





Abstract
We investigated the roles of cortical microtubules in gravity-induced modifications to the development of stem organs by analyzing morphology and orientation of cortical microtubule arrays in hypocotyls of Arabidopsis (Arabidopsis thaliana) tubulin mutants, tua3(D205N), tua4(S178Δ), and tua6(A281T), cultivated under 1g and hypergravity (300g) conditions. Hypocotyls of tubulin mutants were shorter and thicker than the wild type even at 1g, and hypergravity further suppressed elongation and stimulated expansion. The degree of such changes was clearly smaller in tubulin mutants, in particular in tua6. Hypocotyls of tubulin mutants also showed either left-handed or right-handed helical growth at 1g, and the degree of twisting phenotype was intensified under hypergravity conditions, especially in tua6. Hypergravity induced reorientation of cortical microtubules from transverse to longitudinal directions in epidermal cells of wild-type hypocotyls. In tubulin mutants, especially in tua6, the percentage of cells with longitudinal microtubules was high even at 1g, and it was further increased by hypergravity. The twisting phenotype was most obvious at cells 10 to 12 from the top, where reorientation of cortical microtubules from transverse to longitudinal directions occurred. Moreover, the left-handed helical growth mutants (tua3 and tua4) had right-handed microtubule arrays, whereas the right-handed mutant (tua6) had left-handed arrays. There was a close correlation between the alignment angle of epidermal cell files and the alignment of cortical microtubules. Gadolinium ions, blockers of mechanosensitive ion channels (mechanoreceptors), suppressed the twisting phenotype in tubulin mutants under both 1g and 300g conditions. Microtubule arrays in tubulin mutants were oriented more transversely by gadolinium treatment, irrespective of gravity conditions. These results support the hypothesis that cortical microtubules play an essential role in maintenance of normal growth phenotype against the gravitational force, and suggest that mechanoreceptors are involved in modifications to morphology and orientation of microtubule arrays by 1g gravity and hypergravity in tubulin mutants.
The direction of cell expansion is important for determining the shape of whole plant body. Cortical microtubules are assumed to be responsible for anisotropic expansion of plant cells (Wasteneys and Galway, 2003; Lloyd and Chan, 2004; Mathur, 2004; Baskin, 2005; Paredez et al., 2008). The prevailing view is that cortical microtubule arrays direct or constrain the movement of the cellulose synthase complexes and thus align nascent cellulose microfibrils in the same direction in the innermost layer of the cell wall (Baskin, 2001), although some other mechanisms may also be involved (Baskin, 2001; Sugimoto et al., 2003; Wasteneys, 2004).
It is evident that orientation of cortical microtubules plays an essential role in creating the distinct shape of higher plant organs, even if there is uncertainty over the mechanism by which microtubules influence morphogenesis. The importance of cortical microtubule arrays for anisotropic growth has been documented by pharmacological studies and experiments with helical growth mutants of Arabidopsis (Arabidopsis thaliana). Mutants on α- and β-tubulins as well as microtubule-associated proteins show either left-handed or right-handed helical growth (Thitamadee et al., 2002; Nakajima et al., 2004; Sedbrook et al., 2004; Shoji et al., 2004). The rapidly elongating cells of these mutants skew consistently either to the right or to the left and exhibit cortical microtubule arrays that form shallow helices with fixed handedness (Thitamadee et al., 2002; Abe and Hashimoto, 2005; Ishida et al., 2007). Cortical microtubule arrays in the left-handed helical growth mutants form right-handed helix, whereas those in right-handed helical growth mutants form left-handed helix (Thitamadee et al., 2002; Abe and Hashimoto, 2005; Ishida et al., 2007). These results indicate that dysfunctional cortical microtubules are arranged in helical arrays and affect the direction of cell expansion.
The gravitational force is one of the environmental factors that determine the plant body shape. Under hypergravity conditions produced by centrifugation, plants generally have a shorter and thicker body (Soga et al., 2006). Namely, hypergravity modifies growth anisotropy. In Arabidopsis hypocotyls, the expression of most α- and β-tubulin genes was up-regulated by hypergravity (Yoshioka et al., 2003; Matsumoto et al., 2007). In protoplasts of Brassica hypocotyls, hypergravity stimulated the regeneration of cortical microtubules into parallel arrays (Skagen and Iversen, 1999), and in azuki bean (Vigna angularis) epicotyls it increased the percentage of cells with longitudinal cortical microtubules (Soga et al., 2006). The reorientation of cortical microtubules from transverse to longitudinal directions may be involved in modifications by hypergravity to growth anisotropy.
The aim of this study was to clarify the roles of cortical microtubules in gravity-induced modifications to development of stem organs. For this purpose, we examined the changes in growth, morphology, and orientation of cortical microtubule arrays in hypocotyls of Arabidopsis amino acid substitution mutants in α-tubulin structure, tua3, tua4, and tua6, grown under 1g and 300g conditions. We have reported the possible involvement of mechanosensitive ion channels (mechanoreceptors) in hypergravity-induced modifications to growth and cell wall properties (Soga et al., 2004, 2005, 2006). Thus, we also examined the effect of blockers of mechanoreceptors on helical growth and orientation of cortical microtubule arrays in the tubulin mutants.
RESULTS
Effects of Hypergravity on Growth
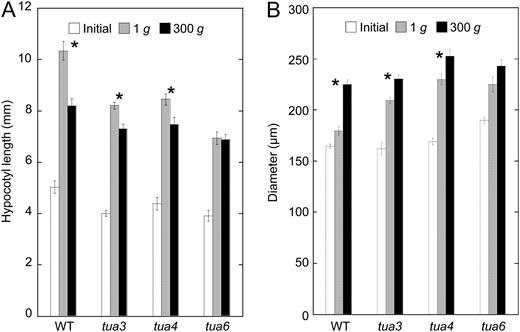
Effects of hypergravity on elongation growth and lateral thickening in hypocotyls of Arabidopsis tubulin mutants. Wild type (WT) and tubulin mutants were grown on agar medium at 1g for 48 h at 25°C. Seedlings were then transferred to 1g or 300g conditions, and grown for a further 24 h at 25°C. A, The length was measured using a scale. B, The diameter of hypocotyls was measured with a digital stereoscopic microscope. Values are means ± se (n = 20). *, Mean values with significant differences between 1g and 300g treatments (P < 0.05).
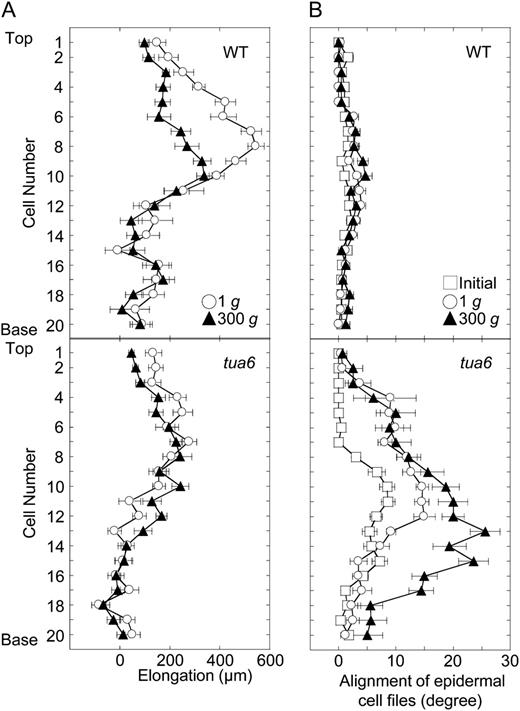
Cell elongation and twisting profiles in an epidermal cell file in hypocotyls of Arabidopsis tubulin mutants. Wild type (WT) and tua6 were grown as in Figure 1. A, The length of individual cells from the top (cell 1) to the base (cell 20) was measured with SEM and elongation for 24 h was calculated. B, The alignment angle of individual cells was measured with SEM. Values are means ± se (n = 15).
Effects of Hypergravity on Cell Alignment
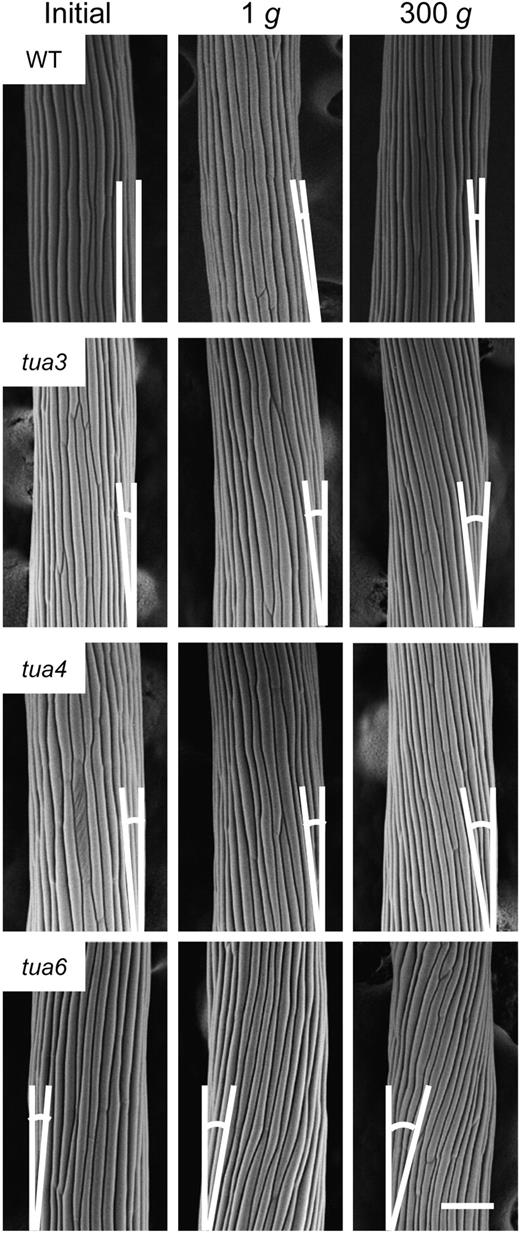
Surface view of epidermal cell files in Arabidopsis tubulin mutants grown at 1g or 300g. Wild type (WT) and tubulin mutants were grown as in Figure 1. The outer surface of cells 10 to 12 from the top of hypocotyls was observed with SEM. Supplementary lines denote the longitudinal axis of hypocotyls and the alignment of epidermal cell files, for measuring the alignment angle. The bar denotes 100 μm.
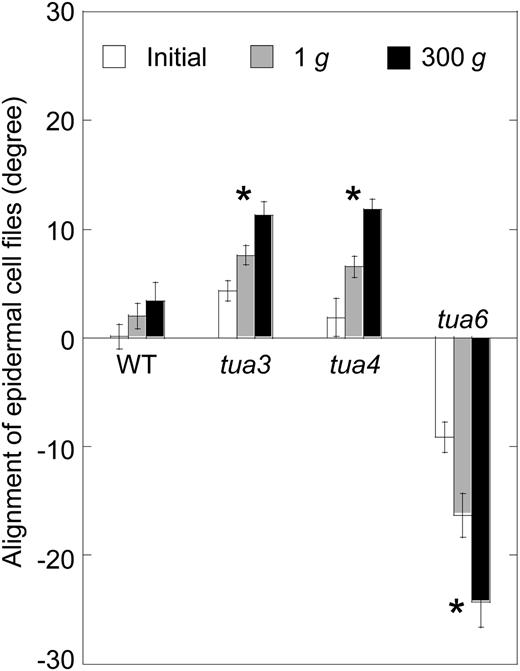
Effects of hypergravity on alignment angle of epidermal cell files in hypocotyls of Arabidopsis tubulin mutants. Wild type (WT) and tubulin mutants were grown as in Figure 1, and the angle of cells 10 to 12 to the longitudinal axis was measured using a protractor, as in Figure 3. Values are means ± se (n = 20). *, Mean values with significant differences between 1g and 300g treatments (P < 0.05).
Effects of Hypergravity on Alignment of Cortical Microtubules
In epidermal cells in the upper half of 48-h-old etiolated hypocotyls, cortical microtubules adjacent to the outer tangential wall were predominantly transverse to the long axis of the cell, whereas cells with longitudinal microtubules were predominant in the lower half. During growth for the following 24 h, reorientation of cortical microtubules from transverse to longitudinal directions occurred in just above the middle of hypocotyls (cells 10–12 from the top), with no changes in orientation in other regions. The results match with the report by Le et al. (2005).
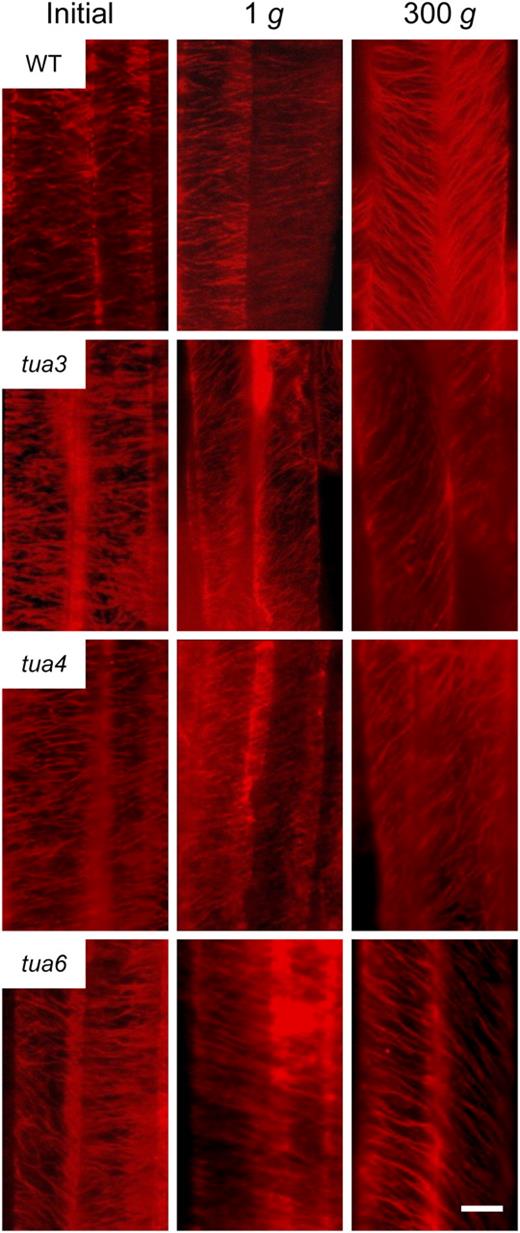
Immunofluorescence images of cortical microtubules in hypocotyls of Arabidopsis tubulin mutants. Wild type (WT) and tubulin mutants were grown as in Figure 1. Epidermal cells 10 to 12 were stained as described in “Materials and Methods.” Typical examples of two adjacent cells with distinct orientation of cortical microtubules are shown. The bar denotes 10 μm.
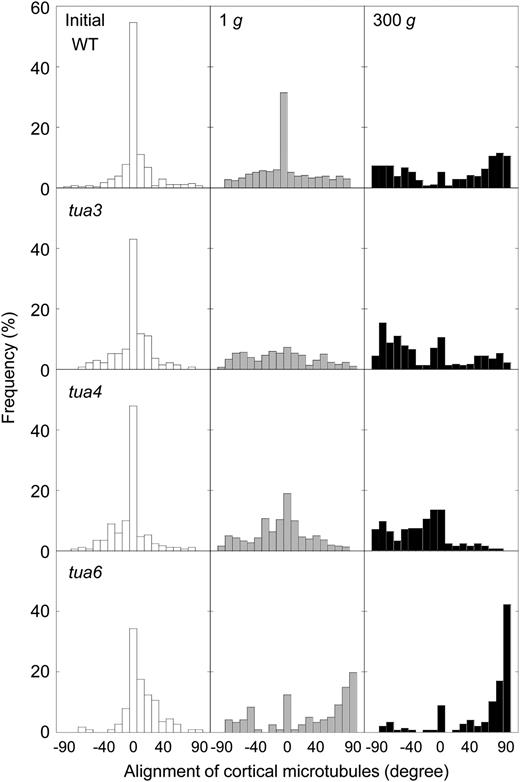
Frequency distribution of alignment of cortical microtubules in hypocotyls of Arabidopsis tubulin mutants. Wild type (WT) and tubulin mutants were grown as in Figure 1. All values were taken from immunofluorescence micrographs at cells 10 to 12, as shown in Figure 5. Data were obtained from epidermal cells in 10 different hypocotyls and normalized as percentage.

Effects of hypergravity on alignment of cortical microtubules in hypocotyls of Arabidopsis tubulin mutants. Wild type (WT) and tubulin mutants were grown as in Figure 1. Values are mean alignment angle of cortical microtubules measured in Figure 6 ± se (n = 114–334). *, Mean values with significant differences between 1g and 300g treatments (P < 0.05).
Effects of Gadolinium Ions on Alignments of Cell Files and Cortical Microtubules
We examined the effects of blockers of mechanoreceptors on phenotypes of tubulin mutants. Gadolinium ions up to 30 μm had no effect on elongation growth of hypocotyls in the wild type or tua6 mutant (Supplemental Fig. S1). In the presence of 100 μm gadolinium ions, elongation growth was suppressed and abnormal growth was induced. Gadolinium ions did not clearly influence the alignment angle of epidermal cell files in the wild type, but in tua6 they decreased the alignment angle by increasing the concentrations (Supplemental Fig. S1). Lanthanum ions had similar effects on elongation growth and the alignment of cell files (data not shown). Thus, we used gadolinium ions at 30 μm in the following experiments.
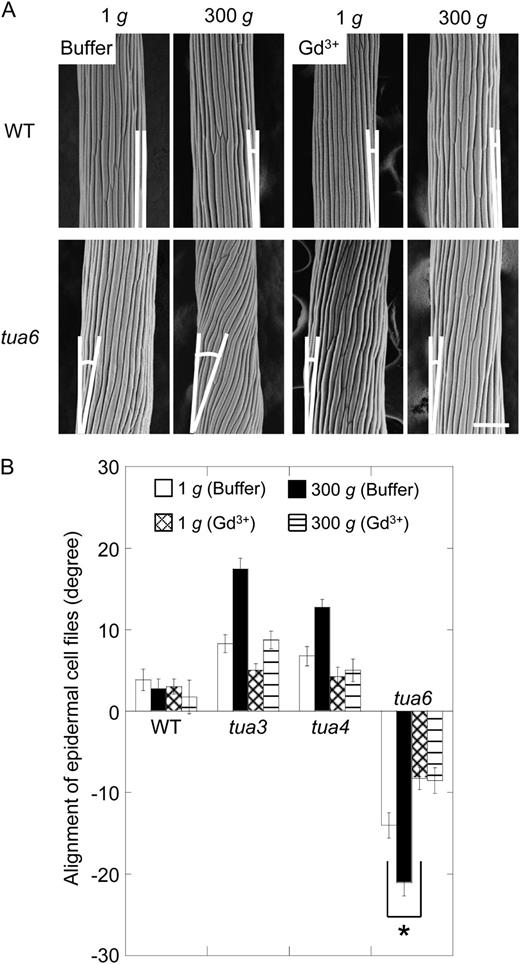
Effects of gadolinium ions on alignment of epidermal cell files in hypocotyls of Arabidopsis tubulin mutants. Arabidopsis seedlings were grown at 1g or 300g in the presence (Gd3+) or absence (buffer) of 30 μm gadolinium chloride for 72 h at 25°C. A, The outer surface of cells 10 to 12 was observed as in Figure 3. The bar denotes 100 μm. B, The angle of epidermal cell files was measured as in Figure 4. Values are means ± se (n = 20). WT, Wild type.
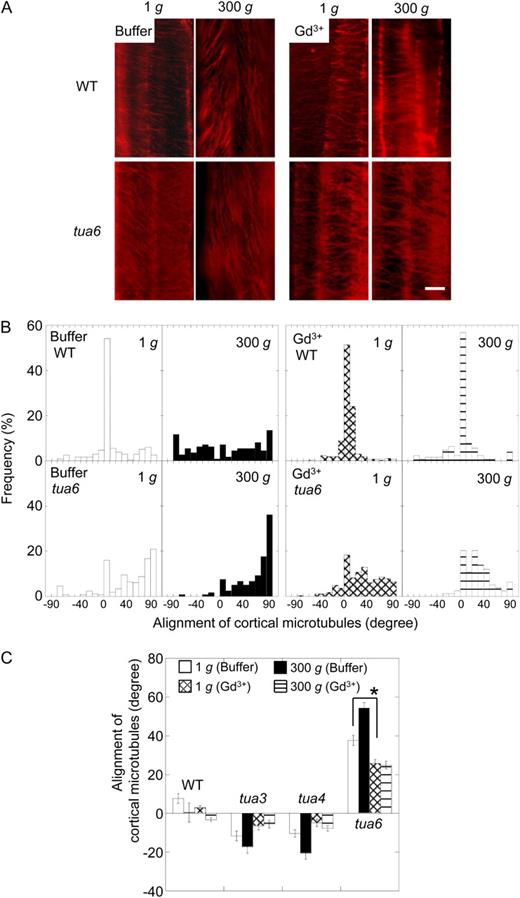
Effects of gadolinium ions on alignment of cortical microtubules in hypocotyls of Arabidopsis tubulin mutants. Wild type (WT) and tubulin mutants were grown as in Figure 8. A, Immunofluorescence images of cortical microtubules. Epidermal cells 10 to 12 were stained as described in “Materials and Methods.” B, Frequency distribution of alignment of cortical microtubules. All values were taken from immunofluorescence micrographs at cells 10 to 12. C, Mean alignment angle of cortical microtubules measured in B ± se (n = 111–278).
DISCUSSION
Hypocotyls of tubulin mutants showed either left-handed or right-handed helical growth, at 1g (Figs. 3 and 4), as observed in other organs such as flower petals, rosette leaves, and roots (Hashimoto, 2002; Thitamadee et al., 2002). Hypergravity further intensified the twisting phenotype of the mutants, in particular in tua6 (Figs. 3 and 4). Helical growth of Arabidopsis hypocotyls was also induced by propyzamide and colchicine even at 1g (Furutani et al., 2000; Matsumoto et al., 2007), and further intensified by hypergravity (Matsumoto et al., 2007). Thus, helical growth may be induced by disintegration of or modification to the structure of cortical microtubules under influence of the gravitational force. In hypocotyls, the left-handed helical growth mutants had right-handed microtubule arrays, whereas the right-handed mutant had left-handed arrays (Figs. 5–7), as observed in root epidermis of tubulin mutants (Ishida et al., 2007). The inclination of epidermal cell files was most obvious at cells 10 to 12 where reorientation of cortical microtubules from transverse to longitudinal directions occurred. The degree of inclination of microtubule arrays was further increased by hypergravity (Figs. 5–7), and there was a close correlation (r = ™0.97) between the degree of helical growth and alignment angle of cortical microtubules. These results suggest that reorientation of cortical microtubules from transverse to longitudinal directions without any bias toward the left or the right induces only simple modification to growth anisotropy, but if reorientation occurs with a bias toward either direction, helical growth may be induced.
Gadolinium and lanthanum ions have been used as blockers of mechanosensitive ion channels in various materials (Ding and Pickard, 1993; Fasano et al., 2002). Both blockers are capable of nullifying hypergravity-induced modifications to growth anisotropy and cell wall properties, as well as reorientation of cortical microtubules (Soga et al., 2004, 2005, 2006). If mechanoreceptors are involved in gravity-dependent modifications to morphology and orientation of microtubule arrays in tubulin mutants, these phenotypes are expected to be suppressed by the blockers. Actually, gadolinium ions decreased the alignment angle of epidermal cell files and that of cortical microtubules of tubulin mutants at 1g (Figs. 8 and 9). Moreover, hypergravity had no effect on the alignment angles of cell files or cortical microtubules in the presence of gadolinium ions (Figs. 8 and 9). These results suggest that mechanoreceptors are involved in modifications to morphology and orientation of microtubule arrays by 1g gravity and hypergravity in tubulin mutants.
Hypergravity suppressed elongation growth and increased lateral expansion of hypocotyls in wild-type Arabidopsis (Fig. 1), as reported in azuki bean epicotyls (Soga et al., 2006). Hypocotyls of tubulin mutants were shorter and thicker than the wild type even at 1g, and hypergravity further suppressed elongation and stimulated expansion (Fig. 1). However, the degree of such changes was clearly smaller in tubulin mutants; in particular in tua6 no further changes were induced by hypergravity, even if they still elongate significantly at 1g (Fig. 1). It is possible that suppression of elongation growth and stimulation of lateral expansion were simply caused by an inclination of cell files. However, inclination of cell files as well as reorientation of cortical microtubules was most obvious at cells 10 to 12 from the top, where active elongation growth has almost ceased (Fig. 2). In addition, the calculated decrease in longitudinal length of hypocotyls based on the pitch angle of inclined cell files (maximally 7%) is too small for the measured decrease (maximally 33%). Moreover, gadolinium ions suppressed the twisting growth phenotype and orientation of microtubule arrays toward the left or the right (Figs. 8 and 9), but they did not influence elongation growth in tubulin mutants (Supplemental Fig. S3). Thus, helical growth may not be the direct cause of modification to growth anisotropy in tubulin mutants. The mechanism of modification to growth anisotropy in tubulin mutants remains to be clarified.
In the mutants used in this study, mutant tubulin proteins are incorporated into the microtubule polymer and produce dominant-negative effects (Ishida et al., 2007). The left-handed helical growth mutant tua4(S178Δ) has one amino acid deletion in a loop preceding helix 5 at the intradimer between α- and β-tubulins (Loݶwe et al., 2001). On the other hand, the lateral association between the protofilaments is mediated largely by the M loop and its interacting regions of the adjacent protofilament (Nogales et al., 1999), and mutations at the region, such as tua6(A281T), led mostly to right-handed helical growth (Ishida et al., 2007). In addition, when GFP or hemaglutinin epitope tag was fused to the N terminus of α- or β-tubulin molecules, plants expressing the modified β-tubulins were phenotypically normal and possessed transversely oriented cortical arrays in the epidermal cells of root elongation zone, whereas the expression of modified α-tubulins caused helical growth (Abe and Hashimoto, 2005). N terminus of α-tubulin is positioned close to loop T7 at the tubulin interdimer interface, where α-tubulin acts to stimulate GTP hydrolysis in β-tubulin (Nogales et al., 1999; Nogales, 2000; Li et al., 2002). The twisting phenotype of these mutants was intensified by hypergravity (Figs. 3 and 4). Thus, the longitudinal interface between α- and β-tubulins and the lateral interface between two adjacent protofilaments may be important for normal organization of microtubules and sensitivity to the gravitational force. Out of three mutants used in this study, the twisting phenotype and inclination of cortical microtubules were most clearly observed in tua6. The expression of TUA6 was most prominently up-regulated under hypergravity conditions among six α-tubulin genes (Matsumoto et al., 2007), which may be related to strong phenotype in tua6.
Plants generally have a shorter and thicker body under hypergravity conditions (Soga et al., 2006). On the other hand, elongation growth of Arabidopsis hypocotyls and rice (Oryza sativa) coleoptiles was stimulated under microgravity conditions in space (Hoson et al., 2002; Soga et al., 2002). Growth parameters such as elongation rate of Arabidopsis hypocotyls varied in proportion to the logarithm of the magnitude of gravity in the range from microgravity to hypergravity (Soga et al., 2001; Hoson and Soga, 2003). Plant stems are rather resistant to gravity stimuli, and in this study we used 300g to induce clear effects. However, hypergravity of 300g brings about only half of the changes in growth parameters, as compared with microgravity in space (approximately 10™4g). Moreover, exposure to 300g had no effect on growth in the presence of lanthanum or gadolinium ions, which are blockers of mechanosensitive ion channels (Soga et al., 2004, 2005; Supplemental Fig. S3). These results indicate that 300g is not an excessive stimulus, because hypocotyls respond to this magnitude of force physiologically normal (Hoson and Soga, 2003). Some other organs are more sensitive to gravity stimuli; for instance, growth of pollen tubes was clearly suppressed by hypergravity at 4g (Musgrave et al., 2009). According to the nature of dose response mentioned above, hypergravity at 4g is estimated to bring about a quarter of the changes in growth parameters, as compared with 300g.
In this study, we found that hypocotyls of Arabidopsis tubulin mutants showed helical growth even at 1g, which was further intensified by 300g (Figs. 3 and 4). Such modifications to development of hypocotyls by gravity and by tubulin mutation were accompanied by reorientation of cortical microtubules from transverse to longitudinal directions with a bias toward the left or the right direction (Figs. 5–7). Furthermore, these phenotypes of tubulin mutants were clearly suppressed by blockers of mechanoreceptors not only at 300g but also at 1g (Figs. 8 and 9). These results suggest that tubulin mutants are hypersensitive to the gravitational force and the effects of gravity are saturated at lower doses. To confirm this possibility, we carried out a space experiment termed Resist Wall in the European Modular Cultivation System on the International Space Station. It was expected that under microgravity condition in space, the defects of growth in tubulin mutants were rescued and they could grow and develop more or less normally (Hoson et al., 2007). Unfortunately, we faced great difficulties in initial watering because of serious anomalies of water supply system and no plants developed to the expected developmental stage (Hoson et al., 2009). We are preparing for the next space experiment to verify the hypothesis.
MATERIALS AND METHODS
Plant Materials and Growth Conditions
Three mutants of Arabidopsis (Arabidopsis thaliana ‘Columbia’) were used in this study: tua3 has the Asp-205-to-Asn mutation in the TUA3, tua4 has the deletion of Ser-180 in the TUA4, and tua6 has the Ala-281-to-Thr mutation in the TUA6 protein. The details of these mutations were as described previously (Ishida et al., 2007). The mutation of TUA4 is a precise one-amino acid deletion in a loop preceding Helix 5 (Loݶwe et al., 2001) and situates very close to the mutated residue in lefty1 and lefty2 (Thitamadee et al., 2002). The mutation of TUA6 is at the lateral contact regions between the microtubule protofilaments mediated largely by the M loop and its interacting regions of the adjacent protofilament (Nogales et al., 1999). The mutation of TUA3 did not map to particular functional regions defined by the structural analysis. These seeds were sterilized in 2% (v/v) sodium hypochlorite solution for 1 min, and then washed thoroughly with water. The sterilized seeds were planted on 1.5% (w/v) agar medium in a 25 mL centrifuge tube, kept at 4°C for 2 d, and exposed to white light (5 W m™2 at seed level) for 6 h to induce germination. For gadolinium treatment, the sterilized seeds were planted on 1.5% (w/v) agar containing 1 mm MES-KOH buffer (pH 6.0) with various concentrations of gadolinium chloride. Seeds were then incubated in the dark at 25°C.
To analyze the growth behavior, we exposed the seedlings to hypergravity at 300g for 24 h at 25°C in the dark with a centrifuge (H-28-F; Kokusan). After the treatment, we measured the length of hypocotyls using a scale, and the thickness of hypocotyls with a digital multiangle stereoscopic microscope system (VB-G25; Keyence). SEM (TM-1000; Hitachi) was used to measure the angle of epidermal cell files to the longitudinal axis of hypocotyls. Because cells 10 to 12 from the top showed typical helical growth, we used this region for measurements of the angle of the cortical cell line. In the measurement, the longitudinal axis of hypocotyls was defined as 0°. The cortical cell lines aligned at angles between 0° and 90° (counterclockwise from the top) form left-handed helical growth, whereas those aligned at angles between 0° and ™90° form right-handed helical growth.
Immunofluorescence Microscopy
Whole seedlings were fixed with a mixture of 1.5% (v/v) formaldehyde and 0.5% (v/v) glutaraldehyde in PEMT buffer (50 mm PIPES, 2 mm EGTA, 2 mm MgSO4, 0.05% [v/v] Triton X-100, pH 7.2) for 40 min and rinsed three times in PEMT buffer. They were subsequently treated with 0.05% (w/v) Pectolyase Y-23 (Kyowa Chemical Products) and 0.05% (w/v) Cellulase Y-C (Kyowa Chemical Products) in PEMT buffer with 0.4 m mannitol for 20 min at 30°C, and then rinsed three times in PEMT buffer. Seedlings were treated with methanol at ™20°C for 10 min, and rehydrated in phosphate-buffered saline (PBS) for 10 min. Autofluorescence caused by free aldehydes from glutaraldehyde fixation was reduced with 1 mg mL™1 NaBH4 in PBS for 15 min, followed by the treatment with 1% (w/v) bovine serum albumin (BSA) in incubation buffer (50 mm glycine in PBS) for 30 min. After incubating in 0.5% Triton X-100 for 1 h, seedlings were incubated with primary antibodies against α-tubulin (product T6199, Sigma) diluted 1:1,000 (v/v) in 1% BSA at 30°C for 16 h, and rinsed three times with incubation buffer. Seedlings were then incubated with Cy3-conjugated anti-mouse IgG (product C2181, Sigma) diluted 1:100 in (v/v) in 1% BSA for 3 h at 37°C, and rinsed three times with PBS. They were mounted on a slide glass and covered with a solution containing 90% (v/v) glycerol. Immunofluorescence images were collected with a fluorescence microscope (Axioskop 2 plus; Carl Zeiss) equipped with a cooled CCD camera (VB-7000, Keyence), and processed with bundled image processing software (BZ-H1A, Keyence).
Quantitative analysis of cortical microtubule alignment was carried out, as reported by Sugimoto et al. (2003). In measurement of cortical microtubule alignment, transverse orientation was defined as 0° to the long axis of the hypocotyls. The cortical microtubules aligned at angles between 0° and 90° (clockwise from 0°) form left-handed helix, whereas those aligned at angles between 0° and ™90° (counterclockwise from 0°) form right-handed helix.
Sequence data from this article can be found in the GenBank/EMBL data libraries under accession numbers M17189 (TUA3), M84697 (TUA4), and M84699 (TUA6).
Supplemental Data
The following materials are available in the online version of this article.
Supplemental Figure S1. Effects of gadolinium ions on elongation growth and alignment of epidermal cell files in hypocotyls of Arabidopsis tubulin mutants.
Supplemental Figure S2. Effects of gadolinium ions on alignment of cortical microtubules in hypocotyls of Arabidopsis tubulin mutants.
Supplemental Figure S3. Effects of gadolinium ions on elongation growth in hypocotyls of Arabidopsis tubulin mutants.
ACKNOWLEDGMENTS
We thank Dr. Sumiko Kaihara for helpful advice in preparing the manuscript.
LITERATURE CITED
Ding JP, Pickard BG (
Furutani I, Watanabe Y, Prieto R, Masukawa M, Suzuki K, Naoi K, Thitamadee S, Shikanai T, Hashimoto T (
Hoson T, Matsumoto S, Soga K, Wakabayashi K, Hashimoto T, Sonobe S, Muranaka T, Kamisaka S, Kamada M, Omori K, et al (
Hoson T, Matsumoto S, Soga K, Wakabayashi K, Hashimoto T, Sonobe S, Muranaka T, Kamisaka S, Kamada M, Omori K, et al (
Le J, Vandenbussche F, De Cnodder T, Van der Straeten D, Verbelen JP (
Lloyd C, Chan J (
Matsumoto S, Saito Y, Kumasaki S, Soga K, Wakabayashi K, Hoson T (
Author notes
This work was supported in part by Grants-in-Aid for Scientific Research from the Ministry of Education, Culture, Sports, Science and Technology (grant no. 17510159) and a Grant for Ground-Based Research for Space Utilization from the Japan Space Forum (to T. Hoson).
Corresponding author; e-mail [email protected].
The author responsible for distribution of materials integral to the findings presented in this article in accordance with the policy described in the Instructions for Authors (www.plantphysiol.org) is: Takayuki Hoson ([email protected]).
The online version of this article contains Web-only data.
Open Access articles can be viewed online without a subscription.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}