





Plants contribute significantly to the global water cycle, redistributing about 10% of liquid water from the land to the atmosphere and through the different soil layers. The water deficit in the atmosphere pulls the water column from the soil to the leaves, thanks to the cohesion between water molecules (Dixon and Joly, 1894). A negative hydrostatic pressure (or tension) is created by the vaporization of water and its loss through the stomata that propagates downstream towards the root system. During the day, the amount of water supplied by the root system does not compensate for transpired water, creating a water potential gradient between below- and aboveground organs. At night, as stomata close, water re-equilibrates along the water column, and the xylem sap reaches the soil water potential prospected by the root system (Améglio et al., 1999). During the night, water is thus transferred through the soil layers and inside the plant to the lowest water potential (leaf or soil). This phenomenon, known as hydraulic redistribution, generally causes water to rise from the depth of the soil in all directions (Richards and Caldwell, 1987). Different patterns of hydraulic redistribution have been described according to the main direction (vertical through hydraulic lift or descent, or horizontal through lateral horizontal roots) and destination (foliar uptake when leaf water potential is higher than soil water potential or tissue dehydration in relation to its capacitance; Nadezhdina et al., 2010).
By chance, Liu et al. (2021) identified a hydraulic distribution mechanism that consists of bi-directional water flow in the lateral roots of Populus tomentosa. In the upper part of the root, the flow of water through the xylem is centripetal, while in the lower part, the flow is centrifugal. Liu et al. (2021) measured sap flow velocity by the heat-pulse method. A heating element surrounded by two temperature probes was inserted inside the root in a radial direction. The sap velocity was calculated as a function of the time required by the flowing sap to transport heat from the heater to temperature probes. This technique allows detecting sap velocity in both directions. The bidirectional pattern was detected because the temperature probes carry two measuring points (thermistors) at different distances along the root diameter. The sap flow rate was therefore measured at two locations along the root diameter, that is, at the lower and upper part of the root.
To gain further insights into the mechanism driving bidirectional hydraulic distribution, Liu et al. (2021) set up an ambitious experiment combining measurements in 14 roots from five trees under three contrasted irrigation regimes over 2 years. The bidirectional characteristic was only evident in surface roots that were not connected to a vertical root (also called a sinker). Contrary to other hydraulic distribution mechanisms that occur when plant transpiration is very limited or null, bidirectional water flow was only detected during the day when the plants were transpiring. Sap flows in the upper part of the roots were always centripetal and proportional to the sap flow of the stem and to the transpiration of the whole plant. However, sap flow in lower roots exhibited centripetal and centrifugal patterns for low and high soil water content, respectively.
Integrating these experimental results, the authors describe a hydraulic redistribution mechanism called the evaporation-driven hydraulic redistribution (EDHR; Figure 1). This mode of water flow was only observed in the superficial roots limited to one horizontal soil layer under extremely dry superficial soil conditions. The tension created by leaf transpiration was transmitted to the base of the horizontal roots. At this particular location, the tension was transmitted both horizontally to the distal part of the root and vertically along the taproot. Since the lateral roots were surrounded by much drier soil than the taproot, the drop in water potential was greater along the lateral root than vertically, creating an inverse gradient between the area below the junction of the lateral root and the root tip.
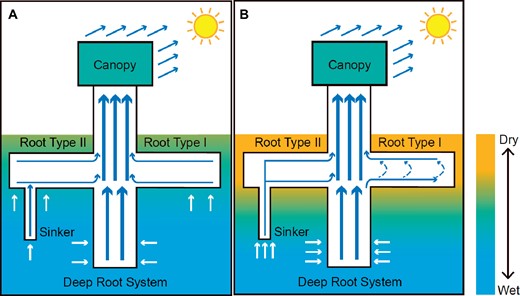
Hydraulic flows in P. tomentosa depending on soil water content (A wet in blue; B dry in yellow). Solid arrows indicate water transport, and dashed arrows indicate the circumferential water transport within the superficial roots limited to one horizontal soil layer when EDHR occurs (adapted from Liu et al., 2021).
EDHR can have profound consequences on the exposure of superficial roots to drought, increasing their lifespan and activity under extremely dry conditions. EDHR has been detected in a significant proportion of superficial roots and could be a part of drought avoidance strategy in species with a high proportion of horizontal lateral roots that experience recurrent drought episodes. This mechanism, contributing to soil moisture conditions, the global water cycle, and soil biological activities, sheds light on the hidden architecture of the root system. Additional work is required to understand the factors that control root system architecture.
Funding
This work has been carried out with the financial support of the Auvergne-Rhône-Alpes regional Council (Doux-Glace) and the Agence Nationale de la Recherche (Acoufollow ANR-20-CE91-0008).
References
Author notes
Senior author.

{kind=link}