





Plants and fungi have established extensive inter-kingdom interactions in the form of mycorrhiza, involving approximately 340,000 plants and more than 50,000 fungal species (Genre et al. 2020). The orchid mycorrhiza (OrM) represents 1 of the 4 major types of mycorrhiza symbiosis and has drawn extensive attention due to the exceptional diversity (>2,700 species) of orchids and the critical roles of OrM in the orchid life cycles (Favre-Godal et al. 2020). Many orchids rely on OrMs to complete seed germination and seedling development (Rasmussen et al. 2015). Knowledge about OrM symbiosis during the propagation stage is of crucial importance for orchid conservation (Bhatti and Thakur 2022), which is particularly relevant because there have been huge declines of orchids around the world due to human-induced habitat destruction and climate changes in recent decades (Phillips et al. 2020).
The dust-like seeds of orchids with a simple embryo have extremely complex life cycle strategies, in part because these tiny seeds have minimal nutrient reserves. The seeds germinate and develop into tuber-like protocorms before they finally form plantlets. Seed germination and protocorm development are crucial stages for establishment of the OrM symbiosis, and sometimes different fungi are recruited in these processes (Selosse et al. 2022).
The complex OrM interactions present particular challenges in orchid conservation and propagation through seed germination, protocorm development, and plantlet formation. Some OrM fungi are compatible to promote both seed development and protocorm development for specific orchid species. However, are there any compatible fungi with broad host spectra to support OrM symbiosis for orchid propagation? New findings presented by Xu et al. (2023) in this issue of Plant Physiology shed new light on OrM symbiosis and its potential application in orchid conservation.
Orchids generally show high specificity of OrM interactions. However, some OrM fungi species, such as Serendipita indica from the Serendipitaceae (Sebacinales), also interact with non-orchid plants and thus provide the possibility for wider compatible OrM symbiosis (Selosse et al. 2022). Xu et al. (2023) tested the compatibility of S. indica in 12 taxonomically different orchid species with diverse life forms from the genera of Acampe, Calanthe, Cymbidium, Dendrobium, and Geodorum. They found that S. indica promoted seed germination and protocorm development of 6 among the 12 orchid species, mainly from the Dendrobium and Cymbidium genera. It is noteworthy that all the 6 species are epiphytic, and none of the terrestrial orchids tested showed compatibility with S. indica.
S. indica sometimes interacts with some plants via a loose and undifferentiated colonization in the roots (named endophytism) (Selosse et al. 2022). The authors next tested whether this fungus promotes seed germination and protocorm development of the 6 orchid species via OrM or endophytism (Xu et al. 2023). Microscopic examination of semi-thin sections of the protocorms revealed toluidine blue O–stained mycelium at the basal regions of the protocorms, and both living and old lysed pelotons (intracellular hyphal coils) were recognized in the colonized cells. These results suggested that OrM symbiosis was established in the basal region of these protocorms.
The well-studied medicinally valuable orchid Dendrobium catenatum, which has an available genome sequence, provides an excellent model to study the underlying mechanisms of S. indica OrM-promoted seed germination and protocorm development. Xu et al. (2023) profiled transcriptomes of aseptic and symbiotic protocorms and identified a total of 356 differentially expressed genes (DEGs). The DEGs were enriched in KEGG pathways related to defense, immune response, and associated metabolism. The core metabolic pathway of glycolysis was especially remarkable because it showed the highest ratio of upregulated versus downregulated DEGs.
A closer scrutiny of the glycolysis pathway revealed that genes encoding alcohol dehydrogenase (ADH) and pyruvate decarboxylase were noticeably upregulated in the symbiotic protocorms in comparison with those aseptic ones. There are 12 homologues of ADH genes in D. catenatum with transcripts from 10 detected in the protocorms. Strikingly, all 10 of these DcADH genes showed higher transcript levels in the symbiotic protocorms compared with those aseptic ones, and 4 (DcADH1, DcADH2, DcADH3, and DcADH7) exhibited more than 2-fold changes. An in situ hybridization assay was performed and showed that DcADH2 was predominantly expressed in the basal region of the symbiotic protocorms, which is key to root development for the plantlet (Xu et al. 2023). RNA levels of DcADH1 and DcADH2 were also found to be closely related to the symbiosis process, suggesting that DcADHs were involved in regulation of the S. indica-D. catenatum symbiosis (Figure 1).
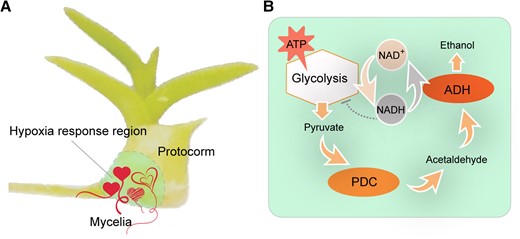
Mycorrhiza-induced hypoxia responses promote seed germination and protocorm development of orchids. A) Symbiosis of the OrM fungi in the basal region of the protocorm. Mycelia of the OrM fungi Serendipita indica induce hypoxia responses in the colonized cells of the protocorm, which is beneficial for orchid seed germination and protocorm development. The “hearts” in the hypoxia response region indicate morphologically various mycelia of the OrM fungi. B) The ADH-mediated working model for hypoxia response in the OrM symbiosis. While generating ATP, the glycolysis metabolic pathway also produces NADH, which in turn inhibits glycolysis. Under low oxygen (hypoxia) conditions, ADH- and pyruvate decarboxylase (PDC)-regulated hypoxia metabolism converts NADH to NAD+, which decreases the NADH accumulation and allows further glycolysis and ATP production for cellular activity. The protocorm cartoon in panel (A) is adapted from Figure 1A of Xu et al. (2023), and the “heart” cartoons are adapted from free vectors of www.vecteezy.com.
ADH and pyruvate decarboxylase are key enzymes for alcoholic fermentation in many plants under hypoxic conditions. Genes encoding the oxygen sensor plant cysteine oxidase also showed upregulated expression in the symbiotic protocorms. These results suggested that the OrM symbiosis by S. indica might induce a hypoxia response in D. catenatum. To test whether DcADH activity is necessary for the OrM symbiosis, Xu et al. (2023) added the ADH inhibitors puerarin and 4-methylpyrazole to the inoculated D. catenatum protocorms. The authors found that S. indica colonization was dramatically reduced in response to the addition of the ADH inhibitors. The most upregulated DcADH genes, DcADH1 and DcADH2, were cloned and expressed in Escherichia coli to analyze their functionality to convert NADH to NAD+. In vitro biochemical assays showed that both DcADH1 and DcADH2 expressed in E. coli were active, and the activity was inhibited when puerarin and 4-methylpyrazole were added. Thus, it is very likely that the hypoxia response–regulated DcADH has a role in the OrM symbiosis.
Taken together, the findings presented by Xu et al. (2023) demonstrate the potential of compatible OrM fungi with broad host spectra in orchid conservation and provided insights into the mechanisms underlying OrM symbiosis. However, it remains unclear how generally S. indica is compatible for orchid species. Larger-scale screening of compatible fungi for OrM symbiosis will bring valuable information for future orchid conservation.
One of their key findings is that the early stages of symbiosis seem to involve the induction of hypoxia in the protocorm, revealed by the expression of several hypoxia-induced genes. Hypoxia might be induced through competition between the plant and fungal cells for oxygen or by increase in plant cell respiration—further studies are needed. The close relationship between the ADH-mediated hypoxia response and OrM symbiosis raises the possibility that the application of hypoxic conditions could promote seed germination and protocorm development for OrM-assisted orchid propagation.
Although oxygen is required in many biological activities, including seed germination, hypoxia is known to promote seed germination of some aquatic plants (Corbineau 2022). The type of seed reserves seems to be crucial for their responses to hypoxia, and starchy seeds are more prone to germination under lower oxygen content than lipid-rich seeds (Corbineau 2022). It remains unclear whether the orchid species compatible with S. indica have starch reserve in their embryos, so further studies on seed reserves of various orchids may help develop better protocols for seed germination and OrM symbiosis in orchid conservation.
Funding
D.C. was supported by the Young Elite Scientists Sponsorship Program (530000221100000144054-4) from the Yunnan Association for Science and Technology.
References
Author notes
Conflict of interest statement. The authors declare no conflict of interest.

{kind=link}