





Abstract
Urbanization represents a multi-dimensional ecological ‘filter’ for birds determined by a myriad of variables that can change over time. Birds colonising an urban system or staying in a habitat that has been recently urbanised need to overcome both the extrinsic (e.g. food predictability, human activities, and inter-specific interaction) and intrinsic filter variables, ranging from genetic to behavioural changes and/or adjustments. An increasing body of knowledge has identified the behavioural component as crucial for individuals facing the spatiotemporal dynamic urban filters, often after other traits and mechanisms have played their role. Through both developmental (i.e. variability in the expression of genes during ontogeny) and activation plasticity (i.e. alteration of behaviour as a result of individual experience), studies have shown that the identification of cues in novel systems—often determined by extrinsic factors—and learning processes, among other factors, have important impacts on decision-making and innovation. The latter are crucial behavioural traits for thriving in urban settings. Thus, we propose an integrative mechanistic framework based on the process experienced by birds who reach a city and manage to persist in the novel system (becoming urban ‘utilisers’) or those that dwell in an urbanised region who increase their fitness through behavioural responses and adaptations, leading to population persistence (becoming ‘dwellers’). Future field research efforts ought not only to widen the range of focal species, regions, and temporal scales of studies, but also to assess behavioural responses in highly urbanised settings, given that much of our knowledge comes from studies performed in urban greenspaces. Additionally, experimental studies are needed to complement the evidence from field research and to determine causal links.
Introduction
As flying animals, most birds have access to sites where habitat connectivity—to some extent—is not a determinant factor, at least for exploration. Thus, birds can reach and explore unknown areas, with some species performing better in well-connected landscapes (MacGregor-Fors et al. 2018). In terms of a landscape perspective, urbanisation represents a multi-dimensional ecological ‘filter’, including the physical urban component (Croci, Butet, and Clergeau 2008; Evans, Newson, and Gaston 2009; Evans et al. 2011; Aronson et al. 2014; Puga-Caballero, MacGregor-Fors, and Ortega-Álvarez 2014; Sol et al. 2014). Here, we consider ‘urban’ as the areas that have been developed (with >50% built cover on a landscape-scale) to provide housing and work infrastructure to cover, at least, basic needs (e.g. electricity, water supply and drainage) for at least 1000 people/km2 (sensuMacGregor-Fors 2011). The extent to which urbanisation acts as a filter is determined by a set of traits that can represent both limitations and opportunities for those individuals and populations seeking to colonize an urban system or remain in a habitat that has been recently urbanised. Some avian traits that impede species from living or thriving in cities range from genetic characteristics (e.g. polymorphism related to personality) and/or plasticity, to morphological, physiological, and behavioural ones. The effects of these traits may compound one another with changing environmental stimuli—including infrastructure, food predictability, human activities, urban hazards (e.g. collisions with artificial structures, depredation by cats and parasitism; Santiago-Alarcon and Delgado-V 2017), and inter-specific interactions (Martin and Bonier 2018)—the fate of an individual or population when facing the initial process of exploring the urban life. Recently, Aronson et al. (2016) synthesised some of the major, hierarchical, filters that determine urban species pools (i.e. climate, biogeography, anthropogenic facilitation, nature of urbanisation, development history, socioeconomic and cultural factors, and species interactions).
General population-level responses to urbanisation
For a species to succeed in an urban system, a complex process needs to take place, in which meta-population dynamics can play a major role. Fischer et al. (2015) elegantly captured such population level perspective in the schematic categorisation of biodiversity responses to urbanisation (partially based on the urban ‘avoider’, ‘adapter’ and ‘exploiter’ scheme proposed by Blair 1996). The framework of Fischer et al. (2015) recognises the importance of population dynamics at a landscape scale and considers how non-urban populations influence their presence and numbers within cities. While ‘urban dwellers’ are proposed to be those that can persist in urban systems independent of the immigration from non-urban areas, the persistence of ‘urban utilisers’ does depend on meta-population dynamics, where the immigration from non-urban areas is fundamental. Considering this schematic categorisation of biodiversity responses to urbanisation, González-Lagos and Quesada (2017) suggested a broad framework that sets the avian decision-making process of colonising an urban system or staying in a habitat that has been recently urbanised.
Integrative conceptual framework on avian urban filtering
Based on previous reviews and conceptual studies (e.g. Croci, Butet, and Clergeau 2008; Leveau 2018), here we propose an integrative mechanistic framework based on the process by which an individual reaches a city and manages to persist in the novel urban system. We synthesize previous proposed frameworks related with the community and population responses that species have shown to follow in order to establish a broader, more systemic, view of the phenomenon. Our framework is focused on birds for two main reasons: (i) they are the best studied, and therefore understood, mobile wildlife group in urban systems (McKinney 2008) and (ii) recent evidence shows that different wildlife groups that manage to have complex assemblages in urban systems can respond differently to urbanisation, with birds representing a great bioindicator of urbanness (e.g. Sattler et al. 2014; MacGregor-Fors et al. 2016). It is important to highlight that the environmental conditions of a given urbanised landscape often include complex landscape matrices in which spatial heterogeneity tends to increase towards the human settlement (Batty 2008; Pickett et al. 2011). The components of such urban complexity, often conceived through the categorisation of land-uses (e.g. Blair 1996), have spatiotemporal dynamic ‘filters’. The coarseness of these filters is an abstraction to represent how broad or narrow the probability is for a species to ecologically enter or not a city, or urban scenarios. Thus, such coarseness will always vary in relation to its predictors, both intrinsic (e.g. mechanisms: genetics, plasticity and maternal effects; traits: behaviour, morphology and physiology) and extrinsic to individuals (e.g. infrastructure, food predictability, human activities, social and economic drivers, urban hazards and inter-specific interactions). For instance, in this abstraction, a narrow filter (related with urban ‘avoiders’) would be represented by that of a species that is behaviourally shy, not successful competing for resources with other urban birds, and with highly specific nesting conditions not present or scarce in urban areas. On the other hand, a broad filter example could be that of a highly bold, food generalist species that copes with human activities and that can successfully nest in urban buildings. This last scenario could apply to both ‘utilisers’ and ‘dwellers’, which response would depend on the particular set of combinations of the intrinsic and extrinsic factors that conform the spatiotemporal filter occurring for a given species in a given time (Fig. 1).
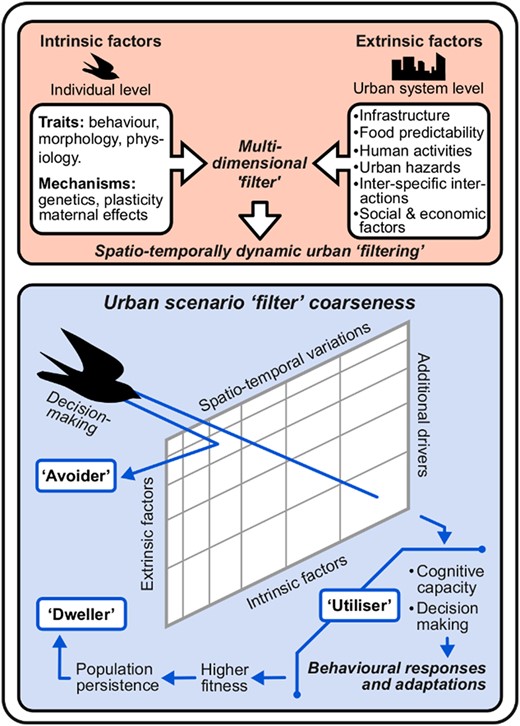
Graphical representation of an integrative conceptual framework on avian urban filtering. It includes: (i) the multi-dimensional set of ‘filters’ that individuals need to go through in highly dynamic urban scenarios, which vary spatially and temporally are given by a set of intrinsic and extrinsic filter factors (upper peach panel) and (ii) the coarseness of such ‘filters’ (abstraction on the narrowness/broadness of the probability for a species to ecologically enter or not a city) and the behavioural responses and adaptations that classify species in relation with their response to urbanisation (i.e. ‘avoider’, ‘utiliser’ and ‘dweller’) (lower blue panel). ‘Utilisers’, through behavioural responses and adaptations, could become ‘dwellers’ by increasing their fitness and leading to population persistence (independence of non-urban individuals in meta-population dynamics to maintain the urban population of a species; for a further detailed description of the framework, see the section ‘An integrative conceptual framework on avian urban filtering’).
As previously stated, one fundamental component of individuals facing dynamic urban ‘filters’ and thriving in a city is the behavioural one (González-Lagos and Quesada 2017). It is important to highlight that many individuals determine their fate in urban systems as a consequence of phenotype selection of their intrinsic traits (Partecke 2014). These intrinsic traits, which can often be adjusted under stress conditions, can facilitate urban life and have been related to the adaptedness of individuals (Alberti, Marzluff, and Hunt 2017). In fact, there is current global evidence regarding the non-random filtering of regional species pools in urban areas, which is closely related with the evolutionary and life-history traits of the assessed avian species (La Sorte et al. 2018). Regarding the latter, there are two different conceptualisations that need to be considered in this framework—including all possible intermediate scenarios—(i) pre-existent traits (e.g. phylogenetically shared traits) that often take generations to prevail and (ii) current adjustable traits (e.g. plasticity and epigenetics) that can even take a few generations to persist, often referred to as innovations (Partecke 2014).
The decision-making process of staying or leaving from a given urban scenario is largely determined by the ability of finding and successfully using vital resources, and being able to perceive and react to the urban hazards, resulting in establishing a life strategy towards thriving in the novel environment (Emlen 1974; Sol, Lapiedra, and González-Lagos 2013). When an individual is able to overcome the elements of the multi-dimensional urban filter and stays in such environment, an important factor for its survival and reproduction is its ability to detect environmental cues that carry information on the expected long-term fitness in the new area (Greggor et al. 2014; Carthey and Blumstein 2018). In light of the novel cues to use resources, conditions, and hazards (Faeth et al. 2005; Slabbekoorn 2013; Santiago-Alarcon and Delgado-V 2017), behavioural adjustments often act in two ways, with the individual having: (i) the ability to detect habitat selection cues and (ii) the plasticity to perform the required life-history changes needed to overcome a given urban scenario (Sol, Lapiedra, and González-Lagos 2013).
In responding to urbanisation, phenotypic variation is mostly mediated by pre-existing traits (van Burskirk 2012; Partecke 2014). Some species do particularly well in urban environments given a set of pre-existing behavioural, morphological and/or physiological traits that are consequences of their evolutionary history (Sol and Maspons 2016). Although the ability to modify habitat selection cues seems to be highly relevant in the initial process of colonisation or when spatiotemporal changes occur within its urban range, in the process of an individual—and further populations and species—adapting to urban conditions, the adaptive ‘flexibility’ of life-history traits gains importance. Variations of reproductive output or survival through alternative strategies (e.g. reproductive diapauses, intra-annual multi-broods) can be crucial in the population persistence of species in becoming an ‘urban utiliser’ (sensuPelletier and Garant 2012; Fischer et al. 2015). Yet, the type and spectrum of the behavioural strategies that an individual can follow is often constrained by the evolutionary history of its species, as well as its adaptiveness (i.e. phenotypic variance; Sol and Maspons 2016).
On the role of behavioural adaptation
The role of behaviour is fundamental in understanding how some organisms can reach, explore, adapt and even thrive (in the case of the well-known urban-related species; e.g. house sparrow-Passer domesticus, rock pigeon-Columba livia, and European starling-Sturnus vulgaris; Aronson et al. 2014) in urban conditions (Hermes et al. 2015). Fortunately, we currently have enough evidence to consider the behavioural component as a crucial one in the filtering process.
Life for species that manage to thrive within cities is given, at least partially, by the rapid change that behaviour represents in the light of a novel condition (González-Lagos and Quesada 2017). Such adaptiveness can be genetic-based (i.e. developmental plasticity; Snell-Rood 2013), but can also occur through activational plasticity (i.e. alteration of behaviour as a result of individual experience), that is mainly mediated by learning processes (Snell-Rood 2013; Hermes et al. 2015). During the learning process, birds acquire new information through their own or social experiences by means of trial and error, and related rewards. This has been studied in urban birds by means of alert and flight initiation distances, where habituation to human disturbance seems to be faster in urban conditions (e.g. Vincze et al. 2016).
Previous studies have highlighted the importance of having large brains in relation to body size in the colonisation of new scenarios (Lefebvre, Reader, and Sol 2004; Sol, Lapiedra, and González-Lagos 2013; but see Dale, Lifjeld, and Rowe 2015); yet, other traits, such as diet, sociability, sedentariness and preferred nesting sites, have also been linked with avian urban life (i.e. urban ‘dwellers’ and ‘utilisers’; Kark et al. 2007). It is notable that acquired information through social experience is particularly important, as it can represent a motor of innovation, which has been identified as a key skill to thrive in the city (Brown 2012). For learning processes to result in a plastic response, they require vast cognitive baggage (Sol and Maspons 2016). Cognitive capacities (i.e. behavioural adaptability and innovation in foraging, communicating, selecting mates and interacting in social groups; Balda, Kamil, and Bednekoff 1996) are essential to explain this type of plasticity, which is largely generated by innovation processes. Furthermore, the innovative mechanism represents an additional evolutionary paradigm (Baldwin effect; Lopez-Sepulcre and Kokko 2012), as it does not require genetic expression to be perpetuated in following generations, but rather occurs through cultural transmission (e.g. social and public information; Danchin et al. 2004). Some authors consider that this behaviour can be eventually fixed as a genetic expression (i.e. genetic integration; Müller 2010; Partecke 2014).
Thus, the role of intrinsic factors, both traits and mechanisms, can help the understanding of the reasons behind some birds avoiding urban conditions, others being able to withstand urban life and others thriving within cities. Such a complex process involves several organisation levels. These levels can vary from mechanisms (e.g. genetics, plasticity, maternal effects and epigenetics), which can determine the expression of traits (e.g. individuals making decisions, learning and innovating), to population dynamics, which have been suggested as an adequate level of organisation to abstract the response of birds to urbanisation [Fischer et al. 2015; see Blair (1996) for an initial classification]. Although urban ‘avoiders’ may respond by dwelling in non-urbanised sites (they can rarely occur in urbanised regions and may persist—even with small viable populations—in ‘natural’ areas embedded within urban centres; Fischer et al. 2015), the response of urban ‘utilisers’ and ‘dwellers’ is related to a much more complex set of scenarios regarding population dynamics and ecological requirement thresholds (simplified in Fig. 1 for clarity).
From our perspective and in the light of the avian urban bird ecology knowledge, there are two main ways of a bird responding as an ‘utiliser’ or a ‘dweller’. On the one hand, a given species may have intrinsic traits related to their evolutionary history that allow it to be successful for urban life. These species often have life-history strategies in which their populations are less prone to large stochastic population fluctuations that can accidentally result in local or regional extinction processes. Known as the life-history buffer (Pimm 1991), this may occur when set in a context where individuals are able to exploit resources, avoid urban risks, and interact with other species (Lewontin 1965; Sol and Maspons 2016). On the other hand, species can have the ability to change their intrinsic traits and adjust some of their life-history traits (e.g. survival reproductions and migration patterns; McDonnell and Hahs 2015). These highly plastic species are often able to trigger adaptive mechanisms, of which examples of behavioural adjustments and adaptations have populated the literature [see Gil and Brumm (2014) for recent topic-specific revisions].
Conclusions
Given the variety of avian strategies to respond to urbanisation, population dynamics determine, by definition (sensuFischer et al. 2015), if a species becomes an urban ‘utiliser’ or ‘dweller’. An urban ‘utiliser’ may be capable of going through many of the spatiotemporally dynamic set of urban-related filters, being able to respond and even adapt to a given urban condition; yet, the persistence of their urban populations still depends on meta-population dynamics with individual influx from non-urban populations. For a species to manage its way toward being an urban ‘dweller’, it needs to successfully overcome all limitations, both intrinsic and extrinsic (filters; see Fig. 1 for details). The resulting effect in fitness would allow the species to make its urban populations independent of non-urban ones, rendering them highly unlikely to be extirpated from the colonised urban system or land-use (Fischer et al. 2015). Nevertheless, it is crucial to consider that given the spatiotemporal variability of the urban filters and the heterogeneous complexity of cities, the way in which species respond to urbanisation can change over time or among landscapes or regions, as intrinsic and extrinsic filtering processes could be driven by a different set of variables depending on their traits and intrinsic mechanisms. Future field research efforts ought not only to widen the range of focal species (representative of the regional life-histories pools), regions and temporal scales of studies, but also to assess behavioural responses in highly urbanised settings (i.e. non-greenspaces), given that much of our knowledge comes from studies performed in urban greenspaces. Additionally, both field and laboratory experimental studies are needed to complement the evidence from field research and to determine causal links.
Authors’ contributions
I.M.-F., J.Q. and M.G.-A. conceived the idea. I.M.-F. and M.G.-A. led the writing of the manuscript. All authors contributed critically to the drafts and gave final approval for publication.
Acknowledgements
We are deeply thankful to Paige S. Warren, Diego Santiago-Alarcon, and Christine Rega-Brodsky, as well as two anonymous reviewers and the journal’s Editor-in-Chief (Mark J. McDonnell) for their valuable insight that undoubtedly improved the clarity of our framework.
Funding
This research did not receive any specific grant from funding agencies in the public, commercial or not-for-profit sectors.
Data Availability
No data were used in this commentary.
Declaration of interests
The authors declare that they have no competing interests.

{kind=link}