





Abstract
In some mammals that breed cooperatively, nonbreeding female helpers nurse offspring born to the breeding female in their group, a process known as allonursing. Previous laboratory studies have suggested that primiparous and multiparous females (those that have successfully given birth once or more, respectively) are more likely to contribute to allonursing than nulliparous females (those that have not given birth). In contrast, few field studies have been able to demonstrate whether breeding experience, or parity, affects the likelihood or duration of allonursing. In this study, we investigated whether the likelihood and duration of allonursing by subordinate female meerkats (Suricata suricatta) were associated with variations in their parity (number of pregnancies reaching a viable gestational age) and their immediate pregnancy status. Comparisons showed that nonpregnant nulliparous females were less likely to allonurse than nonpregnant primiparous and multiparous females, although parity was not related to variation in allonursing duration. Parity may enhance the expression of maternal behavior when exposed to foster pups, especially since mammary gland development has been shown to be directly associated with parity in other mammals.
Allonursing, the occurrence of non-offspring nursing, has been documented in approximately 70 species (Packer et al. 1992) and is thought to confer to the offspring direct benefits relating to immunological function, growth, and survival, while being energetically costly to allonurses (Clutton-Brock et al. 1989; Roulin and Heeb 1999; König 2006). Previous studies have suggested the main hypotheses for the likelihood of allonursing to be misdirected parental care, kin selection, or milk evacuation (Hayes 2000; Roulin 2002). A few publications have overviewed the similarities of allonursing across species. For example, allonursing in monotocous species (those producing a single offspring per birth), but not polytocous (multiple offspring at birth), usually is associated with milk theft. Moreover, allonursing is most commonly observed in species living in small groups with larger litter sizes (Packer et al. 1992). In cooperatively breeding mammals, where young produced by a dominant breeding female are fed and protected by nonbreeders of either sex (Bateman et al. 2012), subordinate females that have not been pregnant also allonurse the offspring born to the dominant female, a process known as spontaneous allonursing (Creel et al. 1991; Doolan and MacDonald 1999). In most cases, only a proportion of nonbreeding females contribute to allonursing, with older nonbreeding females being more commonly involved than younger ones (Macleod et al. 2013).
Parity (the number of pregnancies reaching a viable gestational age) is known to impact both female physiology and behavior. It has been shown to affect the development of mammary glands and to enable mothers to yield more milk (Miller et al. 2006). In goats, cows, rhesus monkeys, and grey seals, primiparous females (those having given birth once) nurse longer than multiparas (having given birth more than once—Wilson 1992; Miller et al. 2006; Orman et al. 2011; Lang et al. 2012). The reason for this difference is that multiparas produce a greater volume of milk owing to a lower secretory cell density and larger alveolar size throughout lactation compared to primiparas (Lang et al. 2012). As a result, primiparas must nurse longer to provide the same amount of milk to their young as multiparas. Primiparas and multiparas are involved more commonly in allonursing than females yet to give birth at all (nulliparas) (Moltz et al. 1971; Scanlan et al. 2006). Parity also has been shown to affect the expression of allomaternal behavior. For example, in female laboratory rats, nulliparas initially show aggressiveness toward foster pups, but express maternal behaviors (in this case, nest building, nursing, licking the young, and keeping them warm) following estrogen and prolactin treatment, associated with a withdrawal of progesterone (Moltz et al. 1971).
Although it is known that allonursing is relatively common in species that form small groups, reproduce communally, and give birth in a confined space (Packer et al. 1992), we still have a limited understanding of the conditions that make females capable of allonursing in wild animals. In particular, it is not clear whether all females are able to allonurse and how experimental factors affect the incidence of allonursing. Most of the literature explores hypotheses either about the physiological mechanisms or the social and environmental influences triggering allonursing, but there remains an information gap on its ontogenetic development (Macleod et al. 2013; Montgomery et al. 2018).
Here we use long-term records of allonursing in wild Kalahari meerkats, Suricata suricatta (Carnivora: Herpestidae), to compare the incidence of allonursing in nonbreeding females with contrasting experience of reproduction. Meerkats are social mongooses living in groups of two to 50 individuals (Doolan and MacDonald 1996). A single dominant female in each group virtually monopolizes reproduction, breeding up to four times a year, delivering one to six pups per litter and lactating for about 35 days post-parturition (Russell et al. 2003). Groups commonly include several subordinate females that normally are born to the same group and either are daughters or other close relatives of the dominant female (Clutton-Brock et al. 2010). Subordinate females sometimes breed, but risk being evicted from their group by the dominant female or having their pups killed at birth if they do so (Clutton-Brock et al. 1998), resulting in their fecundity and breeding success being substantially lower than those of dominants (Clutton-Brock et al. 2001a). After the dominant female gives birth, subordinate females protect, babysit, and also may nurse the pups (Clutton-Brock et al. 2002, 2003). Our aim is to explore the effects of nulliparity versus parity on the likelihood and the duration of allonursing in subordinate female meerkats. Previous research showed that the likelihood of helpers to nurse offspring of the dominant female is related to their immediate pregnancy status, age, and whether they have recently been evicted from their group. Recently pregnant female helpers are more likely to allonurse than pregnant females, which in turn are more likely to allonurse than nonpregnant females; older and recently evicted females also are more likely to allonurse (Macleod et al. 2013). Furthermore, heavier subordinate females are more involved in babysitting and pup-feeding than lighter females (Clutton-Brock et al. 2003). The number of females in a group and litter size also were identified as factors that may influence the likelihood of allolactation, because these factors were shown to increase the rate of pup feeding in meerkats (Clutton-Brock et al. 2001b). The duration of allonursing is positively related to the body condition of all lactating females (allonurses and the mother) and subordinate females also tend to allonurse longer if the breeding female is in poor body condition (Macleod et al. 2013).
Here, we investigate the effect of parity on the likelihood and duration of allonursing in subordinate female meerkats, taking into account other factors that were shown to impact allonursing (Macleod et al. 2013). As primiparous and multiparous females appear to express maternal behaviors more quickly than nulliparous females in laboratory rats (Moltz et al. 1971), we predicted that nulliparous female meerkats would be less likely to allonurse than primiparous and multiparous females and that the duration of allonursing would be longer in primiparas and multiparas than nulliparous females, as seen in some other mammal species (Dwyer and Lawrence 1997; Lang et al. 2012).
Materials and Methods
Study system.
—This study was based on long-term data from a population of wild slender-tailed meerkats at the Kuruman River Reserve and neighboring farms, near Van Zylsrus, South Africa (26°58′S, 21°49′E). The data were collected by trained observers between December 1996 and July 2019 over an area of 50–60 km2 (Cozzi et al. 2018). Details on the study site are provided elsewhere (Russell et al. 2002).
Each meerkat was individually recognizable by dye marks on its fur and a subcutaneous transponder microchip (Hodge et al. 2008). In each group, one individual was fitted with a VHF radiocollar, enabling observers to keep track of the group by means of telemetry (Jordan et al. 2007). Dominance could be clearly identified in females, because the dominant female is the main breeder within a group and expresses a repertoire of dominance behaviors (dominance assertions) that is different from those of the subordinate females (submissive behaviors—O’Riain et al. 2000; Thavarajah et al. 2014).
Each day, between one and 20 groups were visited, each group composed of 3–47 meerkats. All individuals were habituated to the presence of observers, so it was possible to follow most groups and remain within 2 m of most individuals with minimum disturbance to their natural behaviors. Groups were followed three to five times a week. On each visit, group composition was recorded, and pregnancy and allonursing status were checked. Animals were weighed at least once a day, using small crumbs of hard-boiled egg and water to entice them onto electronic scales (Clutton-Brock et al. 2004). Pregnancies were detectable approximately 30 days after conception and births usually could be dated within 3 days due to substantial weight loss of the mothers. Conception date was back-calculated to 70 days before birth (Barrette et al. 2012). Allonursing occurred mainly overnight in the sleeping burrow (Macleod and Clutton-Brock 2015) and could be determined by the observation of dried milk circles around the nipples, usually covered by wet sand.
Data collection.
—Only litters born to dominant females were included in this study, and never while another litter was being nursed within the group, to assure that allonurses were nursing foster pups. Pregnant subordinate females included in the study either aborted, or their litter was eaten or lost within 2 weeks after birth. All litters included survived until three months old—the age at which pups cease to rely on milk and are able to feed independently (Doolan and MacDonald 1997; Brotherton et al. 2001). The birth date of the litter is referred to as “parturition day.”
The number of potential allonurses present in the group (aged at least 6 months old, based on the youngest allonurse in the population) and the number of emerging pups were recorded. We also recorded litter order for each group within each breeding year, starting in August (end of winter), until the following July: rank was attributed in ascending order each time a litter was born to a dominant or subordinate female (first litter born was coded as “1,” second as “2,” etc.).
Only subordinate females with known birthdate, pregnancy, and lactation dates were included in the study, and those nursing their own litters were not included. Females were considered as allonurses if they allonursed at least 1 day during the 3 months following parturition day. The parity of each potential allonurse was identified as to whether she had never given birth (nulliparous), whether she had given birth once (primiparous), or she had given birth multiple times (multiparous). The pregnancy status of each potential allonurse also was determined as to whether she was pregnant on parturition day, or she had aborted or given birth within a month before parturition day (referred to as “recently pregnant”). We also determined whether subordinate females had been evicted from the group during the dominant’s gestation and returned within a week after parturition day.
Statistical analysis.
—Body mass was calculated as the mean of all morning weights recorded during the week following conception day and parturition day, for litter mothers and subordinate females, respectively. The residuals of a generalized additive growth model were used to replace body mass, to account for collinearity between age and body mass (Supplementary Data SD1). The residuals provide the relative body condition of a female for her age, according to predictions of the model, which were based on the age and morning weights of dominant and subordinate females in the population, for litter mothers and subordinate females, respectively. Individuals who were nonpregnant and older than 180 days old were included in this model. Morning weights and ages were collected for all these individuals until they died or emigrated from the study. The same script was applied for the population of subordinate females and for dominant females separately. In the subordinate growth model, females were included until they acquired dominance, whereas in the dominant growth model, females only were included once they had acquired dominance, because females gain weight when they attain dominance (Russell et al. 2004). Data points falling out of 2* minimum and maximum standard deviation values were considered outliers and taken out of the model. The packages mgcv (Wood 2011) and tidyverse (Wickham et al. 2019) were used for the script.
A generalized linear mixed model, fitted with the glmmTMB package on R 3.6.1 (Brooks et al. 2017), was used to investigate the likelihood of subordinate females to allonurse, with a binomial error distribution and logit link. The model was fitted by Wald z tests. Model comparison was performed using the Akaike Information Criterion (AIC), where the lowest AIC score indicates the best fit for the model (Akaike 1974). Of all possible models, none were within 2 ΔAIC of the best model (Supplementary Data SD2). Parity of the subordinate females, their pregnancy status, their eviction status, as well as their age and body condition were considered for this model. Litter order, litter size, the number of subordinate females, the age, and body condition of the dominant mother were also taken into account.
The duration of allonursing was analyzed with a linear mixed model (LMM) using the “lme4” package (Bates et al. 2015). The model was fitted by restricted maximal likelihood (REML) and Wald t tests. Parity of the subordinate females, their pregnancy status, age, and body condition were taken into account. The number of days that groups were visited was included as an offset variable. Duration of allonursing was expressed as the total number of days subordinate females showed signs of allonursing divided by the total number of days the group was visited over the 3 months following parturition day. It accounted for the variation in the number of visits per group. Subordinate females that did not allonurse were not taken into account for this model.
For both models, collinearity for all fixed effects was tested using variance inflation factors with a threshold value of 2. To account for pseudoreplication, subordinate female identity was nested with dominant mother identity (dominant females can keep their dominance status within a group for years, during which they give birth to several litters) and included as nested random variables. Pairwise comparisons were made with Bonferroni tests, using the lsmeans package (Russell 2016). All means were reported with standard errors of the mean, unless specified otherwise.
Ethical note.
—Work was approved by University of Pretoria’s Animal Ethics Committee (#EC010‐13) and Northern Cape Nature Conservation (FAUNA 1020/2016). Research on live animals followed American Society of Mammalogists’ guidelines (Sikes 2016).
Results
The likelihood and duration of allonursing were measured in 403 litters, of which 233 (57.82%) were allonursed, and included records of allonursing by 410 subordinate females out of 1780 potential candidates, between 6 and 72 months of age (23.03%; Fig. 1). Allonurses on average spent 30.10 ± 0.75 days allonursing, out of the total lactation period. Each group was visited 75.14 ± 0.49 days on average. Overall, allonursing duration amounted to a mean of 0.40 ± 0.01 (minimum 0.014 and maximum 0.96) of the total lactation period in a litter.

—Count of allonurses (black) among the 1780 wild subordinate female meerkats included in the study (grey), depending on pregnancy status (NP—not pregnant, P—currently pregnant, and RP—recently pregnant) and parity (nulliparous, primiparous, and multiparous). Data collected from December 1996 to July 2019 at the Kuruman River Reserve, South Africa. Values above the bars represent the percentage of allonurses in each subcategory.
Factors affecting the likelihood that subordinate females will allonurse.
—The likelihood of allonursing by subordinate females varied with their parity, pregnancy status, age, and body condition (Table 1). There was a significant interaction between parity and pregnancy status (Table 1). Nonpregnant nulliparous females were less likely to allonurse than all the other females (Fig. 2). There was no significant difference between pregnant and recently pregnant nulliparas, nonpregnant, pregnant, and recently pregnant primiparas and multiparas (Fig. 2). Finally, older females and females in better body condition were more likely to allonurse (Table 1; Figs. 3A and 3B).
Factors affecting the likelihood of allonursing in 1780 wild subordinate female meerkats
| Best model | Estimate | SE | z value | P value | |
|---|---|---|---|---|---|
| Intercept | −3.34 | 0.28 | −11.97 | <0.001 | |
| Parity | Nulliparous | 0.00 | <0.001 | ||
| Primiparous | 2.47 | 0.34 | 7.25 | ||
| Multiparous | 2.41 | 0.50 | 4.86 | ||
| Pregnancy status | Not pregnant | 0.00 | <0.001 | ||
| Pregnant | 3.64 | 0.35 | 10.44 | ||
| Recently pregnant | 3.43 | 0.76 | 4.52 | ||
| Litter order | 0.00 | <0.01 | |||
| Subordinate age | 0.49 | 0.12 | 4.13 | <0.001 | |
| Litter size | 0.27 | 0.10 | 2.86 | <0.01 | |
| Number of subordinate females | −0.79 | 0.13 | −6.26 | <0.001 | |
| Subordinate body condition | 0.25 | 0.11 | 2.36 | <0.05 | |
| Parity * pregnancy status | 0.00 | <0.001 |
| Best model | Estimate | SE | z value | P value | |
|---|---|---|---|---|---|
| Intercept | −3.34 | 0.28 | −11.97 | <0.001 | |
| Parity | Nulliparous | 0.00 | <0.001 | ||
| Primiparous | 2.47 | 0.34 | 7.25 | ||
| Multiparous | 2.41 | 0.50 | 4.86 | ||
| Pregnancy status | Not pregnant | 0.00 | <0.001 | ||
| Pregnant | 3.64 | 0.35 | 10.44 | ||
| Recently pregnant | 3.43 | 0.76 | 4.52 | ||
| Litter order | 0.00 | <0.01 | |||
| Subordinate age | 0.49 | 0.12 | 4.13 | <0.001 | |
| Litter size | 0.27 | 0.10 | 2.86 | <0.01 | |
| Number of subordinate females | −0.79 | 0.13 | −6.26 | <0.001 | |
| Subordinate body condition | 0.25 | 0.11 | 2.36 | <0.05 | |
| Parity * pregnancy status | 0.00 | <0.001 |
Data collected from December 1996 to July 2019 at the Kuruman River Reserve, South Africa. Results from the best fitted model. Mother identity (n = 88) and subordinate female identity (n = 679) were included as nested random terms (n = 771).
Factors affecting the likelihood of allonursing in 1780 wild subordinate female meerkats
| Best model | Estimate | SE | z value | P value | |
|---|---|---|---|---|---|
| Intercept | −3.34 | 0.28 | −11.97 | <0.001 | |
| Parity | Nulliparous | 0.00 | <0.001 | ||
| Primiparous | 2.47 | 0.34 | 7.25 | ||
| Multiparous | 2.41 | 0.50 | 4.86 | ||
| Pregnancy status | Not pregnant | 0.00 | <0.001 | ||
| Pregnant | 3.64 | 0.35 | 10.44 | ||
| Recently pregnant | 3.43 | 0.76 | 4.52 | ||
| Litter order | 0.00 | <0.01 | |||
| Subordinate age | 0.49 | 0.12 | 4.13 | <0.001 | |
| Litter size | 0.27 | 0.10 | 2.86 | <0.01 | |
| Number of subordinate females | −0.79 | 0.13 | −6.26 | <0.001 | |
| Subordinate body condition | 0.25 | 0.11 | 2.36 | <0.05 | |
| Parity * pregnancy status | 0.00 | <0.001 |
| Best model | Estimate | SE | z value | P value | |
|---|---|---|---|---|---|
| Intercept | −3.34 | 0.28 | −11.97 | <0.001 | |
| Parity | Nulliparous | 0.00 | <0.001 | ||
| Primiparous | 2.47 | 0.34 | 7.25 | ||
| Multiparous | 2.41 | 0.50 | 4.86 | ||
| Pregnancy status | Not pregnant | 0.00 | <0.001 | ||
| Pregnant | 3.64 | 0.35 | 10.44 | ||
| Recently pregnant | 3.43 | 0.76 | 4.52 | ||
| Litter order | 0.00 | <0.01 | |||
| Subordinate age | 0.49 | 0.12 | 4.13 | <0.001 | |
| Litter size | 0.27 | 0.10 | 2.86 | <0.01 | |
| Number of subordinate females | −0.79 | 0.13 | −6.26 | <0.001 | |
| Subordinate body condition | 0.25 | 0.11 | 2.36 | <0.05 | |
| Parity * pregnancy status | 0.00 | <0.001 |
Data collected from December 1996 to July 2019 at the Kuruman River Reserve, South Africa. Results from the best fitted model. Mother identity (n = 88) and subordinate female identity (n = 679) were included as nested random terms (n = 771).
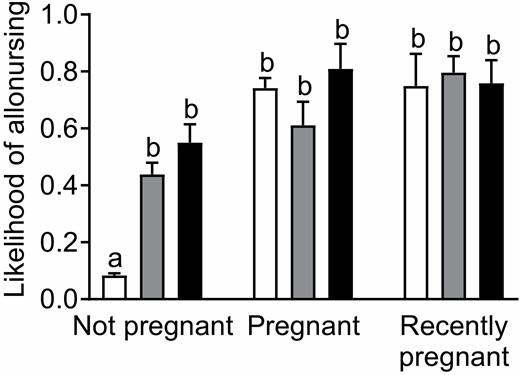
—Likelihood of allonursing in 1780 subordinate female meerkats, depending on parity (nulliparous in white, primiparous in grey, or multiparous in black) and pregnancy status (not pregnant, pregnant, or recently pregnant) on parturition day. Data collected from December 1996 to July 2019 at the Kuruman River Reserve, South Africa. Pairwise comparisons made with Bonferroni corrections. Letters depict significant differences within terms of the interaction. Means presented with standard errors.
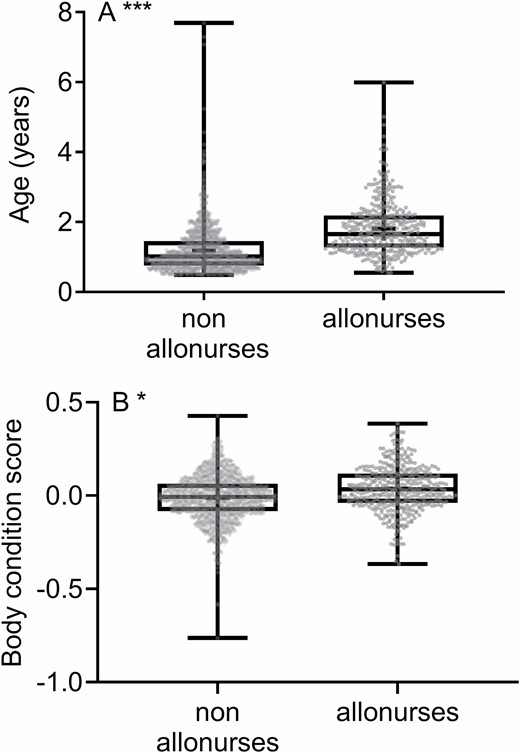
—Mean A) age and B) body condition score of the 1370 non-allonurses and 410 allonurses in meerkats. The data were collected between December 1996 and July 2019 at the Kuruman River Reserve, South Africa. “+” indicates the mean. The box represents first quartile, median, and third quartile. The whiskers represent the minimum and maximum samples. All sample points are represented by grey points. *P < 0.05; ***P < 0.001.
Likelihood of allonursing in subordinate females was related to litter order, litter size, and the number of females present in the group (Table 1). Subordinate females were more likely to allonurse larger litters (Fig. 4B) and litters born later in the breeding season (from August to the following July; Fig. 4A). Also, females were less likely to allonurse in groups where there were more females present (Table 1; Fig. 4C). Eviction status, age, and body condition of the litter’s mother did not explain any variation in the likelihood of allonursing in subordinate females and did not appear in the best model.
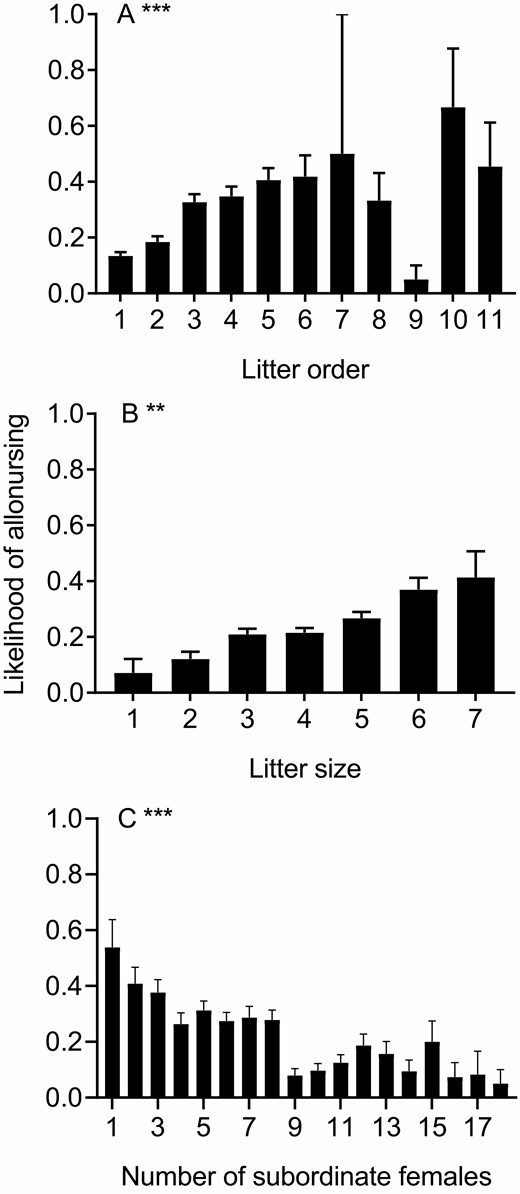
—Factors affecting the likelihood of allonursing in 1780 subordinate female meerkats: A) litter order, B) litter size, and C) number of subordinate females present on parturition day. Data collected from December 1996 to July 2019 at the Kuruman River Reserve, South Africa. Means presented with standard errors. **P < 0.01; ***P < 0.001.
Factors affecting allonursing duration in subordinate females.
—The duration of allonursing varied with pregnancy status (Table 2; Fig. 5B). Pregnant (P < 0.01) and recently pregnant females (P < 0.001) allonursed significantly longer than non-pregnant females, while there was no difference between pregnant and recently pregnant females (P = 0.74). However, parity had no effect on the duration of allonursing (Table 2; Fig. 5A). The age and body condition of allonurses were significant predictors of the duration of allonursing (Table 2; Fig. 6). Older females allonursed significantly longer than younger females (Table 2; Fig. 6A), and females in better body condition allonursed longer than those in poorer body condition (Table 2; Fig. 6B).
Factors affecting the duration of allonursing in 410 wild subordinate female meerkat allonurses
| Best model | Estimate | SE | t value | p value | |
|---|---|---|---|---|---|
| Intercept | -48.87 | 1.62 | -30.10 | <0.001 | |
| Parity | Nulliparous | 0.00 | 0.35 | ||
| Primiparous | 2.40 | 1.96 | 1.22 | ||
| Multiparous | -0.56 | 2.88 | -0.20 | ||
| Pregnancy status | Not pregnant | 0.00 | <0.001 | ||
| Pregnant | 6.73 | 1.87 | 3.60 | ||
| Recently pregnant | 8.66 | 2.25 | 3.85 | ||
| Allonurse age | 1.94 | 0.98 | 1.97 | <0.05 | |
| Allonurse body condition | 4.47 | 0.91 | 4.93 | <0.001 |
| Best model | Estimate | SE | t value | p value | |
|---|---|---|---|---|---|
| Intercept | -48.87 | 1.62 | -30.10 | <0.001 | |
| Parity | Nulliparous | 0.00 | 0.35 | ||
| Primiparous | 2.40 | 1.96 | 1.22 | ||
| Multiparous | -0.56 | 2.88 | -0.20 | ||
| Pregnancy status | Not pregnant | 0.00 | <0.001 | ||
| Pregnant | 6.73 | 1.87 | 3.60 | ||
| Recently pregnant | 8.66 | 2.25 | 3.85 | ||
| Allonurse age | 1.94 | 0.98 | 1.97 | <0.05 | |
| Allonurse body condition | 4.47 | 0.91 | 4.93 | <0.001 |
The data were collected between December 1996 and July 2019 at the Kuruman River Reserve, South Africa. Mother identity (n = 76) and subordinate female identity (n = 260) were included as nested random terms (n = 270).
Factors affecting the duration of allonursing in 410 wild subordinate female meerkat allonurses
| Best model | Estimate | SE | t value | p value | |
|---|---|---|---|---|---|
| Intercept | -48.87 | 1.62 | -30.10 | <0.001 | |
| Parity | Nulliparous | 0.00 | 0.35 | ||
| Primiparous | 2.40 | 1.96 | 1.22 | ||
| Multiparous | -0.56 | 2.88 | -0.20 | ||
| Pregnancy status | Not pregnant | 0.00 | <0.001 | ||
| Pregnant | 6.73 | 1.87 | 3.60 | ||
| Recently pregnant | 8.66 | 2.25 | 3.85 | ||
| Allonurse age | 1.94 | 0.98 | 1.97 | <0.05 | |
| Allonurse body condition | 4.47 | 0.91 | 4.93 | <0.001 |
| Best model | Estimate | SE | t value | p value | |
|---|---|---|---|---|---|
| Intercept | -48.87 | 1.62 | -30.10 | <0.001 | |
| Parity | Nulliparous | 0.00 | 0.35 | ||
| Primiparous | 2.40 | 1.96 | 1.22 | ||
| Multiparous | -0.56 | 2.88 | -0.20 | ||
| Pregnancy status | Not pregnant | 0.00 | <0.001 | ||
| Pregnant | 6.73 | 1.87 | 3.60 | ||
| Recently pregnant | 8.66 | 2.25 | 3.85 | ||
| Allonurse age | 1.94 | 0.98 | 1.97 | <0.05 | |
| Allonurse body condition | 4.47 | 0.91 | 4.93 | <0.001 |
The data were collected between December 1996 and July 2019 at the Kuruman River Reserve, South Africa. Mother identity (n = 76) and subordinate female identity (n = 260) were included as nested random terms (n = 270).

—Duration of allonursing among 410 meerkat allonurses only depending on A) parity (N—nulliparous, P—primiparous, and M—multiparous) and B) pregnancy status (NP—not pregnant, P—currently pregnant, and RP—recently pregnant) on parturition day. Data collected from December 1996 to July 2019 at the Kuruman River Reserve, South Africa. Pairwise comparisons made with Bonferroni tests. Means presented with standard errors. **P < 0.01; ***P < 0.001.

—Factors affecting the duration of allonursing among 410 meerkat allonurses: A) age of allonurses on parturition day and B) body condition of allonurses on parturition day. Data collected from December 1996 to July 2019 at the Kuruman River Reserve, South Africa. *P < 0.05; ***P < 0.001.
Discussion
Our results show how the likelihood of allonursing in subordinate meerkats is associated with individual characteristics (parity, pregnancy status, age, and body condition) and societal factors (litter order, litter size, and number of females available in the group), while the duration of allonursing seems to be mainly driven by the physiological state of the subordinate females (pregnancy status, age, and body condition). Likelihood of allonursing varied both with pregnancy status and parity, while pregnancy status, but not parity, explained the duration of allonursing. Parous females and pregnant or recently pregnant females were more likely to allonurse than nulliparous and nonpregnant females. Nonpregnant females allonursed for less time than pregnant and recently pregnant females. Older females and females in better body condition also were more likely to allonurse and allonursed longer than young females and females in poor body condition. In addition, our results show that external factors were involved in the likelihood of allonursing in meerkats. Subordinate females were more likely to allonurse for litters born later in the breeding season and they also were more likely to allonurse larger litters. However, the likelihood of allonursing decreased with increasing number of available females in the group.
These results resemble previous findings in nonpregnant ewes, where maternal behavior (licking, low-pitched vocalizations, acceptance at udder, and reduced aggression) is expressed more often and udder volume increases significantly more in multiparas than nulliparas following estradiol/progesterone treatment and vaginal and cervical stimulation (Dwyer and Lawrence 1997). In the same study, nulliparous ewes showed a stronger negative response to the presence of a lamb. In rats, primiparous and multiparous females also express maternal behaviors faster than nulliparous females when exposed to foster pups (Moltz et al. 1971). Having given birth before, primiparas and multiparas are physiologically and behaviorally primed for suckling litters. However, it is possible that some of the nulliparous females involved in this study were not yet sexually mature since the youngest females included in this study were between 6 and 12 months old and females may not be sexually mature until they are 1 year old (O’Riain et al. 2000). The significant interaction between parity and pregnancy status indicated that only nonpregnant nulliparas were less likely to allonurse, while pregnant and recently pregnant nulliparas were equally likely to allonurse as primiparas and multiparas. In previous findings (Macleod et al. 2013), pregnancy status was also involved in predicting the likelihood of allonursing in meerkats, and it was suggested that increasing prolactin levels in pregnant and recently pregnant mammals were involved, owing to its role in promoting maternal care and lactation (Falconer 1980).
The factors involved in affecting the duration of allonursing in meerkats appear to differ from those involved in triggering allonursing. Contrary to our initial expectation, there was no difference in the duration of allonursing between nulliparous, primiparous, and multiparous females, although pregnancy status did explain some of the variation. This could be explained by the fact that the development of mammary glands is controlled mainly by fluctuating hormone levels related to pregnancy and recent parturition (Svennersten-Sjaunja and Olsson 2005). Maintenance of lactation is supported by prolactin and cortisol (Buhimschi 2004), and our results suggest that its control in meerkats may be regulated by immediate physiological factors (hormones related to pregnancy, age, and body condition) and not by long-term breeding experience, i.e., parity. In addition, it has been shown that suckling is one of the main factors responsible for maintaining prolactin releases leading to lactation in many mammal species (McNeilly et al. 1983; Fuchs et al. 1984; Grosvenor et al. 1986; Lupoli et al. 2001).
The use of the long-term dataset showed that some of nonpregnant nulliparas showed signs of allonursing, although this was not common. In dogs, allonursing in nulliparous females can be a consequence of pseudopregnancy (maternal behavior and signs of lactation), which usually takes place at the end of a luteal phase (when progesterone levels drop) and is associated with high levels of prolactin (Tsutsui et al. 2007). Pseudopregnancy is one of the mechanisms that may be involved in allonursing for several species (Creel et al. 1991; Montgomery et al. 2018). Spontaneous lactation also is known to occur in rats, primates, goats, heifers, and women with no preceding pregnancy or pseudopregnancy (Erb 1977; Creel et al. 1991; Peña and Rosenfeld 2001), although the mechanisms are not fully understood.
As in previous analyses of data from the same study of meerkats (Macleod et al. 2013), subordinate females were more likely to allonurse litters born later in the breeding season. As the breeding season pass, they are more likely to have had more interaction with pups. As a result, they may become more responsive to the presence of pups, even without being physiologically primed by pregnancy or recent birth. Both parous and nulliparous female rats were shown to display long-term retention of maternal memory, which triggers maternal behavior when exposed to foster pups (Bridges 1977; Bridges and Scanlan 2005). In addition, our work showed that individuals also were more likely to allonurse larger litters and less likely to allonurse when there were more subordinate females available. Madden et al. (2009) showed that helpers respond to increased begging by increasing food provisioning. If allonursing is also triggered by pup begging, females already physiologically primed for suckling litters may be responsive to increased pup begging, which is likely to increase with litter size and to decline with the number of allonurses.
Our results also show that older females and females that are in better body condition were more likely to allonurse and did so for longer than younger females and females in poor body condition; these data should be interpreted carefully, because we only were able to study the number of days where there were signs of allonursing and not the volume produced or the duration of each suckling bout. A similar tendency for allonursing to increase in older and more experienced females has been found in other mammals. For example, in ewes, body condition has a positive effect on the daily milk yield (Doney et al. 1981). In contrast, in humans, the volume of milk produced is regulated almost entirely by infant demand, whereas body condition impacts the composition of milk (Neville 2001). In meerkats, older individuals generally are more successful foragers than younger ones and are heavier and usually in better condition than younger animals and are likely to suffer disproportionately high metabolic costs of lactation (Hanwell and Peaker 1977).
The likelihood of allonursing in subordinate meerkats is associated with individual characteristics (parity, pregnancy status, age, and body condition) and societal factors (litter order, litter size, and number of females available in the group), while the duration of allonursing seems to be mainly driven by the physiological state of the subordinate females (pregnancy status, age, and body condition).
Supplementary Material
Supplementary material are available at Journal of Mammalogy online.
Supplementary Data SD1.—R script is used to obtain residuals from the generalized additive growth model (GAM). The residuals of this model were used to replace body mass, to account for collinearity between age and body mass. The residuals provide the relative body condition of a female for her age, according to predictions of the model, which were based on the age and morning weights of nonpregnant dominant and subordinate females in the population, for litter mothers and subordinate females, respectively. Using a GAM allows the age effect on body mass to be slightly non-linear, and logging the values makes the variance more even across the range of ages.
Supplementary Data SD2.—Models considered for the study on likelihood of allonursing based on their AIC. The interaction “Parity*Pregnancy status” is included in every model. The relevance of terms were compared with Wald statistics (χ 2).
Acknowledgments
We are grateful to the Kalahari Research Trust for their permission to use the data. We thank Dave Gaynor for his input on the long-term data collection; Tim Vink for organization of the field site and database; and all the volunteers, managers, students, and researchers involved in the Kalahari Meerkat Project for access to facilities and habituated animals and the organization of data collection. Many thanks to Jack Thorley and to Zoe Allin for helping out with the growth model and for proof-reading this paper, respectively. The long-term field site Kalahari Meerkat Project was financed by the Universities of Cambridge and Zurich and the MAVA foundation. The Kalahari Meerkat Project is logistically supported by the Mammal Research Institute, University of Pretoria. We thank the Northern Cape Department of Environment and Nature Conservation for permission to carry out the research.
Funding
We also thank the French National Research Institute for Agriculture, Food, and Environment and the French National Centre for Scientific Research for their financial support to M.A.A.B. and M.K. We also thank G. Brachet, E. Chave, W. Faith, I. Ledoux, P. Brachet, the Schaeverbeke family, and S. Hirbec for their financial contribution. This work used long-term data provided by the Kalahari Meerkat Project, which was supported by the European Research Council (Research Grant Nos 294494 and 742808 [to T.H.C-.B.] since 1/7/2012) and the Human Frontier Science Program (funding reference RGP0051/2017), the University of Zurich and MAVA foundation, and with logistic support by the Mammal Research Institute, University of Pretoria.
Authors’ Contributions
M.B.M. and T.H.C.B. organized the long-term data collection with the help of several field assistants, including M.A.A.B. M.A.A.B. and P.V. analyzed the data. The article was written by M.A.A.B., A.G., and M.K. All authors gave final approval for publication. M.K. and T.H.C-.B. are co-last authors.
Conflict of Interest
The authors declare that they have no conflict of interest.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}