus (MPC-D 100/11) and Ornithomimus, have more developed epipophyses in the axis (Kobayashi & Lü, 2003). The diapophyses are laterally reduced. The long and thin axial ribs are articulated with the axis on both sides.
All postaxial cervical vertebrae have centra that are longer than tall, especially the anterior ones (Figs 5, 6). Therefore, posterior cervical vertebrae (Fig. 6) are shorter and more quadrangular in lateral view than anterior ones, which are more rectangular and long, as in other ornithomimosaurs such as Harpymimus (MPC-D 100/29). The cervical centra of some ornithomimosaurs, such as Nqwebasaurusde Klerk et al., 2000, Archaeornithomimus, Anserimimus and the Bissekty taxon (ZIN PH 130/16), have ventrolateral crests running from the parapophyses to the ventral surface (Smith & Galton, 1990; Bronowicz, 2011; Choiniere et al., 2012; Sues & Averianov, 2016). These crests are prominent in the 8th and 9th cervical vertebrae of Pelecanimimus in right-lateral view (Fig. 6). In these vertebrae, the ventrolateral crests are projected from the parapophyses to the ventral surface of the centrum in lateral view. The ventrolateral crests delimit the ventral surface laterally. In the 10th cervical vertebra (Fig. 6), the centrum seems to have a small bulge, ventral to the anterior surface, similar to that of the carotid process described in AustroraptorNovas et al., 2009. However, it is not visible in the rest of the cervical vertebrae. The centra lack ventral keels, ridges or grooves. The cervical pneumatic foramina are oval and anteroposteriorly long and, although present in all vertebrae, the best preserved one is that on the 8th vertebra (Fig. 6). The cervical pneumatic foramina in Pelecanimimus are single, unlike the double pneumatic foramina, split by a posterodorsal–anteroventral oriented ridge, of Harpymimus (MPC-D 100/29; Kobayashi & Barsbold, 2005a). The diapophyses are triangular in lateral view, with their lateral process ventrally oriented. The articular surfaces of the anterior prezygapophyses are oval, strongly anteroposteriorly elongated, slightly convex and anteroventrally inclined like other ornithomimosaurs, such as the taxon from the Bisskety Formation (Sues & Averianov, 2016) and Gallimimus (MPC-D 100/11), but not as extremely ventrally projected like those observed in Nqwebasaurus (Choiniere et al., 2012), and unlike the non-ventrally curved ones of Garudimimus (MPC-D 100/13). The posteriormost cervical prezygapophyses are less anteriorly projected, reaching only as far as the anterior surface of the centrum, and their articular surfaces are horizontal (Fig. 6). The lateral surfaces of the prezygapophyses have a foramen in all cervical vertebrae. The postzygapophyseal facets have the same morphology as prezygapophyseal ones, but are posterodorsally oriented and slightly concave. However, the most posterior ones are horizontally oriented, like in Garudimimus (MCPD 100/13) and unlike the outwardly curved ones in more derived ornithomimids (Kobayashi & Barsbold, 2005b). The postzygapophyses are long, with those of the posteriormost cervical vertebrae especially projected further posteriorly, a tendency similar in other ornithomimosaurs (Osmólska et al., 1972; Kobayashi & Lü, 2003; Makovicky et al., 2004; Kobayashi & Barsbold, 2005a, b; Lee et al., 2014; Sues & Averianov, 2016). The epipophyses are poorly developed, being nearly absent in the posterior cervical vertebrae (Fig. 6), like in other ornithomimosaurs (e.g. Kobayashi & Barsbold, 2005a). In the anterior cervical vertebrae (Fig. 5), the more developed epipophyses are more pronounced than the faintly marked ones observed in other ornithomimosaurs, such as Gallimimus (Osmólska et al., 1972), Nqwebasaurus (Choiniere et al., 2012), the Bissekty taxon (Sues & Averianov, 2016) or Garudimimus (MPC-D 100/13; Kobayashi & Barsbold, 2005b). The neural arch is anteroposteriorly longer in anterior vertebrae than in posterior ones; so anterior cervical centra are less posteriorly developed than the posterior end of the neural arch, as in other basal ornithomimosaurs (Choiniere et al., 2012). The neural spines are only visible in anterior cervical vertebrae (Fig. 5), probably due to posterior cervical vertebrae lacking neural spines or having extremely reduced ones, as other ornithomimosaurs (Kobayashi & Barsbold, 2005; Sues & Averianov, 2016). Their height is relatively short, although slightly taller than in other ornithomimosaurs, such as Nqwebasaurus (De Klerk et al., 2010), Harpymimus (MPC-D 100/29; Kobayashi & Barsbold, 2005a), the Bissekty Formation taxon (Sues & Averianov, 2016), Garudimimus (MPC-D 100/13) and Gallimimus (Osmólska et al., 1972). The spines are anteroposteriorly long and are located slightly posterior to the midlength of the neural arch, like in Garudimimus (MPC-D 100/13) and Sinornithomimus (Kobayashi & Lü, 2003), but unlike in derived ornithomimids, in which it is positioned in the middle of the neural arch (Kobayashi & Lü, 2003).
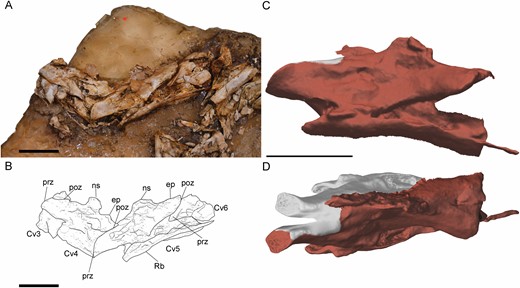
Anterior cervical vertebrae of Pelecanimimus (MCCM-LH 7777) in left side. A,, B cervical vertebrae from 3–6 in connection, photo (A), drawing (B). C, D, digitally reconstructed fifth cervical vertebra in 3D in lateral view (C) and in dorsal view (D) with restored areas in grey. Abbreviations: ep, epipophysis; ns, neural spine; podl, postzygodiapophyseal lamina; pop, posterior process; poz, postzygapophysis; prz, prezygapophysis; Rb, rib. Scale bars: 20 mm.

Posterior cervical vertebrae of Pelecanimimus (MCCM-LH 7777) in right side. A B, cervical vertebrae (Cv) from 3–10 in connection, photo (A), drawing (B). Abbreviations: CRb, cervical rib, ns, neural spine; pop, posterior process; poz, postzygapophysis; pozrg, postzygapophysis ridge; prz, prezygapophysis; prerg, prezigapophysis ridge. Scale bars: 20 mm.
Anterior cervical ribs (Fig. 5) are strongly anteroposteriorly elongated, especially their posterior processes, and are dorsoventrally thinner than posterior ones. Posterior cervical ribs (Fig. 6) are triangular, with strongly projected anterior process, like in Harpymimus (MPC-D 100/29). The capitular and tubercular region is dorsoventrally thick.
Dorsal vertebrae:
The preserved dorsal vertebrae are the anteriormost three (Fig. 7) and three posterior dorsal vertebrae in an isolated block, which also preserves part of the forelimb (Fig. 8). The centra of the three anterior vertebrae are laterally compressed at midlength and have oval pneumatic foramina (Fig. 7), posteroventral to the parapophysis, unlike Nqwebasaurus, (Choiniere et al., 2012; Sereno, 2017) or Deinocheirus (Lee et al., 2014). The condition in Pelecanimimus is more similar to that of Harpymimus (MPC-D 100/29; Kobayashi & Barsbold, 2005a) or Gallimimus (Osmólska et al., 1972). The parapophysis on the 3rd vertebra is dorsoventrally tall, oval and concave. It occupies the anterolateral surface of the centrum, and is slightly dorsally displaced but does not reach the neural arch, as in Nqwebasaurus (Choiniere et al., 2012) and Harpymimus (Kobayashi & Barsbold, 2005a). A ridge runs horizontally from its ventral surface toward the middle of the centrum, delimiting a dorsal depression like in the Bissekty taxon (Sues & Averianov, 2016) and Archaeornithomimus (Smith & Galton, 1990), and unlike Garudimimus (MPC-D 100/13). The 3rd dorsal vertebra has a ventral keel in the anteroventral surface of the centrum like Nqwebasaurus (Choiniere et al., 2012), the Bissekty Fm taxon (Sues & Averianov, 2016) and Ornithomimus (Kobayashi & Barsbold, 2005b). This ventral keel is rounded and well developed, likely an osteological correlate for the origin of m. longus colli ventralis (Currie & Zhao, 1993; Snively & Russell, 2007). In Shenzhousaurus (NGMC 97-4-002), the ventral keels are in the posterior dorsal vertebrae and cannot be observed in the anterior ones, because the latter are not preserved in the fossil. The diapophyses are dorsally oriented, proximodistally long, dorsoventrally thin and anteroposteriorly narrow, and distinct from the laterally oriented transverse processes in Harpymimus (MPC-D 100/29). The anterior and posterior centrodiapophyseal laminae are extremely thin, like in the Bissekty taxon (Sues & Averianov, 2016), especially the anterior one, and unlike the well-developed ones in Harpymimus (MPC-D 100/29). The centrodiapophyseal fossa is well excavated like in Shenzhousaurus (NGMC 97-4-002; Ji et al., 2003). The articular surfaces of the prezygapophyses are more horizontally oriented than those observed in cervical vertebrae, as in Harpymimus (MPC-D 100/29). A posteriorly directed ridge runs through its lateral surface, bordered ventral to the articular surface, like in Garudimimus (MPC-D 100/13). The prezygocentrodiapophyseal fossa is wide and deeply excavated, like those described in Shenzhousaurus (NGMC 97-4-002; Ji et al., 2003). In the 6th and 7th dorsal vertebrae of Garudimimus (MPC-D 100/13), this fossa has a posterodorsal-anteroventrally oriented septum, parallel to the anterior centrodiapophyseal lamina, which is absent in the vertebrae of Pelecanimimus. The centrodiapophyseal fossa is also anteroposteriorly wide. The neural canal in the 3rd dorsal vertebra is transversally narrow and dorsoventrally long, but it is deformed due to lateromedial compression. The neural spine is dorsoventrally poorly developed but anteroposteriorly long and sheet-like, especially at its distal end that is slightly posteriorly projected. In this respect it is similar to those of Harpymimus (MPC-D 100/29) and unlike Garudimimus (MPC-D 100/13), in which the neural spines are taller and anteroposteriorly shorter than those of Pelecanimimus.
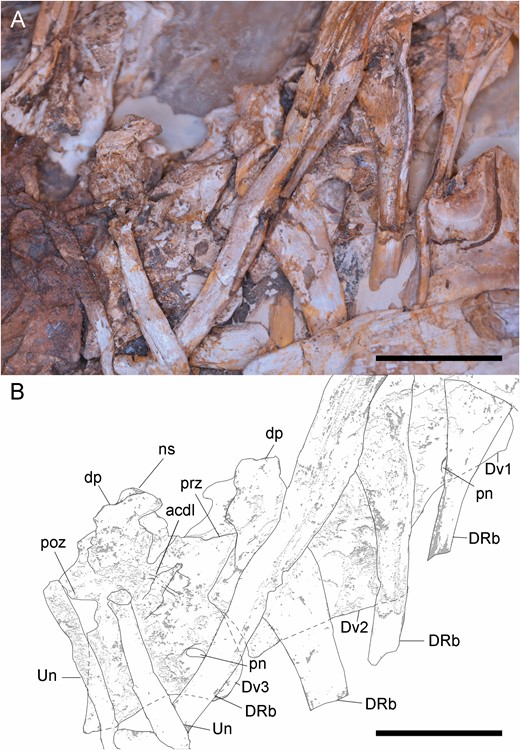
Anterior dorsal vertebrae of Pelecanimimus (MCCM-LH 7777) in right side. A, B, dorsal vertebrae (Dv) from 1–3 in connection, photo (A), drawing (B). Abbreviations: acdl, anterior centrodiapophyseal lamina; dp, diapophysis; DRb, dorsal rib, ns, neural spine; pn, pneumatic foramina; pop, posterior process; poz, postzygapophysis; prz, prezygapophysis; un, uncinate process. Scale bars: 20 mm.
Posterior dorsal vertebrae and left forearm of Pelecanimimus (MCCM-LH 7777). A, B, isolated posterior dorsal vertebrae (Dv) and left forearm in anatomical position, photo (A), drawing (B). C, close-up view of the neural spine of the posterior dorsal vertebra. D–I, digital reconstruction of the posteriormost preserved dorsal vertebra in dorsal (D), anterior (E), lateral left (F), posterior (G), lateral right (H) and ventral (I) views with restored areas in grey. Abbreviations: acdl, anterocentrodiapophyseal lamina; acdl, anterior centrodiapophyseal lamina; apns, anterior process of neural spine; dp, diapophysis; DC, distal carpal; DRb, dorsal rib, kl, keel; Mc, metacarpal; ns, neural spine; pa, parapophysis; pn, pneumatic foramina; pcdl, posterior centrodiapophyseal lamina; pop, posterior process; poz, postzygapophysis; ppns, posterior process of neural spine; prz, prezygapophysis, R, radius; Rb, rib; Scu, scapholunare: U, ulna. Scale bars: A, B, 50 mm; C, 20 mm.
The posterior dorsal vertebrae are represented by two complete centra and two complete neural arches and the fragment of another one (Fig. 8). The neural arch and the centrum are unfused and separated from each other. The centra have a strong hourglass shape as in other ornithomimosaurs, like Garudimimus (MPC-D 100/13), Kinnareemimus (PW5A-123; Buffetaut et al., 2009), Gallimimus (MPC-D 100/11) or Harpymimus (MPC-D 100/29), and are anteroposteriorly longer than dorsoventrally tall. Compared with those of the anterior dorsal vertebrae, the posterior centra are longer than tall. There are weakly developed anterior and posterior keels on the ventral surfaces of the centra. The ventral surfaces also bear strongly marked longitudinal ligament scars, which are also present on the posterior lateral surface. The parapophyses are visible in two posterior neural arches. These parapophyses are located on the anterolateral surface of the neural arches, slightly below the prezygapophyses and transverse processes. Peduncles on the parapophyses are not developed, unlike Deinocheirus (Lee et al., 2014); their overall morphology is oval. Pneumatic foramina are not developed in these posterior vertebrae, but the lateral surface of the centrum is concave. The transverse processes are slightly posterodorsally oriented, unlike the anterodorsally oriented ones in Deinocheirus (Lee et al., 2014), but like other ornithomimosaurs. These processes are mediolaterally wide and anteroposteriorly long. The anterior and posterior centrodiapophyseal laminae are well developed and robust, especially the anterior one, unlike Garudimimus (MPC-D 100/13), which has poorly developed anterior centrodiapophyseal laminae in the posterior vertebrae. The prezygapophyses and postzygapophyses are articulated, and their articular surface is horizontal. These articular surfaces have a dorsal horizontal ridge in lateral view. The prezygapophyses and postzygapophyses are slightly anteriorly projected and posteriorly overhang the centrum, respectively, less than those of the anterior dorsal vertebrae, which is the same condition observed in other ornithomimosaurs such as Harpymimus (MCPD 100/29). The prezygocentrodiapophyseal fossa is extremely anteroposteriorly short; contrary to the centrodiapophyseal fossa, which is strongly anteroposteriorly elongated and well excavated. The postzygocentrodiapophyseal fossa is also well developed, although shorter than the centrodiapophyseal one.
The neural arches are compressed by compaction of the fossil. The neural spines are slightly taller than those on anterior dorsal vertebrae and anteroposteriorly longer, occupying the full length of the neural arches. However, the spines are not as tall as those observed in Garudimimus (Kobayashi & Barsbold, 2005b). These posterior dorsal neural spines have anteroposteriorly expanded with fan-shaped distal ends, developing strongly anterior and posterior processes that are almost in contact with those of the preceding and following neural spines (Fig. 8C). This morphology is similar to that observed in some compsognathids, such as CompsognathusWagner, 1861 (Ostrom, 1978; Peyer, 2006) and HuaxiagnathusHwang et al., 2004 (Hwang et al., 2004), but different from other basal ornithomimosaurs. In Shenzhousaurus (NGCM 97-4-002), only the posterodorsal end is projected but does not have the marked fan-shape. Harpymimus (MPC-D 100/29) has anterior and posterior processes in the 8th and 9th dorsal vertebrae, but they are not as strongly anteroposteriorly expanded as in Pelecanimimus, and the posteriormost spines are rectangular and relatively high. However, the condition in the basal-most known ornithomimosaur, Nqwebasaurus, is still unknown. Other ornithomimosaurs also have a slightly anteroposteriorly flared distal end of the spine, such as Garudimimus (MPC-D 100/13) in the 8th dorsal vertebra or Gallimimus (MPC-D 100/11) in some posterior vertebrae. However, the morphology is still rectangular, they are taller and unlike the fan-shape of the posterior dorsal spines of Pelecanimimus.
Some dorsal ribs are preserved in articulation with vertebrae, although most are disarticulated. The capitulum is anteriorly elongated and dorsoventrally thick in the anterior ribs. By contrast, the tuberculum is not strongly developed, and is rounded on its posterior surface. The first dorsal rib is the best-preserved one, and its shaft is anteroposteriorly flared in the proximal end like in Shenzhousaurus (NGMC 97-4-002; Ji et al., 2003).
Associated with the dorsal ribs, close to the sternum, there are four possible uncinate processes, which are partially obscured by other ribs (Fig. 9). The two more complete ribs are parallel to each other, distally curved, flattened transversally, long and in articulation with two dorsal ribs. The articulations with these dorsal ribs are bell-shaped, which is a typical morphology in the uncinate processes in non-avian theropods and birds (Codd, 2004, 2010; Codd et al., 2008). This author also asserts that the typical uncinate processes are distally tapered, but the distal ends of those in Pelecanimimus are missing or non-visible. The flattened condition is also observed in other non-avian theropods uncinate processes, as in Oviraptor philoceratopsOsborn, 1924 (AMNH 6517) or Velociraptor mongoliensisOsborn, 1924 (MPD 100/25) (Codd et al., 2008). However, in Pelecanimimus, the taphonomic condition of the whole fossil led to flattened bones. Although its association and some described features seem to indicate that these bones could be identified as uncinate processes, all above-mentioned characteristics are not clearly observed in the fossil and could have been altered by taphonomical processes.
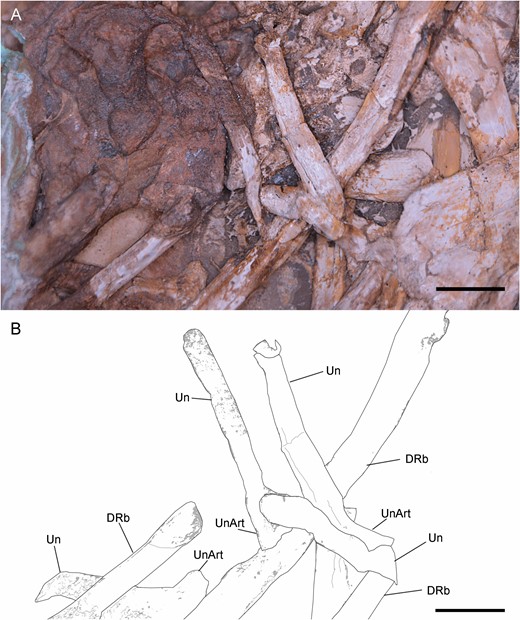
Uncinate processes of Pelecanimimus (MCCM-LH 7777): photo (A), drawing (B). Abbreviations: Drb, dorsal rib; Un, uncinate process; UnArt, articulation of uncinate process. Scale bars: 10 mm.
APPENDICULAR SKELETON
Sternum:
The sternum of Pelecanimimus comprises two sternal plates. These paired sternal plates are unfused and disarticulated. The left sternal plate is partially hidden by the sediment matrix, other pectoral girdle elements, ribs and the right sternal plate. The latter is preserved perpendicular to the left sternal plate and is almost complete and well preserved (see Supporting Information, Appendix S3). This right sternal plate (Fig. 10) has an oval shape, anteroposteriorly longer than mediolaterally wide. Its lateral margin is thicker than in the medial margin, where it is partially fragmented. The anteriormost part of the sternal plate is anteriorly tapered. Its anterolateral margin is anteromedially inclined, slightly thicker than the medial margin and slightly deformed. This area corresponds to the coracoid facet, which articulates with the coracoid. The lateral surface seems to have a pitted groove for articulation, like that observed in Velociraptor MPC-D 100/985 (Norell & Makovicky, 1997). However, this entire coracoid facet is partially broken along its lateral margin. Posteriorly, the facet is limited by a small, laterally oriented and incomplete process that could correspond to the sternocoracoidal process, observed in other theropods such as Tawa hallae Nesbitt et al., 2009 (Bradley et al., 2019) and other maniraptoriforms (e.g. Norell & Makovicky, 1997, 1999; Clark et al., 1999; O’Connor et al., 2015). The costal processes of Pelecanimimus are located posterior to this process, along the lateral margin of the sternal plate. There are three well-preserved, rounded and evenly spaced costal processes, two of which are in articulation with two sternal ribs. However, two other sternal ribs are preserved posteriorly, disarticulated and associated with the lateral margin of the right sternal plate. The posterior end of the sternal plate is mediolaterally wider than the anterior one, but it is incomplete and featureless. Therefore, the presence of the posterior process (xiphoid process) of other theropods, such as oviraptorosaurs (Clark et al., 1999; Balanoff & Norell, 2012; Wei et al., 2013; Funston & Currie, 2016; Norell et al., 2018), is unknown in Pelecanimimus. The four sternal ribs are arranged parallel to each other, and the ends that attach to the costal processes are expanded. Medially, the right sternal plate is deformed by taphonomy, resulting in it being curved with respect to the whole orientation of the plate. Its medial margin is thinner than the lateral one. It is partially crushed, fragmented and the midline between both paired plates is only preserved in the anteriormost region. This midline is straight in dorsal view, and slightly thickened. The left sternal plate is fragmented, poorly preserved and featureless, but the mediolateral width is practically the same than the right plate.
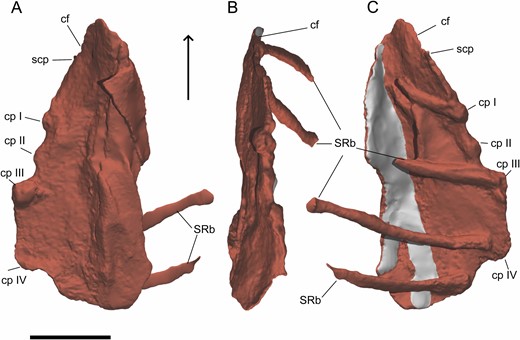
Digital reconstruction of the right sternal plate of Pelecanimimus (MCCM-LH 7777) in ventral (A), lateral (B) and dorsal (C) views. Arrow points to anterior. Restored areas in grey. Abbreviations: cf, coracoid facet; cp, costal process; scp, sternocoracoidal process; S RB, sternal ribs. Scale bar: 20 mm.
Scapulocoracoids and furcula:
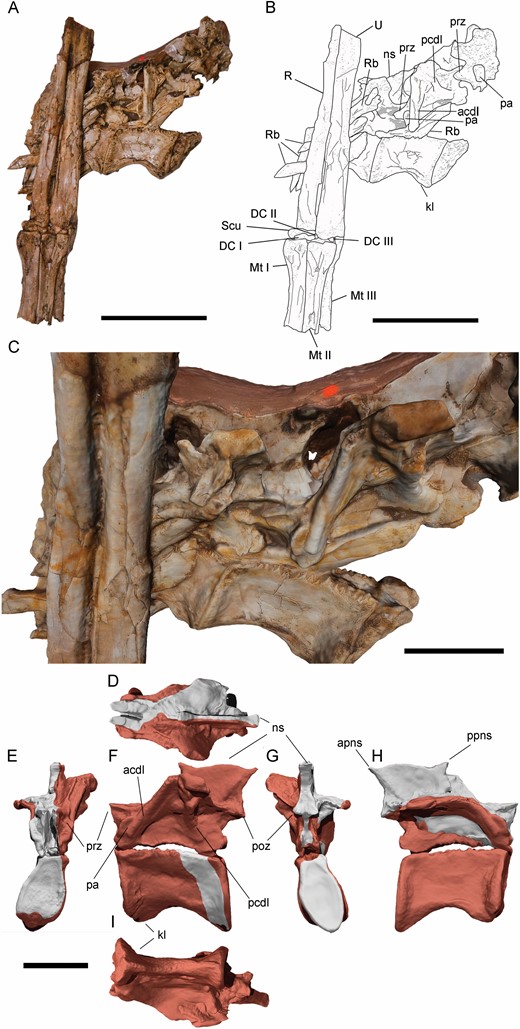
Both scapulocoracoids are preserved, but scapulae and coracoids are unfused and disarticulated (Fig. 11). The ventral ends of both scapulae are incomplete, as well as the dorsal ends. The shafts of the scapulae are slender, lateromedially comprised and anteroposteriorly expanded at the dorsal and ventral ends, like in other ornithomimosaurs (e.g. MPC-D 100/11; MPC-D 100/29; Kobayashi & Lü, 2003; Makovicky et al., 2009). These shafts are also slightly anteriorly and laterally curved, like in Harpymimus (MPC-D 100/29). The left scapula preserves the dorsal extension of the glenoid, and a shallow fossa is preserved in its posterior surface, just dorsally to the glenoid. This fossa is also observed in other ornithomimosaurs, such as Harpymimus (MPC-D 100/29; Kobayashi & Barsbold, 2005a), the Bissekty taxon (Sues & Averianov, 2016), Beishanlong (Makovicky et al., 2009), Deinocheirus (MPC-D 100/18; Osmólska & Roniewicz, 1970), Sinornithomimus (Kobayashi & Lü, 2003), Gallimimus (MPC-D 100/11; Osmólska et al., 1972), Anserimimus (MPC-D 100/300) and Struthiomimus (Nicholls & Russell, 1985). The dorsal end of the acromion process is preserved, but incomplete. It is dorsoventrally long and anteroposteriorly narrow, like in other ornithomimosaurs (De Klerk et al., 2000; Makovicky et al., 2009). The left coracoid is visible in medial view and the right one in dorsomedial view, although its lateral surface is also partially visible. Overall, the coracoid is dorsoventrally low, which is the common morphology for Ornithomimosauria (Nicholls & Russell, 1985: proposed to Ornithomimidae; Makovicky et al., 2004) and alvarezsauroids [supplementary data of Choiniere et al. (2010)], although it is dorsoventrally taller in Gallimimus (MPC-D 100/11) and Deinocheirus (MPC-D 100/18). The posteroventral processes are well developed and further projected in both coracoids, like in other ornithomimosaurs (Makovicky et al., 2004); it is separated from the coracoid glenoid by a dorsoventrally narrow notch. However, the glenoid is almost completely hidden in both coracoids, with the infraglenoid buttresses not visible at all. The coracoid tubercle is visible in the right coracoid in lateral view. This tubercle is a long and horizontal ridge extending from the coracoid centre through the dorsal border of the posteroventral process, like in other ornithomimosaurs, such as Nqwebasaurus (De Klerk et al., 2000), Bissekty taxon (Sues & Averianov, 2016), Archaeornithomimus and Beishanlong (Makovicky et al., 2009). The medial surface is concave, and the dorsal border of the posteroventral process is developed as a ridge, like in others ornithomimosaurs as Gallimimus (MPC-D 100/11).
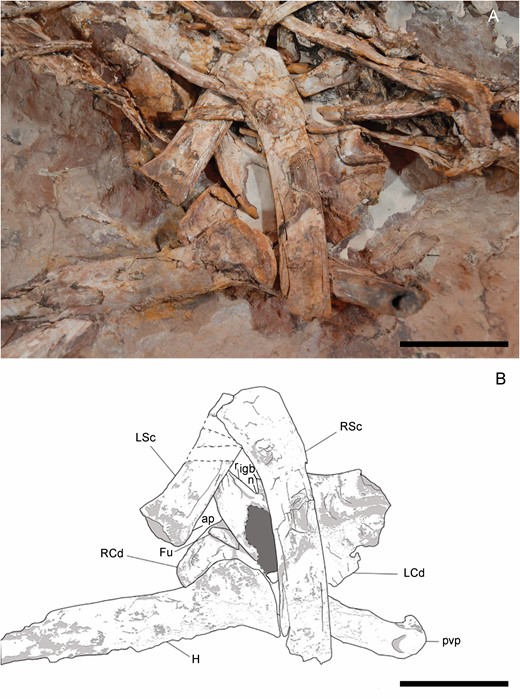
Left and right scapulocoracoids of Pelecanimimus (MCCM-LH 7777): photo (A) and drawing (B). Abbreviations: ap, acromion process; Fu, furcula; H, humerus; igb, infraglenoid butress; L Cd, left coracoid; L Sc, left scapula; n, notch; r, ridge; R Cd, right coracoid; R Sc, right scapula; pvp, posteroventral process. Scale bars: 50 mm.
A small fragment under the left coracoid is interpreted as the epicleidial process of the furcula (Fig. 11). This process is rounded, not tapering at all in the end, and the articular surface is not visible. Only Deinocheirus has a complete furcula preserved among Ornithomimosauria (Lee et al., 2014), but the scarce information in Pelecanimimus does not allow a comparison.
Humeri:
The distal-most fragment of the left humerus is positioned in anterior view, and its shaft is long and dorsoventrally thin (Fig. 12). This is also the condition in the right humerus (Fig. 13). The condyles of the humerus are slightly dorsoventrally compressed (Figs 12, 14), like in Harpymimus (MPC-D 100/29) and Nqwebasaurus (De Klerk et al., 2000), but the distal end is not transversally expanded, like in the latter taxon (De Klerk et al., 2000; Sereno, 2017). On the ventral surface, a supracondylar fossa is slightly developed above the condyles, similar to that observed in the humerus of Harpymimus (MPC-D 100/29) and unlike the strongly excavated one in Gallimimus (MPC-D 100/11), and it is bordered by anterior and posterior ridges. Dorsal to the posterior extent of the condyles the olecranon depression is shallow, like in other ornithomimosaurs (e.g. Nicholls & Russell, 1985; Kobayashi & Lü, 2003; Sereno, 2017). On the anterior border, the ectepicondyle is developed as a ventral ridge (Fig. 14). The midlength shaft has the same thickness as the shaft at the distal end (Fig. 12). The proximal end increases in width proximally, and it is triangular in this view. The deltopectoral crest is slightly developed as a triangular flange, located along the anteroventral border of the proximal end (Figs 12, 14). Its tip is oriented posteriorly. It is different from those observed in Nqwebasaurus, Gallimimus, Anserimimus or Deinocheirus, which are strongly developed (MPC-D 100/11; MPC-D 100/300; MPC-D 100/18; Osmólska & Roniewicz, 1970; De Klerk et al., 2000; Sereno, 2017) and also weaker than that of Sinornithomimus (Kobayashi & Lü, 2003), but similar to the weakly developed one of the Bissekty taxon (Sues & Averianov, 2016), Archaeornithomimus (Smith & Galton, 1990) and Struthiomimus (Nicholls & Russell, 1985). The deltopectoral crest is gradually reduced proximally and distally along the shaft. In the middle of its ventral surface, there is a foramen (Fig. 12) that probably was produced by collapse of the surface layer of the bone. The fragments associated with the proximal end (Fig. 12) seem to indicate that the humerus was proximally anteroposteriorly expanded and quadrangular. This morphology is like that observed in the Harpymimus humerus (MPC-D 100/29; Kobayashi & Barsbold, 2005a) and those of other ornithomimosaurs (Makovicky et al., 2009; Sues & Averianov, 2016), but distinct from the spherical one in Sinornithomimus (Kobayashi & Lü, 2003). The internal tuberosity is developed as a posteroventrally oriented flange (Fig. 13), slightly ventrally located with respect to the humeral head, with which both are confluent, as in all ornithomimosaurs (e.g. MPC-D 100/300; Kobayashi & Lü, 2003; Sues & Averianov, 2016). The humeral head is slightly visible in posterior view (Fig. 13), but it is not possible to determine its width and height in order to compare with the conspicuous humeral head in other ornithomimosaurs (e.g. Osmólska & Roniewicz, 1970; Osmólska et al., 1972; Kobayashi & Barsbold, 2005a; Sues & Averianov, 2016). Although Pérez-Moreno et al. (1994) proposed that the humeral shaft was mostly straight, its whole curvature is impossible to determine due to the fragmentary condition of the bone. However, in the right humerus (Fig. 13), its proximal end seems to be slightly bowed medially, like in other ornithomimosaurs, such as Sinornithomimus (Kobayashi & Lü, 2003), Gallimimus (MPC-D 100/11), ‘G. mongoliensis’ (FPDM-V-7216) and an ornithomimid from Tajikistan (PIN 3041/2; Alifanov & Averianov, 2006).
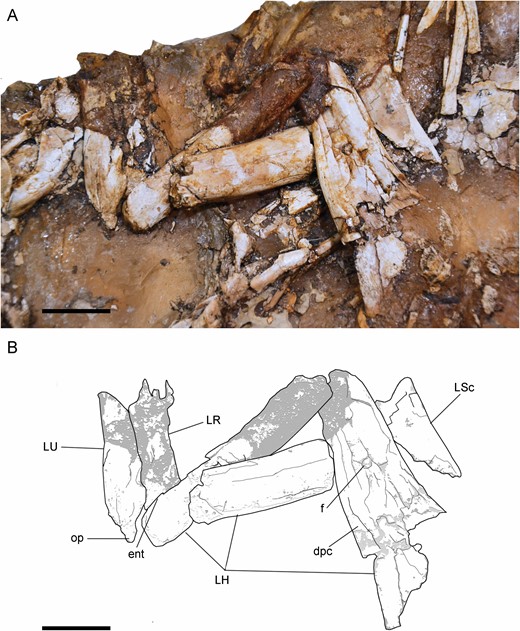
Left forelimb of Pelecanimimus (MCCM-LH 7777) in the left slab: photo (A) and drawing (B). Abbreviations: dpc, deltopectoral crest; ent, entepicondyle; f, foramen; LH, left humerus; LR, left radius; L Sc, left scapula; LU, left ulna; op, olecranon process. Scale bars: 20 mm.
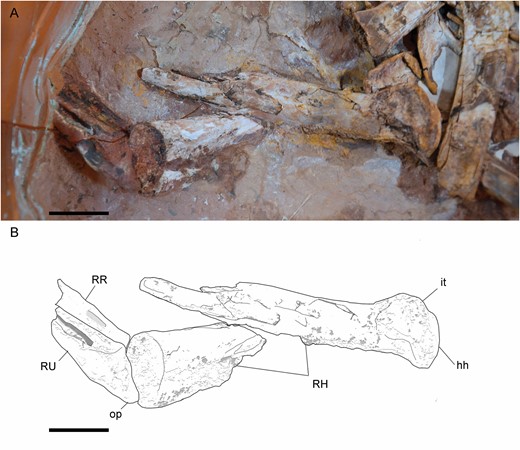
Right forelimb of Pelecanimimus (MCCM-LH 7777) in the right slab: photo (A) and drawing (B). Abbreviations: hh, humeral head; it, internal tuberosity; RH, right humerus; RR, right radius; RU, right ulna; op, olecranon process. Scale bars: 20 mm.
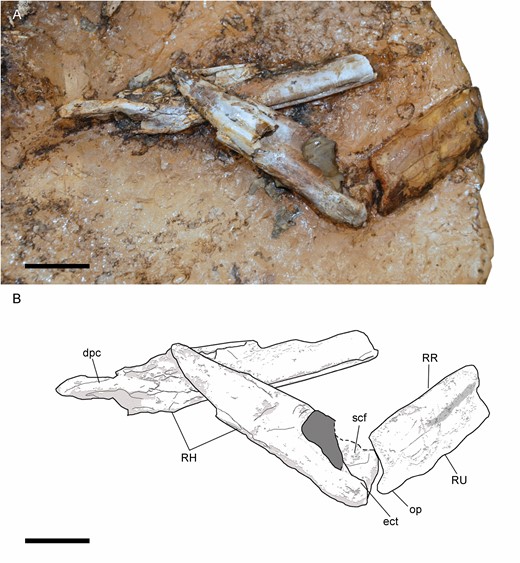
Right forelimb of Pelecanimimus (MCCM-LH 7777) in the left slab: photo (A) and drawing (B). Abbreviations: dpc, deltopectoral crest; ect, ectepicondyle; RH, right humerus; RR, right radius; RU, right ulna; scf, supracondylar fossa; op, olecranon process. Scale bars: 20 mm.
Ulnae and radii:
The shafts of ulna and radius are straight in lateral view (Fig. 12) like in Nqwebasaurus (De Klerk et al., 2000) and, at midlength, slender and long (Fig. 8). The radius rests ventrolaterally on the ulna. The radial shaft is thinner than that of the ulna, like in Harpymimus (Kobayashi & Barsbold, 2005a) or Archaeornithomimus (Smith & Galton, 1990). The olecranon process of the ulna is well developed (Figs 13, 14), like in Harpymimus (MPC-D 100/29; Kobayashi & Barsbold, 2005a), Beishanlong (Makovicky et al., 2009), Struthiomimus (Nicholls & Russell, 1985) and Gallimimus (MPC-D 100/11; Osmólska et al., 1972), but more tapered than in these taxa, like in Nqwebasaurus (Sereno, 2017). Dorsal to the olecranon process, the ulna is strongly curved, unlike the straight surface observed in the ulna of Harpymimus (MPC-D 100/29). The coronoid process is ventrodistally oriented. The proximal shaft of the ulna narrows distally. The surface of the bone is strongly fragmented. The medial surface of the mid-shaft of the ulna has a longitudinal groove. This groove is expanded dorsoventrally on the proximal end.
The distal shaft of the right ulna has a bulge on the dorsal surface (Fig. 15), also observed in ‘Gallimimus mongoliensis’ (FPDM-V-7216). Distally to this bulge on the left ulna (Fig. 8), a dorsal, wide groove is shallowly developed. Distally, both bones are dorsoventrally slightly wide with respect to the rest of the shaft, like in others ornithomimosaurs (e.g. Osmólska & Roniewicz, 1970; Osmólska et al., 1972; Smith & Galton, 1990). The distal end of the ulna is quadrangular in lateral view. The forearm bones are attached to each other along their lengths and strongly joined in the distal and proximal ends, unlike those in Nqwebasaurus (Sereno, 2017). This syndesmosis has been also described in Struthiomimus (Nicholls & Russell, 1985), Beishanlong (Makovicky et al., 2009) and observed in Harpymimus (MPC-D 100/29) and Gallimimus (MPC-D 100/11). In medial view, the ulna has a dorsomedially protruding and rounded distal condyle, separated from the anterior condyle by a shallow groove. This medial condyle is perpendicular to the surface of the shaft, like in Harpymimus (MPC-D 100/29; Kobayashi & Barsbold, 2005a) and Struthiomimus (Nicholls & Russell, 1985), but unlike the semi-oval shape in the distal view of the ulna of Gallimimus (MPC-D 100/11). The radius is dorsoventrally thinner at mid-shaft, although it is broadened at the distal end in lateral view. Proximally to its distal end, the shaft is anteriorly curved, showing a slight anterior bulge.
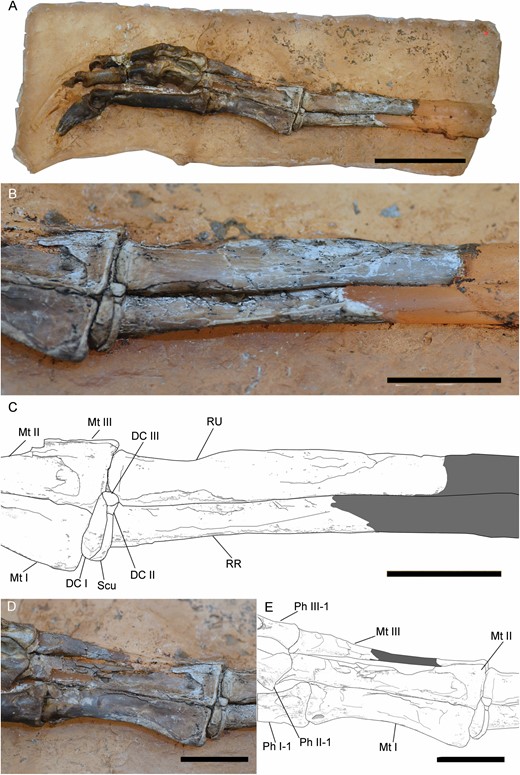
Right forearm and manus of Pelecanimimus (MCCM-LH 7777). Complete manus (A), close-up of ulna, radius and carpals in lateral view (B), drawing (C), close-up of metacarpals in dorsal view (D) and drawing (E). Abbreviations: DC, distal carpal; Mt, metacarpal; Ph, phalanx; RR, right radius; RU, right ulna; Scu, scapholunare. Scale bars: 20 mm.
Manus:
The right manus is positioned in dorsal view, although the digits are slightly twisted and exposed in dorsomedial view (Fig. 15). The surfaces of bones show several fractures due to collapse during compaction. There are five articulated carpals identified (Pérez-Moreno et al., 1994) (Figs 8, 15). The scapholunare [sensuBotelho et al. (2014) for Huaxiagnathus] is triangular and is located between the contact of the ulna and radius, like in Struthiomimus (Nicholls & Russell, 1985). The radiale is proximodistally thin and disc-like. The distal carpals are proximodistally compressed, although the medial distal carpal is larger and thicker than the lateral one, like in Nqwebasaurus (De Klerk et al., 2000). This medial distal carpal is in contact with the radius and the proximal end of metacarpal I. The lateral distal carpal is in contact with the ulna and the medial half of the proximal surface of metacarpal II.
Metacarpal I is the shortest (Fig. 15), but is relatively long with respect to metacarpal II and relatively longer than it is in other basal ornithomimosaurs, such as Nqwebasaurus (De Klerk et al., 2000) and Harpymimus (MPC-D 100/29; Kobayashi & Barsbold, 2005a). The metacarpals of Pelecanimimus have the proportion 0.81:1:0.98/I:II:III (Pérez-Moreno et al., 1994), which is similar to that of some derived ornithomimosaurs such as Gallimimus (MPC-D 100/11; Chinzorig et al., 2018). Metacarpal III has a similar length to metacarpal II, unlike in Harpymimus (MPC-D 100/29), Anserimimus (MPC-D 100/300) and Deinocheirus (MPC-D 100/18), which have a metacarpal III slightly longer than metacarpal II. Moreover, metacarpal I in Pelecanimimus is slenderer than in any of those taxa. The shaft is not waisted, unlike in Nqwebasaurus (Sereno, 2017). The medial cotyle on the proximal end is rounded and slightly more proximally projected than the lateral one. However, this medial tab is poorly developed compared with that strongly proximally pronounced process, called the proximo-radial process by Gishlick & Gauthier (2007) and observed in Compsognathus (Gishlick & Gauthier, 2007) and Sinosauropteryx (Currie & Chen, 2001). The shaft bulges medially in dorsal view. The distal end has a ginglymoid surface like in other basal ornithomimosaurs (Fig. 15) (e.g. Pérez-Moreno & Sanz, 1995; Kobayashi & Lü, 2003; Kobayashi & Barsbold, 2005a), with an intercondylar groove between the well-developed medial and lateral condyles. This latter condition is distinct from the shallowly developed intercondylar groove and non-ginglymoid condition observed in some derived ornithomimosaurs, such as Anserimimus (MCP-D 100/300; Kobayashi & Lü, 2003) or Ornithomimus velox (Claessens & Loewen, 2016). The medial and lateral articular condyles are evenly developed in size in dorsal view, distinct to those unevenly condyles observed in Nqwebasaurus (De Klerk et al., 2000; Choiniere et al., 2012), Harpymimus (MPC-D 100/29), the Bisskety taxon (Sues & Averianov, 2016), Deinocheirus (MPC-D 100/18; Osmólska & Roniewicz, 1970), Ornithomimus velox (Claessens & Loewen, 2016), Anserimimus (MPC-D 100/300) and Gallimimus (MPC-D 100/11; Osmólska et al., 1972). The medial ligamental pit is deeply excavated and rounded. The distal end is wider than the shaft (Fig. 15). In the dorsal surface, there is a proximal groove, bordered by a medial and slightly developed ridge. Metacarpals I and II are appressed along their full lengths. Therefore, metacarpal I is poorly divergent from metacarpal II, a condition similar to that of Anserimimus (MPC-D 100/300; Chinzorig et al., 2018), Ornithomimus edmontonicus (Sternberg, 1933) and Harpymimus (MPC-D 100/29), and unlike that of ‘Gallimimus mongoliensis’ (FPDM-V-7216) and Gallimimus bullatus (MPC-D 100/11), Ornithomimus velox (Claessens & Loewen, 2016) or Deinocheirus (MPC-D 100/18; Chinzorig et al., 2018).
Metacarpal II is extremely wide and triangular dorsally in its proximal end (Fig. 15), but is much thinner at the distal one. The distal end forms a modest ginglymus (Figs 15, 16), distinct from the rounded one observed in the Bissekty taxon (Sues & Averianov, 2016). The articular surface of the distal end is not strongly developed dorsally as in Harpymimus (MPC-D 100/29). The extensor pit is not strongly developed in dorsal view, unlike the condition described in Nqwebasaurus (Sereno, 2017). The shaft is straight in dorsal view.
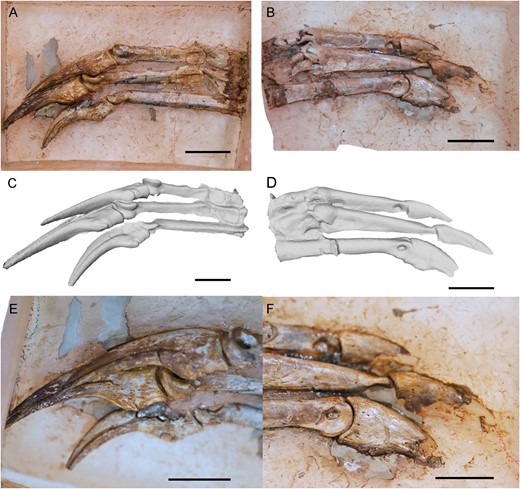
Left manus of Pelecanimimus (MCCM-LH 7777). A, C, E, in ventral view; B, D, F in dorsal view. A, B, photos; C, D, 3D reconstruction; E, F, close-up of ungual phalanges. Scale bars: 20 mm.
Metacarpal III is the slenderest (Fig. 15). The proximal and distal ends are slightly broadened with respect to the mid-shaft. The proximal end is triangular in dorsal view, and the medial side is straight in dorsal view where it contacts metacarpal II. The distal end is rounded (Figs 15, 16), without ginglymoid articulations and with a well-developed articular condyle, like in other ornithomimosaurs (e.g. MCPD 100/29; Kobayashi & Barsbold, 2005a; Makovicky et al., 2009; Sues & Averianov, 2016). The extensor pit is strongly excavated compared with the other metacarpals. Metacarpal III is proximally more medially widened in ventral surface than in its dorsal surface, overlapping with the ventral surface of metacarpal II (Fig. 16A, C).
In general, the manus of Pelecanimimus are long, with specialized, elongated distal phalanges in all digits like other ornithomimosaurs (Makovicky et al., 2004). All phalanges are slender and proximodistally straight (Figs 15, 16). Phalanx I-1 is the longest and most slender in the manus (Figs 15, 16). The shaft of phalanx I-1 is almost rounded in cross-section, but has a ventral flat surface at proximalmost midlength (Fig. 16A). In medial view, the shaft of the phalanx is straight (Fig. 15). The proximal end is dorsoventrally wider with respect to the rest of the phalanx. The medial ligament pit on the distal end is rounded and deeply excavated (Fig. 15A), as is the lateral one (Fig. 16B, D). The distal condyles are thin and marked as parallel ridges (Figs 15, 16A, C). In ventral view, these distal condyles are lateromedially narrow and rounded, separated by a pronounced flexor pit.
The ungual phalanx I-2 has a lateral vascular groove slightly shallow and less proximally extended compared to the other ungual phalanges (Fig. 16A, C, E). The flexor tubercle is well developed and as dorsoventrally thick as in Harpymimus (MPC-D 100/29; Choiniere et al., 2012) and Shenzhousaurus (NGMC 97-4-002), and unlike the poorly developed one observed in Gallimimus (MPC-D 100/11). This flexor tubercle is slightly distally situated respect to the proximal articulation like in other ornithomimosaurs (MPC-D 100/29; Ji et al., 2003; Kobayashi & Barsbold, 2005a; Choiniere et al., 2012; Chinzorig et al., 2018). This ungual phalanx is slightly more curved than those of digits II and III, although it is not as strongly curved as the ungual phalanx I-2 of Nqwebasaurus and Beishanlong (Choiniere et al., 2012), Harpymimus (MPC-D 100/29; Kobayashi & Barsbold, 2005a) or like the strongly curved ungual phalanges of Gallimimus (MCPD 100/11) and Deinocheirus (MPC-D 100/18; Chinzorig et al., 2018).
Phalanx II-1 is proximodistally short and robust compared with phalanx II-2, which is long and slender (Figs 15A, 16A–D). The shaft is dorsoventrally taller than lateromedially thick. The proximal end is strongly ginglymoid. The distal condyles are strongly distally projected and well developed (Fig. 15). The medial ligament pit is barely excavated. This phalanx has paired flexor processes on the ventral surface of the proximal end (Fig. 16A, C). Phalanx II-2 is more than twice as long as the preceding phalanx. The proximal end is ginglymoid (Figs 15, 16A, C). The shaft also is dorsoventrally taller than lateromedial thick, as the preceding phalanx. The medial ligament pit is strongly excavated and rounded (Fig. 15), like in the other phalanges. The flexor tubercle is preserved in ungual phalanx II-3 and it is slightly developed, like that observed in Beishanlong (Makovicky et al., 2009). The ungual phalanges of digits II and III are almost straight along their ventral surfaces (Fig. 16A, C, E), as in other ornithomimosaurs (e.g. Nicholls & Russell, 1985; Smith & Galton, 1990; Choiniere et al., 2012; Sues & Averianov, 2016), but not as much as Anserimimus (MPC-D 100/300; Chinzorig et al., 2018), and unlike the curved ungual phalanges of Harpymimus (MPC-D 100/29), Gallimimus (MCPD 100/11) and Deinocheirus (MPC-D 100/18; Chinzorig et al., 2018). The lateral vascular groove is deeper than in phalanx I in these two unguals (Fig. 16E). The flexor tubercles of III-4 also are well developed compared with other ornithomimosaurs, but these tubercles are still small compared with non-ornithomimid theropods.
The two proximalmost phalanges of digit III are shorter than the distal phalanx III-3 (Fig. 15). Both proximal ones have the same proportions, dorsoventrally high and lateromedially robust. Phalanx III-1 has distal condyles that are mediolaterally thin, compared with the condyles of the other two phalanges (Fig. 15). The proximal articulation is ginglymoid in all the phalanges of digit III. Phalanx III-3 is longer than the sum of the lengths of both preceding phalanges, like in derived ornithomimids (Barsbold & Osmólska, 1990) and Shenzhousaurus (Ji et al., 2003). As in other distal phalanges, the medial ligamental pit is notably marked. Ventrally, the proximal articulations are slightly concave in phalanges II-2 and III-3, and are bordered laterally by a protruded ridge.
RESULTS
A total of 43 postcranial skeletal characters of Pelecanimimus were recoded, based on this new detailed study, in the matrix of Lee et al. (2014). Most of them were originally coded as uncertain ‘?’ by Lee et al. (2014), but can be coded in this study based on first-hand revision of the original material. The heuristic search in TNT resulted in four MPTs of 2953 steps (CI = 0.227; RI = 0.595). The strict consensus configuration is similar to that found in Lee et al. (2014) (Fig. 17; and see Supporting Information, Appendix S1). Pelecanimimus still occupies a basal position within Ornithomimosauria, being more derived than Nqwebasaurus and more basal than Shenzhousaurus. The Bremer support values are similar to those obtained in the original matrix, with a higher value at the Ornithomimosauria node (Bremer index of 5). However, most nodes within the clade are still weakly supported (Bremer values of 1 and 2).
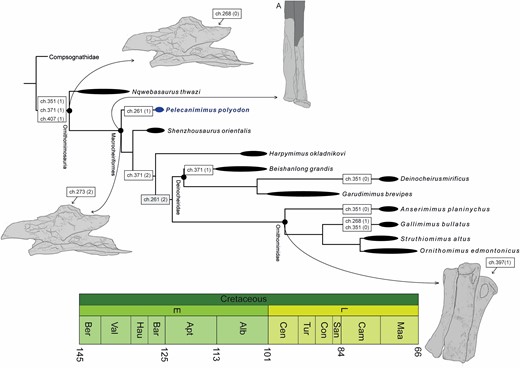
Phylogenetic relationships of Ornithomimosauria and synapomorphies map. Strict consensus topology of four most parsimonious trees recovered from the cladistic analysis. Synapomorphies are indicated in boxes in nodes, with character states in brackets. A, is the syndesmosis in ulna–radius. Terminal nodes/taxa are time-callibrated (thick lines), but more inclusive nodes are not.
Character mapping of the updated characters shows distinct character reconstructions with respect to the results of the previous matrix (Lee et al., 2014) (Fig. 17). The ventral surfaces of anterior cervical centra with a groove (ch. 261 = 2) was the character state defined in derived ornithomimosaurs, unknown in basal members of the clade. Pelecanimimus shows a ventral surface with the intermediate state (ch. 261 = 1), as with most other averostrans. The epipophyses of Pelecanimimus also are considered poorly developed (ch. 268 = 0), as in other ornithomimosaurs (proposed as a synapomorphy of the group; Choiniere et al., 2012), and not well developed as proposed by Lee et al. (2014). Lee et al. (2014) proposed that a strongly reduced cervical neural spine (ch. 273 = 2) was present in most ornithomimosaurs, except for Nqwebasaurus, which presents the intermediate state, but with an unknown coding for Pelecanimimus. The new coding of Pelecanimimus (ch. 273 = 2) establishes that the derived state is present in all ornithomimosaurs, except in the basal-most Nqwebasaurus (the clade here named Macrocheiriformes). Like other basal ornithomimosaurs, such as Shenzhousaurus and Harpymimus (and also some alvarezsaurids and compsognathids), Pelecanimimus has posterior dorsal vertebrae centra that are significantly elongated, and much longer than tall (ch. 282 = 2). Deinocheirus and Garudimimus show a reversed state to the intermediate condition of relatively short dorsal vertebrae centra. The fan-shape of the posterior dorsal neural spines (ch. 293 = 1) is observed in Pelecanimimus and compsognathids (see Discussion below). However, Lee et al. (2014) also coded this as derived in Deinocheirus, although the hypertrophied elongation of its vertebrae is distinct to the morphology observed in Pelecanimimus. The articular facet for the coracoid on the sternum is more anterolateral than anterior in orientation in Pelecanimimus (ch. 345 = 0), unlike as coded by Lee et al. (2014). This orientation is similar to other ancestral sterna, unlike the derived condition in oviraptorosaurians and Paraves. However, it is not possible to establish the condition in the rest of the group due to the absence of sternal plates. One of the synapomorphies proposed for ornithomimosaurians is the long and well-developed coracoid tubercle formed as a ridge (see Discussion below), which is visible in the right coracoid of Pelecanimimus and coded here as having the derived condition (ch. 351 = 1). However, Deinocheirus, Anserimimus and Gallimimus present a reversal of the character state to a mound-like coracoid tubercle. New codings also reflect that Pelecanimimus has an intermediate condition in the transverse width of the distal humerus (ch. 371 = 1), a character only shared with the most basal ornithomimosaurs, Nqwebasaurus and Beishanlong, unlike other ornithomimosaurs that have the derived state of a narrow distal end. Contrary to the coding of the former matrix, Pelecanimimus has been recoded to the ancestral state in characters 397 and 399, presenting a ginglymoid articulation in metacarpal I and metacarpal II. This ancestral condition is unlike the rounded articulation present in those of Ornithomimidae. Another ancestral condition not observed in some Ornithomimidae is the weak curvature of the manual unguals of Pelecanimimus, similar to other basal ornithomimosaurs (ch. 419 = 1) (see Discussion below). Future studies on the cranial osteology of Pelecanimimus and a revision of ornithomimosaur matrices are in progress.
Discussion
Remarks on Ornithomimosauria
In the first description of Pelecanimimus (Pérez-Moreno et al., 1994), several features were compared to those of other ornithomimosaurian taxa, and some potential synapomorphies of the group were suggested. However, given the increase in information available about the group, a review of these comparisons, based on the current record, is necessary. On the axial skeleton, Pérez-Moreno et al. (1994) suggested that Pelecanimimus lacked pneumatic foramina in cervical and dorsal vertebrae, as is usual in Ornithomimosauria. However, Pelecanimimus actually has pneumatic foramina in all cervical vertebrae and the first three dorsal vertebrae, like some ornithomimosaurs that also have pneumatic foramina in the presacral vertebrae, both cervical and dorsal ones (Kobayashi & Barsbold, 2005a; Choiniere et al., 2012; Lee et al., 2014; Sereno, 2017). The presence of pneumatic foramina among the group varies: for instance, Nqwebasaurus has foramina only in its cervical vertebrae (Sereno, 2017), but lacks them in the dorsal series (Choiniere et al., 2012); Deinocheirus also has a peculiar distribution, having pneumatic foramina in all dorsal vertebrae (Lee et al., 2014), not only limited to the the first dorsal centrum, as in Harpymimus (Kobayashi & Barsbold, 2005a), or in the anteriormost two dorsal vertebrae, as Gallimimus (Osmólska et al., 1972).
Pérez-Moreno et al. (1994) noticed two ornithomimosaurian features in the coracoids of Pelecanimimus: a conspicuous coracoid tubercle and a large posteroventral process (Nicholls & Russell, 1985, Makovicky et al., 2004). Only the first one has been maintained as a synapomorphy supporting the monophyly of Ornithomimosauria by Choiniere et al. (2012) and Lee et al. (2014), although an additional character state observed in Ornithomimosauria was added: the development of the coracoid tubercle as a ridge. This state is not exclusive of this group, as it is present also in coelophysoids (Carrano et al., 2012) and non-coelurosaurian tetanurans, except Megalosauria (Benson et al., 2010; Carrano et al., 2012). Another feature shared by Pelecanimimus and ornithomimosaurs is the dorsoventrally low coracoid, proposed as synapomorphy of Struthiomimus and ornithomimids by Nicholls & Russell (1985), and suggested as present in all ornithomimosaurs, except Gallimimus (Osmolska et al., 1972), by Makovicky et al. (2004). Although the specimen MPD 100/14 (cast FPDM-V-7216), informally called ‘Gallimimus mongoliensis’ (Kobayashi & Barsbold, 2006), has a dorsoventrally lower coracoid than G. bullatus (MPC-D 100/11). However, Kobayashi & Barsbold (2006) have suggested that ‘G. mongoliensis’ belongs to a different genus than Gallimimus. In the phylogenetic results of Choiniere et al. (2010), the overall low coracoid is proposed as a feature also shared by Pelecanimimus and alvarezsauroids.
The humeral shaft of Pelecanimimus was suggested as being straight by Pérez-Moreno et al. (1994) and, subsequently, also by Choiniere et al. (2012). The latter authors also have suggested that this is a derived condition in Ornithomimosauria that is absent in Nqwebasaurus and Beishanlong. However, most ornithomimosaurs have a medially bowed humeral shaft (Sereno, 2017), which is strongly bowed in Beishanlong (Makovicky et al., 2009). Therefore, the derived condition would be a medially bowed humerus, contrary to the ancestral condition in Nqwebasaurus that is posteriorly bowed (Sereno, 2017). In Pelecanimimus, the preservation of the humerus makes it difficult to determine the curvature of the shaft, but some fragments of the proximal ends seem to be more (albeit slightly) medially bowed than previously thought (Fig. 18; Supporting Information, Appendix S2).
Digital reconstruction of the left forelimb of Pelecanimimus (MCCM-LH 7777). Reconstructed from broken elements of the left (red) and mirrored right (blue) bones of LH-7777, with hypothetical reconstructed bone in grey, in left lateral (A), anterior (B), medial (C) and posterior (D) views. Notice the slight medial curvature of the humerus in the proximal humerus fragment in anterior view (B). Scale bar: 100 mm.
The tightly adhered distal articulation of the ulna and radius [proposed as a syndesmosis by Nicholls & Russell (1985)] is considered a synapomorphy of the group (Makovicky et al., 2004). This feature is observed in all ornithomimosaurs (Nicholls & Russell, 1985; Makovicky et al., 2010; MPC-D 100/29; MPC-D 100/11), including Pelecanimimus (clade named here Macrocheiriformes) but not in Nqwebasaurus, which has disarticulated ulnae and radii (Choiniere et al., 2012; Sereno, 2017). This latter taxon also has a strongly interosseous space between the bones in the forearm (Sereno, 2017), which is unlike the condition of appressed shafts observed in the rest of ornithomimosaurs. Although Beishanlong also has an interosseous space, the distal end of the ulna and radius also present syndesmosis in the articulation (Makovicky et al., 2009), as does Deinocheirus (MPC-D 100/18). This syndesmosis was proposed as a diagnostics character of Pelecanimimus in the first description (Pérez-Moreno et al., 1994), but its presence in most ornithomimosaurs indicates that it likely is a synapomorphy of a larger group.
Choiniere et al. (2012) proposed, based on a cladistic analysis, that poorly developed epipophyses is a diagnostic character of Ornithomimosauria, also present in other taxa such as some abelisaurids and alvarezsauroids. Pelecanimimus shares this condition with the rest of ornithomimosaurs, with poorly developed epipophyses only in the anteriormost cervical vertebrae. However, in general, the epipophyses observed in Pelecanimimus are more raised than the faintly marked ones in the rest of the Ornithomimosauria taxa, including the most basal taxon, Nqwebasaurus (Choiniere et al., 2012). Choiniere et al. (2012) also suggested that a small flexor tubercle on manual ungual phalanges is a synapomorphy of the group, although it is also known in other coelurosaurians, such as Compsognathus, Alvarezsauridae and EpidexipteryxZhang et al., 2008. The flexor tubercles of Pelecanimimus were described as poorly developed (Pérez-Moreno et al., 1994; Pérez-Moreno & Sanz, 1995), but the current comparison with other ornithomimosaurs shows high variability of the size of flexor tubercles within the clade, with those of Pelecanimimus better developed than in other ornithomimosaurs. Chinzorig et al. (2018) proposed that the degree of development of this tubercle is disparate in the group, related to functional diversity. Another synapomorphy proposed in the phylogenetic analysis of Choiniere et al. (2012) is the presence of paired flexor processes on the ventral surface of the proximal ends of proximalmost manual phalanges (Sereno, 2001). However, this is not exclusive to this group, as TanycolagreusCarpenter et al., 2005 and Alvarezsauridae also present these paired flexor tubercles (Choiniere et al., 2012). This feature is present in all ornithomimosaurs, even in the most basal taxa, such as Pelecanimimus and Nqwebasaurus (Choiniere et al., 2012; Sereno, 2017).
The postzygapophyses in posterior cervical vertebrae tend to be more posteriorly projected than in anterior cervical vertebrae, in which the centrum extends beyond the postzygapophyses in ornithomimosaurs (e.g. Osmólska et al., 1972; Kobayashi & Lü, 2003; Makovicky et al., 2004; Kobayashi & Barsbold, 2005a, b; Lee et al., 2014; Sues & Averianov, 2016). This condition is also observed in Pelecanimimus.
The new clade Macrocheiriformes has a series of synapomorphies based on this osteological study, which are present in Pelecanimimus: (1) a reduced neural spine of the anterior cervical vertebrae; (2) a medially bowed humeral shaft; (3) a syndesmosis between the ulna and radius distal articulation; and (4) the enlargement of metacarpal I and distal phalanges (see Discussion below).
Pelecanimimus has some traits considered plesiomorphic for ornithomimosaurs such as: (1) a shorter and higher axial centrum, different from the enlarged centrum observed in ornithomimids (Kobayashi & Barsbold, 2005b); (2) a smooth, ventral surface of the anterior cervical vertebrae, distinct to the presence of a ventral depression in derived ornithomimosaurs (e.g. Lee et al., 2014); (3) horizontally oriented postzygapophyses in posterior cervical vertebrae, unlike the curved condition observed in some derived ornithomimids (Kobayashi & Barsbold, 2005b); (4) a posteriorly located cervical neural spine on the neural arch, unlike ornithomimids (Kobayashi & Lü, 2003); (5) a distal end of the humerus that is two times transversally wider than the shaft [ch. 371 in Lee et al. (2014)]; and (6) a ginglymoid articulation on the distal ends of metacarpals I and II (Pérez-Moreno & Sanz, 1995), different from the rounded distal end present in most derived ornithomimosaurs (Makovicky et al., 2004).
Diagnostic features of Pelecanimimus
In the original description of Pelecanimimus, two diagnostic postcranial features were proposed: (1) ulna and radius tightly adhered distally and (2) length of the metacarpals showing a proportion of 0.81:1:0.98 (Pérez-Moreno et al., 1994). This postcranial diagnosis has been emended here with four new autapomorphies within Ornithomimosauria.
Ulna and radius tightly adhered distally:
Pérez-Moreno et al. (1994) described briefly the ulna and radius, only remarking that both are strongly adhered distally. In the original description, this condition was compared with the syndesmosis described in Struthiomimus altus (Nicholls & Russell, 1985). However, the authors proposed the character as diagnostic of Pelecanimimus. The recent discoveries of new Ornithomimosauria, and the newer descriptions of some historical taxa, show that almost all ornithomimosaurs have a tight adherence in the distal end of the ulna and radius (e.g. Osmólska & Roniewicz, 1970; Osmólska et al., 1972; Smith & Galton, 1990), even those taxa that have a strongly interosseous space between both forearm bones, such as Beishanlong (Makovicky et al., 2009) and Deinocheirus (MPC-D 100/18). Indeed, this character was proposed, after the description of Pelecanimimus, as a synapomorphy of Ornithomimosauria (Makovicky et al., 2004). Therefore, the adhered distal end of ulna and radius will not be considered here as an autapomorphy of Pelecanimimus.
Proportion of the metacarpals’ length:
One of the most notable features of Pelecanimimus is the proportion of the metacarpals. Other basal ornithomimosaurs, such as Nqwebasaurus, Harpymimus and Shenzhousaurus [although this bone is not preserved in the fossil (NGMC 97-4-002), it is infered by Ji et al. (2003) because the digits are articulated and in life position], present a shorter metacarpal I that is around 50% the length of metacarpal II (Kobayashi & Barsbold, 2005a; Choiniere et al., 2012). This latter condition resembles the ancestral theropod condition, observed in outgroups to Ornithomimosauria (Makovicky et al., 2004). However, Pelecanimimus, an ornithomimosaur more basal than either Harpymimus or Shenzhousaurus based on the most recent phylogenetic analyses (Lee et al., 2014; Chinzorig et al., 2017, this study), has the derived condition with a metacarpal I that is subequal to the other two metacarpals. This autapomorphy, proposed by Pérez-Moreno et al. (1994), is considered also as a possible convergence with the condition present in more derived ornithomimosaurs (Makovicky et al., 2004).
Dorsally tall neural spine of the axis with a posterodorsally tapered, posteriorly projected and rounded apex:
A new potential autapomorphy of Pelecanimimus within Ornithomimosauria is related to the striking morphology of the neural spine of the axis. In other ornithomimosaurs, the neural spine of the axis is dorsoventrally low, anteroposteriorly long and with a straight or sligthly rounded distal outline of the apex (Osmolska et al., 1972; Kobayashi & Lü, 2003; Makovicky et al., 2004; Kobayashi & Barsbold, 2005b). On the contrary, Pelecanimimus has a highly developed and triangular neural spine on the axis, dorsally taller than those observed in other ornithomimosaurs, and with a posterodorsally tapered, posteriorly projected and dorsally rounded and convex apex. Rauhut (2003) established two character states for the axial neural spine: the ancestral condition is a sheet-like neural spine (described as anteroposteriorly long and with the dorsal margin rising in a convex slope from the anterior toward the posterior end); and the derived condition is an anteroposteriorly reduced and rod-like neural spine, restricted to the posterior part of the neural arch and rising abruptly in a concave arc. In Compsognathidae, the sister-taxa of Maniraptoriformes and outgroup to Ornithomimosauria based on Lee et al. (2014), the neural spine of the axis is slightly anteroposteriorly shorter than that of Pelecanimimus and mostly rod-like (Currie & Chen, 2001). This latter condition would be considered as derived for tetanurans [based on Rauhut (2003)]. This condition is also shared by some coelurosaurians, such as tyrannosaurids and dromaeosaurids, and some basal tetanurans. Although the axis of the compsognathid Huaxiagnathus (Hwang et al., 2004) also has a posteriorly projected apex, this apex is acutely, and rod-like, tapered, and not rounded like that of Pelecanimimus. The neural spine of the axis is anteroposteriorly long and extended in all the length of the neural arch in ornithomimosaurs as Gallimimus, Garudimimus, Sinornithomimus and even Pelecanimimus, differing in the derived condition proposed by Rauhut (2003). However, the sheet-like neural spine defined by Rauhut (2003) as the ancestral state for basal theropods, as well as for other non-theropod dinosaurs, is associated with a dorsal margin of the axial neural spine developed as a convex slope toward the posterior end (Rauhut, 2003). Therefore, the spine is like a dorsally curved sheet in lateral view, unlike the condition of a straight and long dorsal apex in other ornithomimosaurs, but more similar to the condition of Pelecanimimus. In contrast, the condition of derived ornithomimosaurs could be a new character state regarding the morphology of neural spines of the axis, because it is distinct from the two conditions proposed by Rauhut (2003). The absence and the incompleteness of this element in the most basal ornithomimosaurs, aside from Pelecanimimus, preclude determining if this is an actual autapomorphy of Pelecanimimus, a convergence with basal theropods or if the ancestral condition in theropods (Rauhut, 2003) is present in the basal taxa of Ornithomimosauria.
Fan-shaped neural spine of posteriormost dorsal vertebrae:
In posterior dorsal vertebrae, neural spines are dorsoventrally low and anteroposteriorly flared in the distal end, with anterior and posterior processes, having a fan-shape in Pelecanimimus, unlike the generally rectangular and dorsoventrally taller neural spines in other basal and derived ornithomimosaurs. The morphology of Pelecanimimus is similar to that described as a synapomorphy of compsognathids (Chen et al., 1998). The phylogenetic position of Compsognathidae is still uncertain. Their position has shifted, being considered as the most basal branches within Coelurosauria (Rauhut, 2003; Holtz et al., 2004), as the sister-taxa of Maniraptoriformes (e.g. Holtz, 1994; Choiniere et al., 2012; Lee et al., 2014) or as member of Maniraptora (e.g. Gauthier, 1986; Holtz, 1998; Norell et al., 2001; Hwang et al., 2004; Chinzorig et al., 2017). Even so, in all possible scenarios for the position of Compsognathidae, the polarity of the character indicates that the condition shared with Pelecanimimus is derived and that the rest of the ornithomimosaurs have the ancestral condition (Rauhut, 2003). The most parsimonious explanation about the presence of this feature in Pelecanimimus depends on the condition in the most basal ornithomimosaur, Nqwebasaurus, independently of the position of Compsognathidae within Coelurosauria. However, although Nqwebasaurus preserves posterior dorsal vertebrae, their preservation precludes observing their neural spines (Sereno, 2017), hampering the knowledge of the evolution of this character. In case the condition of Nqwebasaurus was ancestral, as in other ornithomimosaurians (non-fan-shape), it would be most parsimoniously described as a convergence between compsognathids and Pelecanimimus. If the condition of Nqwebasaurus is the derived condition (fan-shaped), convergence and reversal would be described as equally parsimonious between compsognathids and the early branch of ornithomimosaurs (Pelecanimimus and Nqwebasaurus).
Evenly developed distal condyles in metacarpal I and poor divergence from metacarpal II:
There are two character states in metacarpal I of Pelecanimimus that are unique: (1) evenly developed distal condyles and (2) a poor, almost absent, divergence from metacarpal II. In general, the distal condyles of metacarpal I of ornithomimosaurs are unevenly developed, the lateral condyle being larger than the medial one (Chinzorig et al., 2018) in dorsal and distal views. Although the distal view of the metacarpals of Pelecanimimus is inaccessible, the dorsal view of the right metacarpals allows us to assess that both condyles are equally developed. Although this character could not be exclusive of Pelecanimimus, since two possible new taxa from the Nemegt Formation in Mongolia have evenly developed condyles in the metacarpal I in distal view (Chinzorig et al., 2018). Both taxa have metacarpals I that are strongly and moderately divergent medially compared with the lateral divergence of metacarpal III (Chinzorig et al., 2018). Other ornithomimosaurs also have a less medially divergent first metacarpal, similar to that observed in Pelecanimimus, which has metacarpals I and II contacting each other through all their length. However, these ornithomimosaurs, such as Anserimimus (MPC-D 100/300; Chinzorig et al., 2018), Ornithomimus edmontonicus (Sternberg, 1933) or Harpymimus (MPC-D 100/29), also have unevenly developed distal condyles, especially notable in Harpymimus (MPC-D 100/29).
The new clade Macrocheiriformes
Ornithomimosauria was first erected by Barsbold (1976) as the monophyletic group comprising Garudimimidae, Harpymimidae and Ornithomimidae. Posteriorly, the clade was given a stem-based definition by Sereno (1998) as all maniraptoriforms closer to Ornithomimus than to Neornithes. However, this definition overlaps with that of Holtz (1996) of Arctometatarsalia and also encompassed Therizinosauroidea and Alvarezsauridae, whose relationship to Ornithomimosauria is not supported in recent phylogenies (e.g. Choiniere et al., 2012; Lee et al., 2014; Tsogtbaatar et al., 2017). Therefore, Padian et al. (1999) redefined Ornithomimosauria as the node-based taxon that includes Pelecanimimus and Ornithomimus and all the descendants of their most recent common ancestor. This definition overlaps with that proposed here for Macrocheiriformes. However, Padian et al.’s (1999) definition of Ornithomimosauria excludes the discovery of earlier-branched ornithomimosaurs, such as Nqwebasaurus (De Klerk et al., 2000; Choiniere et al., 2012) and other taxa more related to ornithomimosaurs than to other theropods that branched earlier than Pelecanimimus. For example, several phylogenies (Choiniere et al., 2013, 2014) have retrieved Harpymimus and Shenzhousaurus branching earlier than Pelecanimimus. To secure the inclusion of Nqwebasaurus and other early-branched taxa, Lee et al. (2014) emended Sereno’s (1998) definition of Ornithomimosauria as Ornithomimus velox Marsh, 1890 and all taxa sharing a more recent common ancestor with it than with Allosaurus fragilis Marsh, 1877, Tyrannosaurus rexOsborn, 1905, Compsognathus longipes Wagner, 1859, Alvarezsaurus calvoiBonaparte, 1991, Therizinosaurus cheloniformisMaleev, 1954, Deinonychus antirrhopusOstrom, 1969, Troodon formosuskLeidy, 1856 and Passer domesticus (Linnaeus, 1758), thus excluding taxa originally included by Holtz (1996) and Sereno (1998). Considering the unstable phylogenetic context of early-branching coelurosaurs resulting in complex stem-based definitions, we propose the use of a node-based definition of Ornithomimosauria, such as Nqwebasaurus and Ornithomimus and all the descendants of their most recent common ancestor. This definition of the clade would be compatible with the results of most recent phylogenetic analyses. Therefore, macrocheiriforms would be a less inclusive clade that highlights the relationship between Pelecanimimus + derived ornithomimosaurs (Deinocheiridae + Ornithomimidae) that have been recovered frequently by several authors (e.g. Makovicky et al., 2004; Choiniere et al., 2010, 2012, 2013, 2014; Lee et al., 2014; Tsogtbaatar et al., 2017).
Sternal and uncinate structures of Pelecanimimus
Preserved sterna are rare in the fossil record of theropods, probably due to their fragility, partly due to poor ossification or the weakness of the articulation with the pectoral girdle (Romer, 1956; Holtz et al., 2004; Bradley et al., 2020). Therefore, the knowledge of the evolutionary patterns of the non-avian theropod sternum is patchy, poorly informed and complicated. In the case of Ornithomimosauria, the only known ossified sternum is that of Pelecanimimus, making a comparison with other members of the clade impossible. Although the presence of a sternum is also suggested in Struthiomimus (Nicholls & Russell, 1981) due to a gap between the coracoids and the gastralia, there is no evidence of the ossification of this element in any specimen. It is worth noting that even articulated specimens of ornithomimosaurs with well-preserved pectoral regions, such as Sinornithomimus (Kobayashi & Lü, 2003) or Ornithomimus edmontonicus (RTMP 1995.110.1), lack preserved sternal plates, indicating that this absence could not be a matter of preservation. There are a few specimens of non-avian maniraptoriforms that also preserve the sternum, such as some oviraptorosaurs (Clark et al., 1999; Balanoff & Norell, 2012; Wei et al., 2013; Funston & Currie, 2016; Norell et al., 2018), dromaeosaurids (Norell & Makovicky, 1997, 1999; Hwang et al., 2002; O’Connor et al., 2015) and alvarezsauroids (Perle et al., 1994; Alifanov & Barsbold, 2009).
The morphology of the sternum of Pelecanimimus has the conservative structure of other non-avian maniptoriforms and basal birds (O’Connor et al., 2015), formed by a single pair of bilateral plates that articulate along a midline, with a plate-like overall shape. This morphology is also observed in the recently discovered sternum of the basal non-maniraptoriform theropod Tawa hallae (Bradley et al., 2020). Although Bradley et al. (2020) affirm that costal processes are confirmed only in Tawa and Aves, costal processes are present commonly in many non-avian maniraptoriforms, such as Pelecanimimus, several species of MicroraptorXu et al., 2000 (Hwang et al., 2002; Gong et al., 2012), BambiraptorBurnham et al. 2000 (Burnham et al., 2004) or other dromeosaurids (Godfrey & Currie, 2004). The morphology of oviraptorid sterna is slightly distinct from other non-avian maniraptoriforms, such as Pelecanimimus: the sterna have strongly projected lateral processes, the costal margin being intensely concave and lacking costal processes (Clark et al., 1999; Balanoff & Norell, 2012; Xuefang et al., 2013; Funston & Currie, 2016; Norell et al., 2018). Alvarezsauroidea is a problematic clade within Coelurosauria, but recent discoveries and phylogenies (e.g. Choiniere et al., 2010; Lee et al., 2014; Xu et al., 2018) seem to indicate a consensus of its basal position among Maniraptora (Rauhut & Foth, 2020). Alvarezsauroidea have a striking sternum that differentiates them from other non-avian maniraptoriforms: fused, unpaired and with a ventral keel. In non-maniraptoriform theropods, the sternal record is patchy, and lacks the morphology general in other non-avian theropod sterna (O’Connor et al., 2015), including Pelecanimimus, except in the case of Tawa hallae (Bradley et al., 2020). For instance, those described in some Tyrannosauroidea (Lambe, 1917) have been interpreted later as the possible central portion of the conjoined most anterior pair of gastralia (Brochu, 2003). The sternum in SinraptorCurrie & Zhao, 1993 is unpaired, asymmetrical and irregular (Currie & Zhao, 1993). Moreover, it presents a midline ventral keel (Currie & Zhao, 1993), although distinct to those of alvarezsauroids and not present in any other non-avian theropod sternum. The presence of two pair of ribs and similarity with that of GorgosaurusLambe, 1914 (Lambe, 1917) could indicate that this is also a fused gastralia, as proposed by Brochu (2003) and Bradley et al. (2020). XuanhanosaurusDong, 1984 also seems to have a sternum (Dong, 1984), but it still remains in the sediment and the structure is featureless (Dong, 1984). The sternal plate of CarnotaurusBonaparte, 1985 (Bonaparte et al., 1990) also has a distinct morphology with respect to other sterna, and is more similar to those in tyrannosauroids and Sinraptor, hence, Bradley et al. (2020) also suggest that these sternal plates are likely to be gastralia.
Although there is uncertainty regarding the homology of the uncinate processes of Pelecanimimus due to its preservational state, the morphology of these structures suggests that it is possible that they are actually uncinate processes and, if it is corroborated, their presence in an ornithomimosaur is worth noting in the current phylogenetic context. In theropods, uncinate processes have been only reported in maniraptorans, such as Velociraptor (e.g. Codd, 2004), Microraptor (Xu et al., 2003), Deinonychus (Codd, 2004; Codd et al., 2008; ‘probable ventral ribs’ in Ostrom, 1969), some Oviraptoridae (Clark et al., 1999; Codd, 2004; Codd et al., 2008) and basal and extant birds (Codd, 2004). The record of these elements is patchy in non-avian maniraptoran theropods because the uncinate processes have cartilaginous attachments to the ribs (Codd, 2004). The only evidence of uncinate processes outside Maniraptora are so far those of Pelecanimimus, described here. The uncinates of Pelecanimimus resemble those described in other non-avian maniraptoriforms, such as Oviraptor philoceratops (Codd et al., 2008), Citipati osmolskae (Clark et al., 1999; Codd, 2004) or Velociraptor mongoliensis (Codd et al., 2008): flat, transversally broad, L-shaped and with bell-shaped ends. In Oviraptor, the morphology of the uncinates varies (Codd et al., 2008), but this is not possible to observe in Pelecanimimus due to the preservation of the possible uncinates. In these taxa, the uncinate processes are large and tapered distally, closely resembled extant flying and diving birds (Codd et al., 2008). However, the distal ends and complete lengths of Pelecanimimus uncinate processes are unknown. The presence of uncinate processes in non-avian maniraptorans is suggested as homologous between them and extant birds (Codd et al., 2008). However, the absence of these processes in basal Maniraptora, such as Therizinosauroidea and Alvarezsauroidea (sensu phylogenetical analyses of: Choiniere et al., 2010; Lee et al., 2014), would suggest that its presence in Ornithomimosauria is a homoplastic condition. However, there are taphonomic and excavation biases, and a re-examination of existing basal maniraptoran specimens looking for evidence of ossified uncinates would be helpful to unravel the evolutionary history of these processes (Codd, 2004).
Manual evolutionary pattern in Ornithomimosauria
The manual morphology of Pelecanimimus is one of the striking features of this taxon, especially regarding the proportions of the metacarpals (Pérez-Moreno et al., 1994; Pérez-Moreno & Sanz, 1995). Considering the basal position of Pelecanimimus (see recent phylogenies in: Lee et al., 2014; Chinzorig et al., 2017; and in this study), it would be expected that metacarpal I was around 50% the length of metacarpal II, the basal condition (Makovicky et al., 2004), as in Nqwebasaurus and Harpymimus (Kobayashi & Barsbold, 2005a; Choiniere et al., 2012; Sereno, 2017), whose proportions are two-thirds or half the length between Mc I and Mc II, respectively. However, metacarpal I of Pelecanimimus is subequal in length to metacarpal II, a condition considered derived in Ornithomimosauria (Makovicky et al., 2004). Although Russell (1972) has suggested that having a metacarpal I and III relatively shorter than metacarpal II was a plesiomorphic character, since it is a usual condition in theropods, Makovicky et al. (2004) remarked that metacarpal I in ornithomimosaurs is longer than in other coelurosaurians and subequal in length to metacarpal II. Therefore, these authors proposed a long metacarpal I as the derived condition in Ornithomimosauria. Nevertheless, the disparity of the manus within Ornithomimosauria is higher than previously thought (Chinzorig et al., 2018). There are several derived taxa that have a longer metacarpal I than metacarpal II, such as the genera Ornithomimus (Russell, 1972; Makovicky et al., 2004; Claessens & Loewen, 2016) and Anserimimus, and some specimens from the Nemegt Formation (Chinzorig et al., 2018). Other derived specimens, such as Gallimimus bullatus (Osmólska et al., 1972; Chinzorig et al., 2018), Archaeornithomimus (Nicholls & Russell, 1981; Smith & Galton, 1990; Chinzorig et al., 2018), Struthiomimus (Nicholls & Russell, 1985), Deinocheirus (Chinzorig et al., 2018) or some new specimens from the Nemegt Formation (Chinzorig et al., 2018), have a metacarpal I almost 80% the length of metacarpal II, the same condition as Pelecanimimus.
In general, the manus of theropods have a tendency toward enlargement relative to the size of the forelimb (Gauthier, 1986), caused by elongation of both metacarpal and manual phalanges. Rauhut (2003) established that the most derived condition in theropods is to have a long phalanx I-1 with respect to metacarpal I (ratio greater than 1.5). However, as also stated by this author, in advanced ornithomimosaurs, metacarpal I is strongly enlarged compared with metacarpal II (Makovicky et al., 2004). Therefore, the ratio in derived ornithomimosaurs is less than the most early-branching condition. This is also the condition observed in Pelecanimimus, with a ratio of 1.375 (see measurements in Table 3), contrary to the condition of the basal Harpymimus that has a ratio of 2.5 [based on measurements of Kobayashi & Barsbold (2005a)] due to its smaller metacarpal I. Deinocheirus also has a high ratio (1.5, based on the measurements of Osmólska & Roniewicz, 1970) due to the smaller phalanx I-1 relative to other ornithomimosaurs (Chinzorig et al., 2018). Due to the disparity observed in the length of metacarpal I in this group (Chinzorig et al., 2018), which affects the coding of this character state as proposed by Rauhut (2003), the length of this phalanx is compared to the humerus length. Pelecanimimus has a slightly smaller phalanx I-1 (ratio 0.34) than Harpymimus [0.41, based on measurements of Kobayashi & Barsbold (2005a)]. Although Gallimimus bullatus has a one of the smallest phalanges I-1 compared with the humerus (Osmólska et al., 1972), this is due to the shorter manus compared with the forelimb that is characteristic of this taxon (Osmólska et al., 1972; Kobayashi & Barsbold, 2006). However, based on the quantitative studies of Chinzorig et al. (2018), Gallimimus has an elongation of phalanx I-1 compared with other phalanges, as is the case with other ornithomimosaurs (Chinzorig et al., 2018). Although most theropods have a longer phalanx II-2 than phalanx II-1 (Rauhut, 2003), the condition in most ornithomimosaurs is having a penultimate phalanx around twice as long as the first phalanx, based on measurements of previous studies (e.g. Sternberg, 1933; Osmólska & Roniewicz, 1970; Osmólska et al., 1972; Nicholls & Russell, 1985; Kobayashi & Barsbold, 2005a, Sereno, 2017). However, Nqwebasaurus has the shortest phalanx II-2 compared with phalanx II-1 within Ornithomimosauria (ratio 1.17). Deinocheirus (Osmóslka & Roniewicz, 1970) also has a small phalanx II-2 (ratio 1.62) relative to the rest of Ornithomimosauria (Chinzorig et al., 2018), with a similar low ratio in Anserimimus (1.40). The latter taxon had a special proportion among ornithomimosaurs in having relatively elongated proximal phalanges instead of elongated distal phalanges (Chinzorig et al., 2018). Pelecanimimus and other more derived ornithomimosaurs, not including Deinocheirus and Anserimimus, have a longer phalanx III-3 than both proximal phalanges taken together, the most derived condition for theropods proposed by Gauthier (1986). This condition is also observed in other theropods, such as Guanlong (Xu et al., 2006), the alvarezsauroid Haplocheirus (Choiniere et al., 2012 coding), most paravian taxa (Rauhut, 2003) and it is an unusual feature observed in the carcharodontosaurid ConcavenatorOrtega et al., 2010 (Cuesta et al., 2018). In general, except for the striking condition of Anserimimus and Deinocheirus (Chinzorig et al., 2018), all ornithomimosaurs more derived than Nqwebasaurus (clade Macrocheiriformes) have strongly elongated distal phalanges in all manual digits, even Gallimimus, which has relatively the shortest manus. Although the relative length of the manus varies in ornithomimosaurs, there is a tendency toward having elongate forelimbs relative to others theropods (Makovicky et al., 2004). Recent studies support that and have suggested that, in spite of a main evolutionary trend of negative forelimb allometry for non-avian Theropoda, some specific clades, such as Coelophysoidea, Ornithomimosauria and Oviraptorosauria, exhibit allometric trends that are closer to isometry in the forelimbs (Palma Liberona et al., 2019). Regarding this forelimb elongation, the manus has also a tendency to a relative enlargement, most conspicuous in this group than in other theropods, especially for metacarpal I (Makovicky et al., 2004), and the strongly elongated distal phalanges, with the exception of the above-mentioned taxa. In Maniraptora there is also a highly allometric trend in the manus in the early stages of their evolution, with the most modified elements being digits I and III, digit II being the least modified (Nebreda et al., 2020). Since the more ancestral hand of Nqwebasaurus has short distal phalanges and a short metacarpal I, this tendency to enlargement of metacarpals and phalanges arises in Macrocheiriformes for the first time, with Pelecanimimus being the most early-branching ornithomimosaur with a long manus with respect to the forelimb. Despite the short metacarpal I of Harpymimus, the enlargement of its phalanges is also appreciable; thus, this is in line with this trend. This derived condition of the group, having a long manus, could vary in some derived taxa to more specialized forms such as Deinocheirus, Gallimimus or Anserimimus.
CONCLUSION
The detailed description and updated character-coding of Pelecanimimus reveal several Ornithomimosauria synapomorphies in its postcranial skeleton: (1) a conspicuous ridge-like coracoid tubercle, (2) a dorsoventrally low coracoid, (3) a tightly adhered distal articulation of the ulna and radius, (4) poorly developed epipophyses in the anteriormost cervical vertebrae, (5) paired flexor processes on the ventral surface of the proximal ends of proximalmost manual phalanges and (6) posterior cervical vertebrae postzygapophyses more posteriorly projected than in anterior cervical vertebrae.
Although the resulting phylogenetic hypotheses are similar to those proposed so far, with an early-branching position of Pelecanimimus within Ornithomimosauria, the detailed description of the taxon shows plesiomorphic characters in Pelecanimimus neither coded nor described in previous datasets and some synapomorphies within Ornithomimosauria. A well-supported node with several shared evolutionary novelties, including an enlargement of the manus, lead to the definition of a new clade, Macrocheiriformes, which includes Pelecanimimus and Ornithomimus but not Nqwebasaurus.
The diagnosis of Pelecanimimus has been emended, with a unique combination of four characters relative to the morphology of the neural spine of the axis, dorsally tall with a tapered and rounded apex, fan-shape of posterior dorsal vertebrae neural spines in lateral view, and the unique morphology and developement of its metacarpals. The striking morphology and proportion of the manus of Pelecanimimus are within the disparity ranges in the manus of ornithomimosaurs. Despite this variability, a tendency toward elongation of the manus in ornithomimosaurs is evidenced in the entire group by metacarpal proportions and the size of distal phalanges. This tendency is less evident in the basal-most member of Ornithomimosauria, Nqwebasaurus. Therefore, the features of the manus of Pelecanimimus reveal that the early specialization of the ornithomimosaur manus arose, so far, in Macrocheiriformes. Pelecanimimus is the only Ornithomimosauria with an ossified sternum. The morphology of the sternum is similar to that of non-avian maniraptoriforms, basal birds and the basal theropod Tawa. Pelecanimimus also shows the first evidence of ossified uncinate processes outside Maniraptora, indicating a convergent appearance of these structures in Ornithomimosauria and the more derived Maniraptora. However, the evolutionary history of both anatomical elements, ossified sternum and uncinate processes, is still unclear due to taphonomic and excavation biases, in particular, and is, therefore, subject to change.
SUPPORTING INFORMATION
Additional Supporting Information may be found in the online version of this article at the publisher’s web-site.
Appendix S1. Phylogenetical information (coding of characters of Pelecanimimus and strict consensus).
Appendix S2. Supplementary Video 1. Reconstructed virtual left forelimb of Pelecanimimus.
Appendix S3. Supplementary Video 1. Reconstructed virtual right sternal plate of Pelecanimimus.
ACKNOWLEDGEMENTS
We acknowledge S. Langreo (MUPA) for providing access to the holotype of Pelecanimimus polyodon. We also thank FPDM staff members; F. Zheng (IVPP, China); L. Zhang (IVPP, China); X. Xu (IVPP, China); T. Chinzorig (MPC, Mongolia); K. Tsogtbaatar (MPC, Mongolia); M. Manabe (NMNS, Japan); Y. Hai-Lu (NGMC, China); and P. Chanthasit (PW, Thailand) for providing access to specimens under their care and K. Miyata (FPDM) for access and assistance in conducting CT scan analyses. We acknowledge T. Hirasawa for his useful comments and discussion about the uncinate process of Pelecanimimus and A. Serrano-Martínez for his helpful comments about the CT techniques. We also thank the editors and two anonymous reviewers, who made constructive suggestions that greatly improved the manuscript. EC research was financed by a Japan Society of Promotion of Science Postdoctoral Research Fellowship (ref. PE18034), a German Academic Exchange Service DAAD Postdoctoral Research Fellowship (ref. 91791394). DV research was financed by predoctoral FPI Universidad Nacional de Educación a Distancia Grant (ref. 0531174813 Y0SC001170). This research also was financed by the project CGL2013-42643 of the Spanish Ministerio de Economía y Competitividad. The authors declare no conflict of interests.
Data archiving statements
The phylogenetic data is included in the Supporting Information, Appendix S1 and the data matrix is available on morphobank (http://morphobank.org/permalink/?P3812). The holotype specimen of Pelecanimimus polyodon (MUPA-LH-7777) and the virtual fossils generated from it are deposited at the Museo de Paleontología de Castilla la Mancha (Cuenca, Spain).
References

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}