





Abstract
Visceral leishmaniasis is an endemic protozoal disease of humans and dogs in tropical and subtropical regions in Asia, Africa, southern Europe, Central America, and South America, where sand flies (genera Phlebotomus and Lutzomyia) act as vectors. An outbreak in a New York foxhound kennel and subsequent surveillance revealed widespread Leishmania infantum infection of dogs in the United States, outside the known range of the vector sand flies. For this study, we conducted surveillance for sand flies during the summers of 2001 and 2002 at two areas: on the grounds of the New York kennel and at the Institute of Ecosystem Studies (IES) 10 km away. CO2-baited light traps were used for surveillance. Populations of Lutzomyia vexator, not previously known in New York, were widespread and locally abundant (range, 0.26–1.16 flies/trap night) at the IES site. These populations showed a bimodal, midsummer activity peak and were most abundant on steep slopes within mature mixed hardwood forests. Further research will be necessary to determine whether the New York populations of L. vexator in the vicinity of the kennel could be involved in transmission of canine leishmaniasis.
In 1999, foxhounds at a hunt club in Dutchess County, southeastern New York, experienced an outbreak of canine leishmaniasis. Beginning in late summer, a number of the dogs developed a variety of signs, including bleeding, wasting, seizures, hair loss, skin lesions, renal failure, edematous limbs, and joint effusion (Gaskin et al. 2002). Cytological fluid analysis of joint aspirates followed by isolation and culture revealed infection with a protozoal parasite, Leishmania infantum, the etiological agent of Mediterranean kala-azar (Gaskin et al. 2002). Forty-one percent of 112 dogs at the kennel were later found seropositive for Leishmania (Gaskin et al. 2002), an observation that led to a nationwide investigation of 10,351 hunt club dogs; of these dogs, 1.8% were seropositive (Schantz et al. 2001). How the hunt club dogs became exposed to this parasite is not known (Enserink 2000, Gaskin et al. 2002).
The primary route of transmission of Leishmania in vertebrates, including humans, is through the bite of hematophagous sand flies of the genera Phlebotomus (Old World) or Lutzomyia (New World). Dog-to-dog transmission is also possible, particularly among animals held at close quarters (Gaskin et al. 2002), but it requires an initial infection event. Although the presence of L. infantum has not been shown in any sand flies in North America, sand fly populations exist in several parts of the United States, including the southeast, where some of the New York hunt club dogs had traveled. Initial exposure to sand fly bites may have occurred during travel to these areas.
The possibility of local exposure to bites of infected sand flies was investigated in a limited study after the outbreak, but no sand flies were found (Enserink 2000). Additionally, there had not been any previous reports of sand flies in New York. The nearest known populations of Lutzomyia shannoni, which feeds on mammals, occur in southern New Jersey. Lutzomyia vexator, which is thought to feed on reptiles, has been identified as far north as Connecticut and Ontario (Young and Perkins 1984). This study focused on conducting surveillance to detect the presence, phenology, and microhabitat associations of sand flies in areas near the New York hunt club where the canine leishmaniasis outbreak occurred. By establishing whether sand fly populations exist in the area, our intention was to assess the potential for vectors to be involved in transmission of Leishmania parasites. Even if infection of the New York dogs was entirely from dog to dog, a local population of competent vector sand flies could use the infected dogs as a reservoir for transmission of leishmaniasis to other vertebrates, including humans (Reithinger and Davies 1999).
Materials and Methods
Study Sites and General Methods.
We conducted surveillance for sand flies in the summers of 2001 and 2002 at sites in the town of Washington, Dutchess County, New York. Our two focal surveillance sites were on the grounds of the hunt club (HC) and ~10 km away at the Institute of Ecosystem Studies (IES). Both areas are characterized by hilly terrain with mature temperate deciduous forest, rocky outcroppings, mammal burrows, and decaying deadfall. The HC site is dominated by black cherry (Prunus serotina), black locust (Robinia pseudoacacia), and red maple (Acer rubrum), and the forest occurred as small (<5 ha) fragments interspersed with lawns, buildings, and roads. The 1,000-ha IES site consists of mature oak-mixed hardwood forest occurring in large (>100 ha), unfragmented areas (see Jones et al. 1998 for more details on habitat). In 2001, both HC and IES were studied, whereas in 2002, research was limited to the IES site.
We used CDC miniature light traps (John Hock and Co., Gainesville, FL) modified to include a CO2 bait (0.5 kg dry ice). We also installed fine mesh lining to permit collection of small insects and a plastic rain shield for use during inclement weather. Traps were set between 1600 and 1900 hours and collected the next morning between 0600 and 0900 hours. Traps were tied to trees and suspended ~0.75 m above the ground; each trap was associated with a habitat feature thought to be attractive to sand flies, including tree holes, mammal burrows, and rocky outcroppings. In 2001, eight traps were rotated among the HC site and two 2.25-ha plots at IES; they were set almost every day from 9 July to 9 August so that each site was trapped for 6–7 nights (48–56 trap nights). The 2.25-ha study plots at IES coincided with locations of long-term small-mammal trapping (see Ostfeld et al. 2001 for details). The study plots were separated by 150–1,500 m and covered much of the 1,000-ha site area. In 2002, trapping was conducted during 23 nights (six traps per night) from late June to late August. Traps were placed within 2 m of HOBO data loggers (Onset Computer, Bourne, MA) programmed to record temperature and humidity at six different 2.25-ha plots (six traps and 36–42 trap nights per plot) within the IES site. During the day after collection, the entire catch was systematically searched for phlebotomines, and the number and sex of any sand flies caught in each trap were recorded. Sand flies were identified according to the method by Young and Perkins (1984).
Abiotic Conditions.
In 2002, we sought to determine whether a set of abiotic habitat variables might affect abundance or activity of sand flies at the IES site. Every 2 wk, we retrieved temperature and humidity data from the dataloggers associated with each trap and generated daily (across plots) and plot-specific (through time) averages from >500,000 data points. Because Lutzomyia sand flies are entirely terrestrial and sensitive to standing water and saturated soils (Kettle 1999), we tested whether soil infiltration rate and slope (in degrees) influenced sand fly catch. To assess soil infiltration, we poured 1 liter of water into a 20-cm-diameter section of PVC pipe placed vertically on bare soil and recorded the time required for complete disappearance (runoff or percolation). Infiltration tests were performed three times at the location of each trap and were averaged to generate a plot-specific value. In addition, we used a clinometer to measure the steepest slope within 3 m of each trap, irrespective of compass direction.
Statistical Analyses.
To assess the effects of temperature and humidity on sand fly abundance, we conducted forward, stepwise multiple regression analyses with α = 0.15 for inclusion. Initially, a multiple regression was used to ask if weather variables affected the number of sand flies caught on a particular night across all sites. Independent variables were average daytime temperature (T), average nighttime T, average daytime relative humidity (RH), average nighttime RH, coefficient of variation (CV) of daytime T, CV of nighttime T, CV of daytime RH, and CV of nighttime RH. A second multiple regression was run with the same set of independent variables to assess the total number of sand flies trapped per plot across the entire sampling period.
To assess any effects of water infiltration rate, we conducted a linear regression with the mean infiltration rate of each of the plots as the independent variable and the number of flies per plot as the dependent variable. Similarly, for slope, we used the maximum percent slope at each plot as the independent variable and the number of flies as the dependent variable.
Results
In 2001 and 2002, respectively, we examined ~100,000 and 350,000 insects and identified 130 and 36 sand flies, all of which were L. vexator (Fig. 1). The catch consisted of 1.16 and 0.26 sand flies/trap night in 2001 and 2002, respectively. Females comprised 75 and 60% of the catch in 2001 and 2002, respectively. In 2001, nine of the females were engorged and four were gravid; no engorged or gravid females were captured in 2002.

Photomicrograph of female (left) and male (right) L. vexator collected at the IES in Millbrook, Dutchess County, New York.
No sand flies were captured at the HC site, but sand flies were widely distributed across the 1,000-ha IES site. In 2001, sand flies were caught on both plots and at 13 of the 16 trap sites within IES plots. In 2002, sand flies were detected at each of the six plots, but in fewer than one-half of the trap sites within plots. Within the IES site, we observed substantial variability among plots in sand fly abundance (minimum = 2, maximum = 13).
Sand fly activity was highly seasonal in both years. In 2001, the first sand fly was captured on 12 July and the last on 6 August; corresponding dates for first and last appearance in 2002 were 23 July and 12 August (Fig. 2). Some evidence of a bimodal activity peak appeared in both years, with an activity lull apparent during late July (Fig. 2).
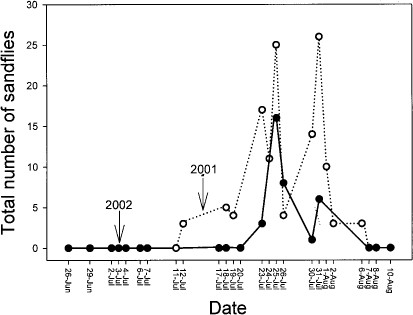
Phenology of sand fly activity at the IES in Millbrook, Dutchess County, New York.
Despite considerable variation in the six plots and through time in T and RH, neither the mean nor the CV of these weather variables had a significant effect on numbers of sand flies captured. Multiple regressions testing effects of weather on captures among plots and through time revealed no significant relationships. Water infiltration rate was not a significant predictor of numbers of flies caught at a plot (R2 = 0.423, df = 5, P = 0.124). However, the number of captures of sand flies was strongly and positively associated with percent slope (R2 = 0.872, df = 5, P = 0.006).
Discussion
This study is the first to reveal the presence of Lutzomyia sand flies in New York. Trapping over two summers in a 1,000-ha mature, oak-mixed hardwood forest within post-agricultural landscapes typical of southern New York and New England suggests that L. vexator is widespread and abundant in this region. More extensive studies, however, will be necessary to assess this species' regional distribution and abundance. Similar to other species of Lutzomyia in temperate regions (Brinson et al. 1992, Comer et al. 1994), L. vexator at our sites showed strong seasonal activity with a bimodal, midsummer peak. Activity was not strongly associated with either spatial or temporal variation in ambient temperature or relative humidity.
Lutzomyia vexator is reported to feed on reptiles (Klein et al. 1987, Schall 2000), but characterization of host distribution and feeding preferences is poor. Terrestrial reptiles active at our sites included box turtles (Terrapene carolina) and several species of snakes, including the common garter snake (Thamnophis sirtalis); lizards were rare or absent. L. vexator is capable of harboring sporozoites of Plasmodium mexicanum, the etiological agent of saurian malaria (Klein et al. 1987, Schall 2000) but has not been reported to maintain or act as a vector of Leishmania species. Further studies of the blood-meal sources and vector competence of this sand fly species for Leishmania are warranted.
Sand fly abundance was not strongly influenced by local variation in temperature and humidity, either at the spatial scale of several kilometers or over the course of a summer. However, our observation of higher capture success in areas with higher slopes and greater topography suggests that slope or some feature associated with high percent slope (e.g., greater drainage, shallower soils, thinner litter layers) will be a good predictor of sand fly abundance.
The strong midsummer activity peak we documented in both years corresponds to the late summer onset of leishmaniasis symptoms in the hunt club hounds, suggesting that sand flies should not be ruled out as vectors involved in initial disease transmission. Even if sand flies were not involved in transmission of parasites to the hunt club dogs, their presence suggests the possibility that endemic transmission could occur in the future. Arboviruses (African horse sickness and Bluetongue virus) transported in domestic livestock from endemic areas in North Africa to virgin areas in Europe have been transmitted among hosts by native Culicoides midges, which had not been infected until recently (Mellor et al. 2000). Similarly, the potential would seem to exist for dogs exposed to Leishmania during travel to southerly areas to serve as a source of infection to endemic sand fly populations to the north (Travi et al. 2002).
In conclusion, the source of the 1999 outbreak of canine leishmaniasis remains unknown, but our surveillance suggests that involvement by populations of L. vexator in the vicinity of the hunt club should not be ruled out. Moreover, these hematophagous insects could potentially spread imported cases of leishmaniasis among dogs or other vertebrate hosts. Host choice and vector competence of this sand fly species remain to be determined.
Acknowledgements
We thank E. Kiviat and S. Zahn for help with equipment and useful discussions. Portions of this research were funded by a Research Experiences for Undergraduates (REU) grant from the National Science Foundation (DBI-9988029) to the Institute of Ecosystem Studies in Millbrook, NY (A.R. Berkowitz, PI), a research grant from the National Science Foundation (DEB-0075277) to R. S. O. and C. D. Canham, and a grant from the National Institutes of Health (R01 AI40076) to R. S. O. and F. K. Two anonymous reviewers provided helpful comments. This is a contribution to the program of the Institute of Ecosystem Studies.
References Cited

{kind=link}
{kind=link}