





Abstract
The effects of different pyrethroid insecticides, formulations, and doses on peridomestic populations of Triatoma infestans (Klug) were evaluated in 128 houses with 148 identified infested peridomestic sites in northwestern Argentina between October 2003 and March 2005. Four treatments were randomly assigned within each community: two doses of 5% suspension concentrate β-cypermethrin in water applied with manual compression sprayers, the standard dose (S) at 50 mg and a double dose (2S) at 100 mg active ingredient (AI)/m2; and two emulsifiable concentrates diluted in diesel fuel and applied with power sprayers, 25% cypermethrin (100 mg [AI]/m2) (CF) and 10% permethrin (170 mg [AI]/m2) (DF). Infestation was assessed by timed manual collections with a dislodging agent at baseline, 5, 12, and 17 mo postspraying, and the sites found to be reinfested at 5 mo postspraying were selectively resprayed. Only 2S eliminated T. infestans from all peridomestic sites up to 12 mo postspraying, and it was significantly more effective than all other treatments. At 5 mo postspraying, more sites treated with CF or DF rather than S had bug colonies that probably represented residual foci, which they also failed in eliminating after a second spray. At 17 mo postspraying, the prevalence of reinfested peridomestic sites was 5% for 2S, 29% for S, 43% for CF, and 54% for DF. The application of suspension concentrate pyrethroids in dose twice as large as that currently in use in the attack phase produces a greater initial impact and may eliminate peridomestic populations of T. infestans.
Trypanosoma cruzi, the causative agent of Chagas disease, and Triatoma infestans (Kug), its main vector, are widely prevalent in the Gran Chaco, a semiarid landscape extending over Argentina, Bolivia, Paraguay, and southwestern Brazil. T. infestans has long been the target of successive control programs that eliminated it from extensive areas in Brazil, Chile, and Uruguay and led to a strong decline in human prevalence and incidence of T. cruzi (Segura et al. 2000, Dias et al. 2002). In the Argentine Chaco, however, symptomatic acute cases of Chagas disease are increasingly reported since 2001 (Ministerio de Salud y Acción Social 2003). Disorganized decentralization of vector control programs in the early 1980s followed by diminishing operational capacity since the 1990s, further compounded by the acute economic crisis in late 2001, led to the present scenario of persistent peridomestic infestation with recurrent domestic recolonization by T. infestans and renewed transmission to humans in the most affected regions (Gürtler et al. 2005).
The ongoing regional elimination program of T. infestans in the southern cone of South America is mostly based on the residual application of pyrethroid insecticides (Schofield 2001, Dias et al. 2002). Pyrethroid insecticides have proven much more effective in domiciles than in peridomestic areas (WHO 1998). In the Chaco region, the success of insecticide control campaigns in rural communities is limited by early reinfestation apparently originating from residual peridomestic foci (Guillén et al. 1997, Cecere et al. 1997). Peridomestic sites were the first to be recolonized, sustained dense populations of T. infestans, and increased the risk of domestic reinfestation in rural northern Argentina (Cecere et al. 1997, 2002). Early and persistent peridomestic infestation after spraying with standard doses of pyrethroids against Triatoma brasiliensis (Neiva) and Triatoma pseudomaculata (Stål) in Brazil (Diotaiuti et al. 2000) and Triatoma pallidipennis (Stål) in Mexico (Ramsey et al. 2003) suggests that this may be a general pattern in arid or semiarid areas. The poor effects of pyrethroid insecticides against peridomestic T. infestans and other triatomines is thought to be due to their short-lasting residual effects in outdoor sites exposed to sunlight, high temperatures, rain, and dust (Roussel 1982, Gürtler et al. 2004). The residual activity of pyrethroid insecticides also is affected by the type of formulation (WHO 1998).
In peridomestic areas, the abundance of natural hiding places for triatomine bugs that are difficult to reach with insecticides (Diotaiuti and Pinto 1991, Cecere et al. 1997) and the frequent addition of new structures or materials from infested areas may contribute to the persistence of bug populations, despite control attempts. Suspension concentrate (SC) deltamethrin applied with manual compression sprayers at 25 mg/m2 (the standard treatment) was the most effective insecticide and formulation among those tested in 350 rural houses in the southern Argentine Chaco, but it failed to eliminate T. infestans from peridomestic sites 1 yr postspraying (Gürtler et al. 2004). The effects of the tested insecticides were modified by the local abundance of bugs before spraying, as in previous trials targeting domestic T. infestans populations (Gürtler et al. 1994, Cecere et al. 2002). Preliminary observations by vector control personnel in Argentina suggested that application of pyrethroid insecticides diluted in diesel fuel with power sprayers apparently eliminated peridomestic populations of T. infestans. Another alternative that may increase the effectiveness of insecticide treatments is to augment the target insecticide dose. The effects of such measures have not been assessed in a randomized field trial. Here, we report the relative effects of different pyrethroid insecticides, formulations, and doses on peridomestic T. infestans populations in rural northwestern Argentina, and reexamine the relationship between reinfestation and the local abundance of bugs before spraying.
Materials and Methods
Study Area
The study was carried out in five rural communities in Figueroa Department (27° 23′ S, 63° 29′ W), Province of Santiago del Estero, Argentina: Vaca Huañuna (VH; number of houses n [n] = 103), Bajo Cequeira (BC; n = 37), Barrio Nueva Esperanza (BNE; n = 44), Barrio San Francisco (BSF; n = 20), and Santa Ana (SA; n = 6). These communities are located within 9–12 km of a paved road and were surrounded by other infested communities distanced >600 m. The maximum distance among the study communities was 21 km. All communities are located in a semiarid plain with hardwood forest undergoing intensive exploitation. Most houses are made of adobe walls and thatched roofs, with one or two adjacent bedrooms and a front veranda 5–10 m in width. These areas share a common roof and are referred to hereafter as domestic or domiciliary areas. The peridomestic areas separated from human habitations consist of a patio and three to eight structures (e.g., store rooms, kitchens, and corrals) that house different kind of domestic animals (Canale et al. 2000). The area was selected because it had high domestic and peridomestic infestation, and it had a recent active transmission of T. cruzi to humans. The last insecticide spray had been conducted by the National Vector Control Program (NVCP) 3 yr before the present trial.
Study Design
The study included a baseline survey of triatomine infestation followed immediately by insecticide spraying in October 2003 and assessment of triatomine infestations in March and October 2004 and March 2005.
Baseline Survey
At each house visited, we explained the objectives and the project phases; surveyed the type and number of animals, and type of building material of domestic and peridomestic structures; requested information of insecticide use and family size; georeferenced the main sites (e.g., pig and goat corrals, kitchen, store room, chicken coop, and corn store) by using a global positioning system (GeoXM, Trimble, Sunnyvale, CA); numbered each house; and posted a note of visit. Four teams, each one composed of one supervisor and three skilled collectors from the NVCP, searched for triatomine bugs in all domestic and peridomestic sites by using timed manual collections (TMC) with a dislodging agent (0.2% tetramethrin, Espacial 0.2, Reopen SA, Buenos Aires, Argentina). Two people searched for bugs in the peridomestic sites most frequently found infested (i.e., goat or sheep corral, pig corral, chicken coop, and storeroom-kitchen) by using 0.25 person-hour on each site, whereas another person searched in domiciliary sites during 30 min. All bugs collected were identified according to species and stage following Canale et al. (2000).
Intervention Phase
Immediately after the baseline survey, a community-wide insecticide spraying aiming at full coverage was carried out by NVCP staff supervised by the research team between 27 October and 3 November 2003. The insecticide spraying was extended to other nearby communities during 16–29 November 2005, but these houses were not included in the analysis. Infested and noninfested houses, as determined by TMC, were listed alphabetically and randomly assigned to a treatment within each community by means of a table of random numbers. All communities received all treatments. Four teams, each composed of one supervisor and three NVCP spray persons, treated peridomestic sites with 5% SC β-cypermethrin (Sipertrin, Chemotecnica, Spegazzini, Argentina) in water applied with 5-liter manual compression sprayers with Nell 8002 fan nozzles at 50 mg (AI)/m2 (S) and at 100 mg (AI)/m2 (2S); 25% emulsifiable concentrate (EC) cis-cyperthrin (Cipermetrina atanor 25, Atanor, at 100 mg [AI]/m2)(CF), and 10% EC permethrin (Depe, Chemotecnica, Buenos Aires, Argentina) at 170 mg (AI)/m2) (DF), both diluted in diesel fuel and applied with Fontán power sprayers fitted with 84 (75 ml/min) nozzles. Before spraying, the discharge volume per minute of every sprayer fitted with each type of plastic nozzle tip was assessed using water. The effects of the application of diesel fuel with power sprayers on six peridomestic sites infested at baseline were assessed at 5 mo postspraying in SA.
All domiciles were sprayed with SC β-cypermethrin (Sipertrin, Chemotecnica) at the standard dose of 50 mg (AI)/m2 or with 2.5% SC deltametrhin (K-Othrina, Bayer, Munro, Argentina, 25 mg [AI]/m2) applied in water with 5-liter manual compression sprayers, following standard procedures (Gualtieri et al. 1984). The fences of goat and sheep corrals, made of piled branches or shrubs, were lifted from the ground during insecticide application to increase penetration into the matrix of vegetation and goat droppings. Samples from two water reservoirs in VH and BC were analyzed to assess water quality (pH 7.6; saltiness, 863–1,553 mg/liter; hardness, 248–494 mg CaCO3/liter); these values do not affect the bioavailability of insecticides.
Surveillance Phase
The intervention effects were assessed 5, 12, and 17 mo postspraying. At 5 mo postspraying (between 14 and 20 March 2004), three teams, each including one supervisor and two bug collectors, searched for triatomines by using 0.2% tetramethrin as dislodgant spray. They searched for 15 min per person in each of 127 peridomestic sites found infested at baseline (from 118 TMC-positive sites before spraying and 30 positive sites by insecticide knock-down during spraying) and in 75 peridomestic sites previously found negative. Some sites positive at baseline could not be evaluated because householders were absent or the structure had disappeared. No domicile was evaluated except when residents reported the finding of triatomine bugs. Only peridomestic sites infested by T. infestans were resprayed selectively on detection by repeating the randomly assigned treatment at 5 mo postspraying. After focal sprays, NVCP staff installed one tetra brick box per site for passive detection of triatomine bugs (Vazquez-Prokopec et al. 2002) in all peridomestic sites found positive before spraying, in those found negative at 5 mo postspraying, and in 14 sites found negative both before and after spraying.
The 12- and 17-mo postspraying evaluations used the same methodology and capture effort as in the baseline survey, and they were carried out between 19 and 23 October 2004 and between 7 and 20 March 2005, respectively. In total, 133 peridomestic sites infested and 66 sites negative at baseline were evaluated at 12 mo postspraying; in addition to 137 domiciles, 183 sites infested and 385 sites negative at baseline were evaluated at 17 mo postspraying. The peridomestic detection boxes installed at 5 mo postspraying (March 2004) were examined for triatomines, exuviae, eggs, and bug feces at 12 and 17 mo postspraying. Specificity of fecal smears was evaluated by the phenolphthalin test (Gürtler et al. 2001a). NCVP staff sprayed all the sites found infested by T. infestans in May 2005.
Data Analysis
The assessment of intervention effects only included 148 peridomestic sites positive for T. infestans (by TMC or by insecticide knockdown collections during spraying) at the baseline survey, which were evaluated for infestation at least once after spraying. A peridomestic compound was considered reinfested when at least one T. infestans bug was captured in any peridomestic site. A site was considered colonized when any nymph of T. infestans was captured. All sites positive by TMC after spraying were considered reinfested sites, although bugs discovered after intervention may be bugs that survived treatment (i.e., not a new infestation) or that have migrated into the trial site after intervention (Schofield 2001). The cumulative percentage of reinfestation at 5, 12, and 17 mo postspraying was estimated as the number of infested sites up to a given date divided by the total number of sites evaluated up to the same date. Repeated-measures multiple logistic regression run on Stata 7.0 (StataCorp 1999) was used to assess the effects on the binary response variable (reinfested site on a given date of evaluation) of the prespraying number of T. infestans collected by TMC per site, and three categorical independent variables each with four levels: type of ecotope (excluding "others"), community, and insecticide treatment. The group variable was the individual site, and the time variable was the evaluation date. An unstructured correlation matrix and the robust variance estimator were used.
Results
At baseline, the prevalence of infestation by T. infestans ranged from 50 to 60% in domiciles (mean, 51%) and from 56 to 78% in peridomestic compounds (mean, 68%) of the four communities. Geometric mean numbers of T. infestans collected per unit of effort were three to four bugs per domicile and five to nine bugs per peridomestic compound. The cumulative percentage of reinfestation from 5 to 17 mo postspraying increased sharply from 38 to 57% for DF, from 21 to 43% for CF, from 17 to 29% for S, and only from 0 to 6% for 2S (Fig. 1). The cumulative percentage of reinfested sites at 17 mo postspraying differed significantly among treatments (χ2 = 15.1, df = 3, P = 0.002) but not among communities (χ2 = 2.24, df = 3, P > 0.5). The infestation of peridomestic sites treated with diesel fuel applied with power sprayers varied from 67% (4/6) to 83% (4/4) at 5 mo postspraying. Repeated-measures multiple logistic regression analysis showed that the likelihood of reinfestation for DF (odds ratio, OR = 21.3, 95% confidence interval [CI] = 2.3–193.9), CF (OR = 15.1, 95% CI = 1.7–136.5), and S (OR = 8.6, 95% CI = 0.9–80.2) treatments was significantly higher than for 2S (the reference class) when adjusted by community, type of ecotope, and the prespraying number of T. infestans collected per site. At 17 mo postspraying, the overall domestic infestation was 27% (from 51% at baseline), whereas in peridomestic compounds infestation was 35% (from 68% at baseline). In general, the geometric mean number of T. infestans in infested domiciles and peridomestic compounds decreased only moderately in each community after spraying.
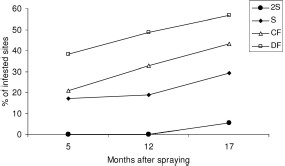
Cumulative percentage of infestation by site after insecticide spraying according to type of treatment (2S, S, CF, and DF) in four rural communities in Figueroa Department (BNE, BSF, BC, and VH), October 2003–March 2005.
The reinfestation and recolonization patterns by T. infestans followed similar trends for each treatment (Fig. 2). To focus on the appearance of newly infested sites, Fig. 2 only includes reinfested sites on initial detection. Treatment 2S achieved the lowest reinfestation (range, 0–6%) during the follow-up, followed by S (4–17%), CF (16–21%), and DF (16–38%). Insecticide effects differed significantly among treatments at 5 mo postspraying (χ2 = 7.98, df = 3, P < 0.05) but not at 12 (χ2 = 6.9, df = 3, P = 0.07) and 17 mo postspraying (χ2 = 1.85, df = 3, P = 0.60). Among 27 sites found infested at 5 mo postspraying, colonizations ranged from 40 to 78%. The earliest that an infested site treated with 2S was found recolonized was at 17 mo postspraying (Fig. 2C). The number of T. infestans per reinfested site was not significantly different among S, DF, and CF treatments at 5 mo postspraying (Kruskal–Wallis test, df = 2, P > 0.2). Mean bug abundance ranged from zero to four bugs per reinfested site.
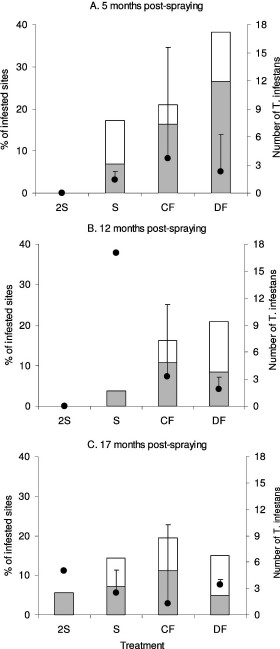
Percentage of infestation (□), colonization (

Reinfestation percentages (noncumulative) were slightly higher in goat corrals (10–37%) than in corn stores or chicken coops (16–25%), pig corrals (15–21%), and kitchens or storerooms (6–16%) for any particular survey over the follow-up (Table 1). Other ecotopes were not found infested. Only 2S eliminated T. infestans from all types of peridomestic ecotopes up to 12 mo postspraying. All other treatments (DF, CF, and S) did not eliminate T. infestans from pig or goat corrals and from kitchens or storerooms at 5 mo postspraying. At 17 mo postspraying, the percentage of reinfested pig or goat corrals (14–17%) and kitchens or storerooms (11%) treated with S was similar and lower than in other structures treated with CF and DF.
Percentage of infestation by T. infestans for sites infested and treated in October–November 2003 according to type of ecotope, date of evaluations, and type of treatment
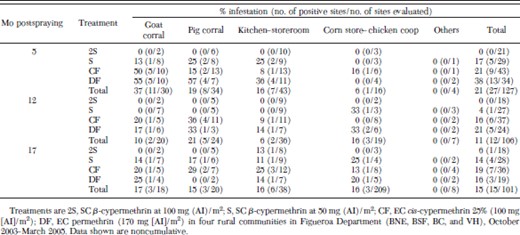
Percentage of infestation by T. infestans for sites infested and treated in October–November 2003 according to type of ecotope, date of evaluations, and type of treatment
Peridomestic detection boxes were used to check whether sites negative by TMC were actually negative. Of 70 detection boxes installed in positive sites before initial insecticide spraying and then found negative at 5 mo postspraying, 29% were lost because of faulty installation or were destroyed by animals. When the presence of any stage or exuviae of T. infestans was taken as evidence of infestation, of 49 positive sites before spraying that were TMC negative at 5 mo postspraying, at 12 mo postspryaying 41 (84%) sites were found negative by both peridomestic boxes and TMC; four were positive by both methods, and only four sites were TMC positive and box negative. When the finding of triatomine fecal smears in the detection boxes (as determined by the phenolphthalein test) also was considered as a sign of infestation, peridomestic boxes added four positive sites for triatomine bugs, which were not detected by TMC. However, these signs of reinfestation may have been from T. infestans, Triatoma guasayana (Wygodzinsky & Abalos), or Triatoma garciabesi (Stål) (other triatomine species that frequently infest peridomestic sites in the study area) because fecal smears cannot be differentiated to species level.
Reinfestation after Focal Spraying
Of 27 positive sites found reinfested after initial treatment and resprayed selectively at 5 mo postspraying, six (22%) were found positive again at 12 mo postspraying (i.e., three times positive). Moreover, of 19 sites found reinfested and resprayed at 5 mo postspraying which subsequently were found negative at 12 mo postspraying, six (32%) were found positive again at 17 mo postspraying. At 7 mo after selective respraying, in October 2004, the percentage of reinfestation for the resprayed sites ranged from 15% (2/13) for DF, 20% (1/5) for S, to 33% (3/9) for the CF treatment at 12 mo postspraying. Numbers of T. infestans per reinfested site were not significantly different among the three treatments (Kruskal-Wallis test, P > 0.5).
Bug Density-Dependent Reinfestation
The percentage of reinfestation at 5 mo postspraying increased significantly with the prespraying abundance of T. infestans per site for S (χ2 = 6.9, df = 2, P < 0.05) and DF (χ2 = 21.1, df = 2, P < 0.001), but not for CF (χ2 = 2.8, df = 2, P > 0.1) and 2S (Fig. 3). The sites treated with S that had fewer than five bugs per person-hour before spraying were less reinfested (0-8%) or recolonized (0%) at 5 mo postspraying than sites with five or more T. infestans before spraying (36% reinfested and 14% recolonized). Sites treated with DF, but not with CF, showed a similar pattern.

Percentage of infestation (□) and colonization (
Discussion
Our insecticide field trial shows that 5% SC β-cypermethrin in water applied with manual compression sprayers at twice the standard dose (2S) was the only treatment that eliminated peridomestic populations of T. infestans for 12 mo postspraying, and was significantly more effective than the standard treatment (S). The latter treatment left residual foci. Also important, the high rate of domestic reinfestation observed just 17 mo after community-wide residual spraying with insecticides attests to the great difficulties in eliminating T. infestans in the study region. Because local T. infestans populations, and all those tested so far in Santiago del Estero, have shown no increased resistance to pyrethroid insecticides, unlike elsewhere in northern Argentina (Picollo et al. 2005), the observed effectiveness of treatments was not confounded by insecticide resistance.
The finding of bugs in peridomestic sites after insecticide spraying is limited by the low sensitivity of timed manual searches aided with a dislodging agent, especially when bug densities are low (Gürtler et al. 2001b). However, successive searches for bugs at the same site during the follow-up and the use of peridomestic detection boxes may compensate for the low sensitivity of point bug searches and provide a valid outcome measure. Low-cost tetra brick boxes had a high concordance with TMC and mostly confirmed the absence of T. infestans at the treated sites. Although well accepted by householders and by vector control staff, one of the disadvantages of the detection boxes is that their performance depends on how well they are installed.
To assess the effectiveness of insecticide treatments over time, a distinction has to be made between residual foci, new colonizations, and recent invasions. Colonized sites were detected in all postspraying evaluations of all treatments, whereas the presence of only one to three adult T. infestans bugs per site occurred less frequently. Shortly after community-wide insecticide spraying, the finding of only a few adult bugs most likely represented a recent invasion by flight dispersal and is not proof of treatment failure, whereas the finding of bug colonies probably represented residual foci derived from unhatched eggs at the time of treatment or from bugs that survived exposure to the insecticides at each site. Shortly after insecticide spraying, residual foci are indicated by the finding of several late instars or adults, by some high-density bug populations, and by the positive relationship between reinfestation and local bug abundance before spraying (Gürtler et al. 2004). The higher the abundance of bugs during the egg-laying season, the greater the likelihood that some of the late instars or eggs inside crevices or hollow logs would emerge after the residual insecticide has lost effectiveness. Appropriate molecular markers may provide conclusive evidence regarding the precise sources of reinfestants (Marcet et al. 2006).
Peridomestic residual foci typically occurred in wooden structures with much of their surface exposed to extreme weather conditions, and in those that were difficult to spray adequately because of the presence of animals, stored grain, or other food (Cecere et al. 1997). The finding of recolonized sites at 12 mo postspraying (in early spring) may represent either residual foci that went undetected in the previous summer when there were very few bugs or sites that were recently invaded (from other residual foci or from an external source) and in which a bug colony became established. If passive transport can be ruled out through appropriate questions to householders, a dense bug colony with late instars at just 5 mo postspraying is most likely evidence of treatment failure.
The observed domestic and peridomestic reinfestation in all the study communities was much higher than in others in the same region treated by professional spray persons aiming at full coverage with 2.5% SC deltamethrin (Paulone et al. 1988, Cecere et al. 1997) or SC β-cypermethrin (Zerba et al. 1997) at standard doses. In the current trial, the use of several treatments differing widely in effectiveness within a given community may have caused some of the observed reinfestation and the lower than expected reduction of bug abundance in domestic and peridomestic sites. More colonized sites were recorded for CF or DF than for S at 5 mo postspraying, whereas colonized sites occurred in all treatments at 17 mo postspraying, although at different frequencies. The only site treated with 2S and found recolonized for the first time at 17 mo postspraying was at 430 m from the nearest suspected source of T. infestans. This source was the most abundant colony (114 bugs per 0.25 person-hour) detected at the baseline survey, and remained a residual focus (with 35 bugs per 0.25 person-hour) at 5 mo postspraying when it was resprayed with CF. This focus may have been the likely source of the T. infestans found 12 mo later because of the time lag between bug dispersal by flight, founding of a new colony, and detection by TMC once the colony reached a minimum abundance. The distance of 430 m is within the known flight range of T. infestans and within the observed distance between reinfested sites and the putative sources of T. infestans (Cecere et al. 2004, Vazquez-Prokopec et al. 2004). The time delay in detecting T. infestans in the neighborhood of the suspected source was within the range observed (up to 2 yr) in other rural communities under surveillance, where focal spatial statistics were applied to determine significant distances of bug abundance clustering from the source (Cecere et al. 2004, unpublished data). The contribution of other external sources to reinfestation was less likely because the closest infested communities were 3 km apart from this site. The single new bug colony detected in a site 17 mo after it was treated with 2S does not detract from the obvious advantage of the double dose treatment over the other treatments, for all of which early reinfestation foci were recorded.
The significant association between reinfestation and prespraying bug abundance per site for S and DF treatments is consistent with previous findings in bedroom areas and in peridomestic sites in northern Argentina (Gürtler et al. 1994, 2004; Cecere et al. 2002). The effects of 2S were independent of the prespraying abundance of bugs per site and probably resulted from the higher insecticide dose provoking a stronger knockdown effect, or prolonging the residual activity of β-cypermethrin, or both. Pyrethroids showed a wide range of residual activity (1–12 mo) against T. infestans indoors or under laboratory conditions (Gualtieri et al. 1984, Diotaiuti and Pinto 1991, Ferro et al. 1995, Guillén et al. 1997) but lasted very little in peridomestic sites at standard doses (Gürtler et al. 2004). The prespraying abundance of T. infestans is a key determinant of control success and therefore of the reinfestation dynamics after a single application of pyrethroid insecticides.
Both CF and DF treatments applied with power sprayers had very poor effects on peridomestic populations of T. infestans. Power sprayers have limitations for maneuvering in small, closed spaces and were not favored by the spraying teams. The microdrops of insecticide released by power sprayers probably have little adherence to the substrate and easily drift away, thus reducing the amount of insecticide deposit on the treated surface. Insecticide deposits under the intended target dose are expected to have reduced activity and leave residual foci.
Finding the insecticide dose that causes a high initial mortality with a considerable persistence under field conditions is crucial to improve the effectiveness of vector control actions. Laboratory results are good indicators for field trials but they will never substitute for them (Oliveira Filho 1988). Field studies that evaluate the effectiveness of field insecticide doses also need to consider the effects of prespraying infestation levels, environmental factors, and types of substrate treated and to assess whether the recommended doses are effective under local conditions. With an increased emphasis on community-based control programs by using insecticides with low environmental impact, we face a great challenge when selecting insecticide formulations, dosages, and frequency of application to retard the appearance of resistance in vector populations, while minimizing negative environmental impacts and achieving the desired level of effectiveness against insect vectors.
The evidence provided here supports the use of suspension concentrate pyrethroids in higher doses to eliminate T. infestans populations from peridomestic structures in high-risk rural settings. The standard dose is not appropriate for the type of infested peridomestic ecotopes found in the Chaco region. Full treatment coverage of well defined rural communities with appropriate insecticide doses is expected to prolong bug suppression beyond present levels of achievement. A cost-effectiveness analysis of the candidate treatments needs to be conducted before a final recommendation is issued. Also important, the current trial provides additional evidence showing that current tactics and procedures fail to eliminate peridomestic populations of T. infestans in rural areas and need to be revised.
Acknowledgements
We thank the staff of the NVCP at Tucumán and Leonardo Lanati for providing active support during fieldwork; residents of the communities for participation; Padre Sergio, Sara and Tuki families of Bandera Bajada for field accommodation; Silvana Ferreyra for technical assistance to set up the database; and the European Community-Latin American Network for Research on the Biology and Control of Triatominae (ECLAT) and Chagas Disease Intervention Activities-European Community (CDIA) networks for helpful discussion. María Inés Picollo (Centro de Investigaciones de Plagas e Insecticidas [CIPEIN]-Consejo Nacional de Investigaciones Científicas y Técnicas [CONICET]) kindly provided results of insecticide resistance monitoring program of T. infestans for Santiago del Estero. This study was supported by awards from the National Institutes of Health/National Science Foundation Ecology of Infectious Disease program award R01TW05836 funded by the Fogarty International Center and the National Institute of Environmental Health Sciences to U.K. and R.E.G., the Agencia Nacional de Promoción Científica y Técnica (Argentina), and the University of Buenos Aires to R.E.G. R.E.G. and M.C.C. are members of the CONICET Researcher's Career.
References Cited

{kind=link}
{kind=link}
{kind=link}