





Abstract
Host-feeding patterns of Culex pipiens L. collected in southwest suburban Chicago in 2005 were studied using polymerase chain reaction (PCR) and DNA sequencing techniques. Culex spp. mosquitoes, most identified to Cx. pipiens and the remainder to Cx. restuans by PCR, had fed on 18 avian species, most commonly American robin (Turdus migratorious), house sparrow (Passer domesticus), and mourning dove (Zenaida macroura). Additional blood meals were derived from four mammal species, primarily humans and raccoons (Procyon lotor). During a West Nile virus (WNV) epidemic in 2005, West Nile virus (WNV) RNA was detected in heads and thoraces of five Cx. pipiens (n = 335, 1.5%) using quantitative PCR. The hosts of these virus-infected, blood-fed mosquitoes included two American robins, one house sparrow, and one human. This is the first report of a WNV-infected Cx. pipiens mosquito collected during an epidemic of WNV that was found to have bitten a human. These results fulfill a criterion for incrimination of Cx. pipiens as a bridge vector.
West Nile virus (WNV) is now endemic throughout temperate North America, with annual amplification events and regular epizootics and epidemics. Although individuals of >60 mosquito species have tested positive for WNV (Centers for Disease Control, West Nile virus home page), species in the genus Culex, especially Culex pipiens L. in the eastern United States north of 36° latitude, have been implicated as the primary enzootic vectors, i.e., those responsible for transmission among bird reservoir hosts (Marra et al. 2004, Turell et al. 2005). More than 23 mosquito species have been implicated as potential bridge vectors or epidemic vectors, i.e., those responsible for transmission to humans (Marra et al. 2004, Turell et al. 2005). Recent, indirect evidence based on blood-meal analysis and theory suggests that Cx. pipiens serves as both an enzootic and an epidemic (i.e., "bridge") vector (Apperson et al. 2004, Kilpatrick et al. 2005). The evidence includes data documenting Cx. pipiens feeding on both birds and mammals (Apperson et al. 2004) and results of a analytical risk model incorporating data on virus infection and feeding rates on birds, mammals, and humans suggesting that Cx. pipiens and Cx. restuans are responsible for 80% of human WNV infection in the northeastern United States (Kilpatrick et al. 2005). However, empirical data showing a virus-infected mosquito biting a human has heretofore been lacking. Our objective was to use blood-meal analysis, individual mosquito virus infection detection, and molecular species identification methods to identify enzootic and epidemic mosquito vectors of WNV in an endemic transmission area in suburban Chicago, IL.
Materials and Methods
We sampled blood-fed mosquitoes in southwest suburban Chicago in 2005 using gravid traps and aspirators at 15 study sites consisting of 11 residential neighborhoods, 3 cemeteries, and 1 wildlife refuge. This region has had a high incidence of human cases of West Nile viral meningoencephalitis since 2002 and of St. Louis encephalitis historically in 1975 (Ruiz et al. 2004). Blood-fed mosquitoes were processed individually to identify the blood-meal source, to identify the mosquito species, and to detect WNV. To identify the blood-meal source, we first scored the Sella stage of blood-meal digestion using an ordinal rating system (Detinova 1962) and digital photography. The abdomen was removed for blood-meal analysis and polymerase chain reaction (PCR) identification of the mosquito species, whereas the thorax and head were retained for RNA extraction and quantitative reverse transcriptase (RT)-PCR for virus detection. We amplified the vertebrate mitochondrial cytochrome B gene in the blood meal using four separate PCRs with established primer pairs, purified the amplicon (if present), directly sequenced it, and compared the sequences to those in GenBank (Apperson et al. 2002, Cupp et al. 2004, Molaei et al. 2006). Results of negative controls were acceptable, and all positive controls (blood from 17 species of birds, 9 species of mammals, and 2 species of amphibians) were accurately and consistently identified to species. The same extracted DNA used for blood-meal analysis also was used for PCR-based molecular identification of Culex species to verify all morphological identifications (Crabtree et al. 1995). A quantitative RT-PCR method was used to detect WNV RNA in the head and thorax using empirically derived crossover thresholds to determine positive samples (Lanciotti et al. 2000, Hamer et al. 2007).
Results
Of 398 blood-fed mosquitoes, most (84%) were identified as Culex spp. morphologically and as Cx. pipiens by PCR. Blood meals of 246 individual Cx. pipiens (n = 335, 73.4%) were identified successfully to an avian or mammalian host (Table 1). The success of identifying a blood meal was negatively correlated with the Sella score of the abdomen (r = -0.90, df = 4, P = < 0.01). Based on the identified blood meals, Cx. pipiens fed primarily on birds (n = 191, 77.6%), with 18 species identified. The most common avian blood sources were American robin (Turdus migratorius; n = 86), house sparrow (Passer domesticus; n = 39), and mourning dove (Zenaida macroura; n = 33). Mammal feeding accounted for the remaining 22.4% of the Culex spp. blood meals, with humans (n = 51) and raccoons (Procyon lotor; n = 12) as the most common blood sources. Blood-meal analysis results for Cx. restuans revealed similar rates of avian feeding (n = 27 or 35, 77%) and similar avian and mammalian host species as Cx. pipiens.
Blood-meal analysis of mosquitoes collected in southwest suburban Chicago in 2005

Blood-meal analysis of mosquitoes collected in southwest suburban Chicago in 2005
WNV RNA was detected in 5 of 335 blood-fed Cx. pipiens mosquitoes (1.5%) using quantitative RT-PCR (Table 2); two additional blood-fed Culex spp. mosquitoes tested positive for WNV but did not yield an amplicon after the blood-meal analysis PCR. The blood-meal host was identified in four of the five WNV-positive Cx. pipiens as follows: American robin (n = 2), house sparrow (n = 1), and human (n = 1).
Blood-fed mosquitoes collected in suburban Chicago, IL, that tested positive for WNV
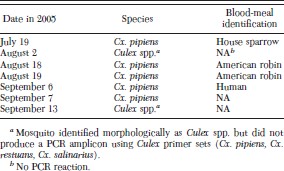
Blood-fed mosquitoes collected in suburban Chicago, IL, that tested positive for WNV
Discussion
Culex pipiens is commonly considered to be ornithophilic (Marra et al. 2004, Turell et al. 2005), although the percentage of bird feeding varies substantially by region (84–96% feeding on birds in New York, Apperson et al. 2002, 2004; 93% in Connecticut, Molaei et al. 2006; 71% in Tennessee, but only 35% in New Jersey, Apperson et al. 2004; 87% in Maryland and Washington, DC, Kilpatrick et al. 2006; and 78% in this study). One possible explanation for this variation is substantial substructuring of Cx. pipiens populations, with mammal and bird feeding forms and hybrids, identified in Europe and New York (Fonseca et al. 2004, Kent et al. 2007). The structure of the population in metropolitan Chicago is not known, but our results show that the mammal feeding rate was comparatively high.
Samples from residential areas such as backyards of homes yielded 75% of the total Culex spp. mosquitoes in our study. Other recent blood-meal analysis studies with Cx. pipiens were done within urban areas, but actual sample sites were parks, uninhabited military forts, sewage treatment plants, golf courses, wood lots, and public thoroughfares (Apperson et al. 2002, 2004, Molaei et al. 2006). Collecting blood-fed mosquitoes in immediate proximity to human habitation could explain our finding of a high frequency of human feeding by Culex mosquitoes. Host availability was not quantified in this study; however, results of blood-meal analysis should not be misconstrued as merely reflecting host preferences.
We found that 7 of 398 individual blood-fed mosquitoes were infected with WNV for an infection rate of 18 individuals per 1,000 (1.8%). Other research efforts conducted at the same sites in the same year found 227 positive pools of 1,195 tested, for an infection rate of 11 per 1,000 mosquitoes (1.1%) (Hamer et al. 2007). The Chicago area experienced a WNV epizootic and epidemic in 2005, during a drought, with Illinois ranking second in the United States in the total number of human WNV cases (Centers for Disease Control, West Nile virus home page). The high Culex spp. infection rate that reached 59 per 1,000 in late July (Hamer et al. 2007) provided an opportunity to identify virus-positive blood-fed mosquitoes and simultaneously to determine the origin of their blood meal. During peak transmission in July 2005, the probability that a WNV-infected Culex spp. mosquito fed on a human was 0.01 (i.e., WNV mosquito infection rate of 0.059 × human feeding rate of 0.173).
Incrimination of an epidemic vector of WNV requires demonstration of direct association of an infected vector with humans, a corollary of Koch’s postulates. Our study contributes to the substantial evidence that Cx. pipiens serves as the enzootic vector of WNV in large parts of North America by showing that virus-infected mosquitoes feed on American robins and house sparrows. Moreover, our study implicates Cx. pipiens as a bridge or epidemic vector of WNV, because a mosquito with a disseminated virus infection in the head plus thorax was found to have fed on a human, the first recorded observation of such an event. Although the identity, infection, and clinical status of the bitten human are unknown, this finding adds support to the hypothesis that Cx. pipiens serves as both an enzootic and bridge vector. Our experience has been that WNV infection is rather common in Culex spp. and Culex pipiens in particular, but rare in non-Culex mosquitoes. In fact, we have found that non-Culex mosquitoes are typically rare during epidemics of WNV at our study site, whether infected or not. Mosquito testing efforts in 2005–2006 in the same region resulted in only seven positive non-Culex pools of 859 tested (an infection rate of 1 per 1,000 mosquitoes), those being one positive pool each of Aedes vexans (Meigen), Anopheles quadrimaculatus (Say), and Ochlerotatus triseriatus (Say) and two positive pools each of Coquillettidia perturbans (Walker) and Ochlerotatus trivittatus (Coquillett) (Hamer et al. 2007). In comparison, of 2,016 Culex spp. pools tested, 425 were positive (estimated infection rate by maximum likelihood method of 12 per 1,000 mosquitoes, a figure comparable to the infection rate of 18 per 1,000 reported here for individually tested mosquitoes). Although we cannot completely exclude the possibility of other mosquito species playing a role as bridge vectors in the Chicago metropolitan area, the finding of a virus-positive Cx. pipiens with a human-derived blood meal combined with the relatively high rate of human feeding by Cx. pipiens suggests that control efforts focused on Cx. pipiens alone may largely reduce both epizootic amplification and transmission risk to humans.
Acknowledgements
We thank L. Abernathy, B. Bullard, M. Goshorn, J. McClain, M. Neville, B. Pultorak, and T. Thompson for providing assistance in the field and laboratory and W. Reisen, M. Turell, and C. Jones for valuable improvements to the manuscript. This research was supported by the NSF/NIH program in the Ecology of Infectious Diseases (Grant 04-29124).
References Cited
