





Abstract
The commercial production of hops (Humulus lupulus L.) in the northeastern United States is on the rise due to demand from local breweries. Several arthropod pests are economically damaging to hop yield and quality. Due to climate and landscape differences between traditional and resurging hop-growing regions, there is a need for region-specific integrated pest management (IPM). We first review hop pest and natural enemy biology and management strategies. Then the phenology, abundance, and peak date of arthropod pests scouted in seven Vermont hop yards is reported. Documentation of natural enemy abundance is also reported. Our 3-yr survey indicated that hop aphid (Phorodon humuli (Schrank)) populations were highest in the continually cool, wet season. Potato leafhopper (Empoasca fabae (Harris)) was a pest with an unpredictable arrival date and of special concern for first-year hop plants. Twospotted spider mite (Tetranychus urticae Koch) was a pest of concern in hot, dry conditions and after some broad-spectrum pesticide applications aimed at leafhoppers. This survey was the first step toward developing appropriate IPM tactics for modern day northeastern hop production. Further research should be focused on adjusting arthropod pest thresholds, disease management, and developing alternative control options for both arthropod and disease management.
Commercial hop (Humulus lupulus L.) production has not occurred in the northeast region of the United States for 150 yr. Vermont production peaked in 1860 when the state produced 289,690 kg of dried hops (Kennedy 1860). A combination of the spread of hop downy mildew (Pseudoperonospora humuli) (Miyabe and Takah G. W. Wilson), the expansion of production in western states, and the passing of prohibition laws later in the 1920s contributed to the decline of the 19th-century northeast hop industry. Today, the Pacific Northwestern states of Washington, Oregon, and Idaho remain the dominant hop production states. However, hop production in nontraditional regions is growing and in 2013 accounted for 2% of the total U.S. hop acreage (Hop Growers of America 2014). Over the past 5 yr, this specialty crop has resurged in the northeast, fueled by the local food movement. In Vermont alone, >40 breweries are seeking local ingredients including but not limited to hops. As acreage increases, regionally specific pest challenges have become apparent and growers look to extension services for relevant scouting and management strategies.
Integrated pest management (IPM) programs are crop, pest, and region specific. Arthropod communities are a reflection of climate, landscape, and management practices (Schweiger et al. 2005). There are clear differences in climate and landscape between the traditional Pacific Northwestern hop-growing region and the rejuvenated northeast. Therefore, it is important that northeast hop IPM is developed to respond to regional arthropod community abundance and phenology.
Twospotted spider mite (Tetranychus urticae Koch) (Acari: Tetranychidae) and hop aphid (Phorodon humuli (Schrank)) (Hemiptera: Aphididae) have been documented and researched extensively as economically damaging arthropod pests in the Pacific Northwest and European hop-growing regions (James 2003a,b; Mahaffee et al. 2009; Woods et al. 2014). To our knowledge, arthropod communities in hop yards were last reported for the northeast region in the 1940s as reports from the Cornell Agricultural Experiment Station that document hop aphid, twospotted spider mite, and potato leafhopper (Empoasca fabae (Harris)) (Hemiptera: Cicadellidae) as major arthropod pests in New York hop yards (Magie 1944).
Communities of predators and parasitoids can contribute to pest management in agricultural landscapes (Symondson et al. 2002). Assemblages of generalist and specialist natural enemy arthropods reduce pest abundance in hop yards (Campbell and Cone 1994; Gardiner et al. 2003; James 2003a,b; Gent et al. 2009a,b; Grasswitz and James 2009, 2011; Woods et al. 2014).
In an effort to provide northeast hop growers and stakeholders with current pest management strategies, it was important to first identify predominant pests and subsequent natural enemies in Vermont hop yards. The objectives of this publication were to 1) present current hop pest biology and management tactics, 2) report the phenology of arthropod pests observed over three growing seasons (2012–2014), 3) report abundance and peak date for each pest, and 4) document natural enemy abundance.
Biology and Management of Major Northeastern Hop Pests
Twospotted Spider Mite
Twospotted spider mites overwinter in the crown of hop plants, woody debris, and trellis pole crevasses as diapausing adult females. As soon as temperatures warm in the spring, females emerge, migrate to new growth, feed, and lay up to 16 eggs per day. Five to eight generations of spider mite are observed during the hop-growing season in the Pacific Northwest (Mahaffee et al. 2009). Twospotted spider mites are a midlate season pest that thrives in hot and dry environmental conditions. Spider mites develop from egg to adult in 7–10 d at 28–30°C (Mahaffee et al. 2009). Scouting for spider mites on the underside of leaves is an important practice given the potential for rapid population increase in the right conditions (Weihrauch 2004).
Symptoms
Twospotted spider mites are small and translucent. Eggs, larvae, and nymphs are white to clear in color while adults can appear white to yellow in color with two dark spots on the back (Fig. 1). Females are the largest life stage of this mite with a length of ∼0.5 mm. Males are ∼0.2 mm in length. This pest feeds with piercing–sucking mouthparts on leaf and cone mesophyll cells. The first sign of damage is “stippling,” pin-prick sized brown spots on leaves (Fig. 2). Adults and eggs first appear on the underside of leaves in the space between leaf veins. As the population grows, webbing will appear and brown stippling will expand, eventually turning whole leaves brown as a result of desiccation. Foliar injury reduces plant vigor while spider mite feeding on cones can drastically reduce the marketability of the hop product via desiccation which causes cones to shatter (Mahaffee et al. 2009).

Twospotted spider mite, Tetranychus urticae Koch, adults, nymphs, and eggs.

Characteristic twospotted spider mite “stippling” damage at a moderate mite infestation level.
Management
Pesticide application is the traditional management tactic used to control twospotted spider mite populations below economically damaging levels in hop production. Spray applications are based on economic thresholds ranging from 1–2 mites per leaf in June and 5–10 mites per leaf in July in Washington State (Strong and Croft 1995) to 60 mites per leaf in German hop yards (Wright and Cone 1999, Weihrauch 2004). Economically damaging levels of twospotted spider mite can be a repercussion of spraying broad-spectrum insecticides and fungicides (James 2002,,2003a,b; Gent et al. 2009a,b; Woods et al. 2012). Twospotted spider mite is a pest of hops whether broad-spectrum insecticides are sprayed or not. However, economic damage to hop yield and quality from this pest is seen with the application of pesticides aimed at reducing other arthropod pests. For example, spring application of imidacloprid is a successful method of reducing hop aphid populations, but natural enemies of hop aphid and twospotted spider mite are also eliminated. This provides an opportunity for secondary pests (twospotted spider mite) to reach uncontrollable outbreak levels (James and Price 2002; James 2003a,b). Hormoligosis, increased female fecundity, has been observed in spider mites when exposed to imidacloprid insecticides (Sclar et al. 1998, James and Price 2002, Ako et al. 2004). Resident natural enemy communities are able to regulate spider mite populations in the absence of pesticides on hops (Huffaker et al. 1969, James et al. 2001, Woods et al. 2014). Due to twospotted spider mite product resistance, research on spider mite pest management in commercial production is evolving toward the conservation of antagonistic arthropods and narrow-spectrum acaricides (James 2003a,b).
Hop cultivars have different susceptibility levels to twospotted spider mites and high farnesol concentration in leaves has been suggested as a possible mechanism behind their preference (Regev and Cone 1975). Peters and Berry (1980) report variation in the density of leaf trichomes on different hop cultivars. It was observed that twospotted spider mite development time was higher on leaves with more dense leaf trichomes (Peters and Berry 1980).
Spider mite populations increase in dry, dusty conditions. Ground cover between hop rows increases humidity, reduces dusty conditions, and therefore reduces favorable twospotted spider mite habitat. Flowering vegetation increases moisture and provides shelter and alternative food resources for predators of twospotted spider mite (Grasswitz and James 2009, Lu et al. 2014). Spider mites can be controlled by an assemblage of natural enemy arthropods, viruses, and pathogens (Jeppson et al. 1975); Stethorus punctum spp. (Coleoptera: Coccinellidae) (James 2003a), Orius spp. (Hemiptera: Anthocoridae), and predatory mites (Mesostigmata: Phytoseiidae) have been identified as effective predators of this pest commonly found in hop yards (Grasswitz and James 2009).
Damson-Hop Aphid
Cool, wet growing seasons are favorable for the damson-hop aphid. Hop aphid alternates between hop as the summer host and Prunus spp. (Rosales: Prunaceae) as winter hosts. Hop aphids have a holocyclic life cycle where adult, winged females (alate) give birth to live nymphs which develop into wingless (apterous) males and females in addition to alate female reproductive adults. Hop aphids overwinter as eggs on Prunus spp. and adult, wingless females hatch in the spring, laying up to four generations of apterous females and alates (Mahaffee et al. 2009). Recently hatched alatae fly from Prunus spp. to hop in early spring when temperatures reach 13°C. Up to 10 generations of apterous hop aphid on hop per season have been observed in the Pacific Northwest depending on weather conditions and management practices. Toward the end of the season, apterous females produce alates which return to Prunus spp. in the fall. They lay alate males and females which subsequently lay overwintering eggs (Wright et al. 2005, Mahaffee et al. 2009).
In Spain, peak hop aphid date is typically in mid-July (Lorenzana et al. 2013), while in Washington State peak populations are observed in late August and continue into early September (Campbell and Cone 1994). The abundance of alternate Prunus spp. hosts in European farm landscapes has been observed to increase the number of migrant female aphids into hop yards in comparison to migrant populations in Washington state where there are fewer surrounding Prunus spp. (Campbell and Cone 1994). While foliar feeding can reduce plant productivity at high population levels, the moist habitat that hop aphids provide for sooty mold to grow in cones, reduces hop yield and quality.
Symptoms
These soft bodied, pear-shaped insects are found on the underside of hop leaves and range in color from white to light green. Hop aphids are often found on the upper and lower leaf surfaces of new hop growth (Fig. 3). Hop aphids have piercing–sucking mouthparts which are used to feed on the phloem of hop plants. On hop, aphids are smaller than on Prunus spp, the winter host plant. Hop aphids secrete a sugary substance commonly referred to as “honeydew” that provides the perfect medium for growth of sooty mold, an Ascomycete fungus, to grow in hop cones (Wright et al. 2005, Mahaffee et al. 2009, Lorenzana and Hermoso 2010). With high aphid population levels during cone development and at harvest time, fuzzy looking grey-black colored mold can be found when hop cones are pulled apart. Sooty mold is the indirect impact that aphids have on hop quality (Fig. 4).

Hop aphids on new hop growth.

Hop cone infested with hop aphids and sooty mold.
Management
Hop aphid population ecology has been studied, and economic thresholds have been determined (Campbell 1977, Wright et al. 2005, Lorenzana et al. 2009). A study conducted in Spain recommends holding off insecticides application against hop aphid until an economic threshold of 8–10 hop aphids per leaf is reached (Lorenzana et al. 2009). An economic threshold specific to the northeast region for hop aphids does not exist. Biological control of hop aphid including the release of parasitoids (Wright and James 2001) and predators (Campbell and Cone 1994) has been described as an effective management tool. Hop aphids are reported to thrive on plant parts highest in nitrogen and in hop yards with ample nitrogen application (Gent et al. 2009a). Hop plants are fast growing and reach a height of 5 m in one growing season (Neve 1991). The large flush of new vegetative growth in hop yards serves as a food source for hop pests and aphids in particular.
Hop cultivars vary in their susceptibility to hop aphid (Campbell 1983, Dorschner and Baird 1988, Weihrauch and Moreth 2005). ‘Cascade’ has been reported as highly susceptible to hop aphid in the Pacific Northwest (Dorschner and Baird 1988). The mechanisms behind the variation in hop aphids by cultivar are likely a combination of plant nutrition and leaf chemistry. Cultivars with high essential oil content are more attractive to hop aphids. In particular, high levels of cariophyllene and farnesene have been highlighted as attractive essential oils to hop aphids (Kralj et al. 1998).
Potato Leafhopper
Potato leafhopper has not been reported or studied on hops since the 1940s (Magie 1944). A native to the northeast, this pest has been researched in other northeastern production systems including snap bean (Phaseolus vulgaris) (Nault et al. 2004), alfalfa (Medicago sativa) (Lamp et al. 2007), maple (Acer spp.) (Frank et al. 2007), and grape (Vitis vinifera) (Lenz et al. 2012). Potato leafhopper is a polyphagous, migratory insect with an appetite for >200 broad leaf plants (DeLong 1971). Adult females can arrive to northern Vermont anytime between May and August, making it an unpredictable pest. Adults overwinter in southeastern states as far north as Virginia (Taylor and Shields 1995). Spring wind currents carry this species of leafhoppers north. Typically, adult females arrive to Midwestern states first and northeastern states last (Sidumo et al. 2005).
Upon arrival, females feed and lay eggs in hop leaf and stem tissue. Potato leafhoppers can develop at temperatures between 10 and 24°C (Simonet and Pienkowski 1980, Sher and Shields 1991). On alfalfa, nymphs hatch between 3 and 10 d after oviposition. Wingless nymphs go through five instars over the course of 10–14 d before finally molting into an winged adult. In another 7–10 d, females will begin oviposition (DeLong 1971). On an average, it takes 3 wk for an egg to develop into an adult (Hogg 1985). Depending on spring arrival time and temperature, potato leafhoppers will have two or three generations per season at northern latitudes.
Symptoms
Potato leafhoppers are light green, wedge-shaped insects. Adults are 3.0 mm in length while first-instar nymphs can be 0.5 mm long (Fig. 5). Injury from this pest is called “hopperburn” and described as “V”-shaped leaf chlorosis where the outer edges and tip turn yellow and can develop further into brown leaf necrosis (Fig. 6). Potato leafhoppers feed with piercing–sucking mouthparts on mesophyll cells or phloem depending on the plant host (DeLong 1971). Either the leafhopper or the plant immune system restricts phloem flow to the leaf edges resulting in leaf edge yellowing and curling. Visual hopperburn is not present until after significant leafhopper feeding has occurred. It is documented that these symptoms are the result of host plant physiological response to potato leafhopper feeding as exhibited in alfalfa (Lamp et al. 2007) and grape (Lamp et al. 2011, Lenz et al. 2012). Adult potato leafhopper feeding on first-year hop leaves was shown to reduce net photosynthesis of eight hop cultivars tested in 2014 (L. B. Calderwood, unpublished data).

Potato leafhopper, Empoasca fabae (Harris), nymphs of different first, third, fourth, and fifth instars on the underside of a hop leaf. Small yellow spots on leaves are lupulin glands, not arthropods.

Leaf chlorosis and necrosis damage caused by potato leafhopper feeding called “hopper burn.”
Management
Potato leafhopper IPM programs involve monitoring of the population weekly. In alfalfa, sweep netting and sticky traps are used to assess leafhopper populations (Degooyer et al. 1998). In hops, scouting the underside of several leaves from each cultivar is recommended. Potato leafhoppers are visible with the naked eye and have a signature side-to-side scuttling movement. There is evidence that copper sulfate fungicides (Bordeaux mixture), primarily used for hop downy mildew control, were indirectly used to manage potato leafhopper populations in the 1800s (Magie 1944). Copper hydroxide-based sprays are currently the most widely used products to manage hop downy mildew in the northeast. As mentioned above, the impact of fungicides on twospotted spider mite, hop aphid, and natural enemies has been investigated in the Pacific Northwest (James 2002,,2003a,b; Gent et al. 2009a,b; Woods et al. 2012). The natural enemy assemblage for potato leafhopper is similar to that of hop aphid (Table 1), and spraying for potato leafhopper, another soft-bodied insect, is likely to have similar secondary outbreak repercussions.
Major hop pests and their natural enemy groups
| Pest common name | Pest species | Natural enemy common name | Natural enemy taxon |
|---|---|---|---|
| Twospotted spider mite | Tetranychus urticae | Predatory mites | Phytoseiidae |
| Spider mite destroyer | Stethorus punctum spp. | ||
| Minute pirate bug | Anthocoridae | ||
| Hop aphid | Phorodon humuli | Minute pirate bug, bigeyed bugs, and damsel bugs | Anthocoridae, Geocoridae, Nabidae |
| Green lacewing and brown lacewing | Chrysopidae and Hemerobiidae | ||
| Ladybird beetles | Coccinellidae | ||
| Parasitoid wasps | Parasitica | ||
| Syrphid fly | Syrphidae | ||
| Potato leafhopper | Empoasca fabae | Minute pirate bug, bigeyed bugs, and damsel bugs | Anthocoridae, Geocoridae, Nabidae |
| Green lacewing and brown lacewing | Chrysopidae and Hemerobiidae | ||
| Ladybird beetles | Coccinellidae | ||
| Parasitoid wasps | Parasitica | ||
| Spiders | Aranea |
| Pest common name | Pest species | Natural enemy common name | Natural enemy taxon |
|---|---|---|---|
| Twospotted spider mite | Tetranychus urticae | Predatory mites | Phytoseiidae |
| Spider mite destroyer | Stethorus punctum spp. | ||
| Minute pirate bug | Anthocoridae | ||
| Hop aphid | Phorodon humuli | Minute pirate bug, bigeyed bugs, and damsel bugs | Anthocoridae, Geocoridae, Nabidae |
| Green lacewing and brown lacewing | Chrysopidae and Hemerobiidae | ||
| Ladybird beetles | Coccinellidae | ||
| Parasitoid wasps | Parasitica | ||
| Syrphid fly | Syrphidae | ||
| Potato leafhopper | Empoasca fabae | Minute pirate bug, bigeyed bugs, and damsel bugs | Anthocoridae, Geocoridae, Nabidae |
| Green lacewing and brown lacewing | Chrysopidae and Hemerobiidae | ||
| Ladybird beetles | Coccinellidae | ||
| Parasitoid wasps | Parasitica | ||
| Spiders | Aranea |
Major hop pests and their natural enemy groups
| Pest common name | Pest species | Natural enemy common name | Natural enemy taxon |
|---|---|---|---|
| Twospotted spider mite | Tetranychus urticae | Predatory mites | Phytoseiidae |
| Spider mite destroyer | Stethorus punctum spp. | ||
| Minute pirate bug | Anthocoridae | ||
| Hop aphid | Phorodon humuli | Minute pirate bug, bigeyed bugs, and damsel bugs | Anthocoridae, Geocoridae, Nabidae |
| Green lacewing and brown lacewing | Chrysopidae and Hemerobiidae | ||
| Ladybird beetles | Coccinellidae | ||
| Parasitoid wasps | Parasitica | ||
| Syrphid fly | Syrphidae | ||
| Potato leafhopper | Empoasca fabae | Minute pirate bug, bigeyed bugs, and damsel bugs | Anthocoridae, Geocoridae, Nabidae |
| Green lacewing and brown lacewing | Chrysopidae and Hemerobiidae | ||
| Ladybird beetles | Coccinellidae | ||
| Parasitoid wasps | Parasitica | ||
| Spiders | Aranea |
| Pest common name | Pest species | Natural enemy common name | Natural enemy taxon |
|---|---|---|---|
| Twospotted spider mite | Tetranychus urticae | Predatory mites | Phytoseiidae |
| Spider mite destroyer | Stethorus punctum spp. | ||
| Minute pirate bug | Anthocoridae | ||
| Hop aphid | Phorodon humuli | Minute pirate bug, bigeyed bugs, and damsel bugs | Anthocoridae, Geocoridae, Nabidae |
| Green lacewing and brown lacewing | Chrysopidae and Hemerobiidae | ||
| Ladybird beetles | Coccinellidae | ||
| Parasitoid wasps | Parasitica | ||
| Syrphid fly | Syrphidae | ||
| Potato leafhopper | Empoasca fabae | Minute pirate bug, bigeyed bugs, and damsel bugs | Anthocoridae, Geocoridae, Nabidae |
| Green lacewing and brown lacewing | Chrysopidae and Hemerobiidae | ||
| Ladybird beetles | Coccinellidae | ||
| Parasitoid wasps | Parasitica | ||
| Spiders | Aranea |
Potato leafhoppers are known to be repelled by glandular trichomes produced at different densities on alfalfa (Shockley and Backus 2002), potato (Kaplan et al. 2008), and dry bean (Gonzales et al. 2004) cultivars. Preliminary data indicated that some hop cultivars were more susceptible to potato leafhopper feeding than others. Additionally, first-year hop plants have been observed to be particularly susceptible to potato leafhopper feeding injury (unpublished data). Further research should identify which hop cultivars are more and less susceptible to this emerging pest and explore mechanisms behind cultivar susceptibility.
Hop Downy Mildew
Although hop downy mildew incidence was not included in this arthropod survey, the biology and management of this pathogen are of great importance, as it is a major challenge in northeastern hop yards. The pathogen has been positively confirmed in numerous yards in NY, MA, and VT by the UVM diagnostic lab and at times by Dr. David Gent of Oregon State University. Hop downy mildew is caused by the oomycete pathogen Pseudoperonospora humuli. Spring in the northeast is often cool and wet providing a perfect habitat for the spread of hop downy mildew. The pathogen overwinters as mycelium in ground leaf litter and on hop crowns (Skotland and Johnson 1983). Research on eastern downy mildew strains is of particular need. It is unclear weather oospores are an overwintering stage of the disease cycle in eastern states (Magie 1942, Skotland and Johnson 1983). Given moist conditions in the spring, sporangia are carried by wind and rain containing zoospores, the primary inoculum. Zoospores enter hop leaves through leaf stomata, germinate, and produce more sporangia, which release secondary inoculum zoospores that infect additional plants. Hop downy mildew zoospores can arrive to a hop yard via wind currents or in already infected rootstock. The crowns that harbor zoospores and oospores over the winter and give rise to infected shoots in the spring, have systemic downy mildew. These shoots are called primary basal spikes (Fig. 7) because they release the first inoculum of the season. When downy mildew is systemic, the pathogen lives in the hop yard year round and will continue to spread unless meticulously managed.

Hop downy mildew primary basal spike with short internodes, chlorosis, and necrosis.
Symptoms
Early spring (March–May) pale green-yellow shoots with short internodes are primary basal spikes. Secondary inoculum infects already trained bines in late spring forming areal spikes. Both basal and areal spikes have a “Christmas tree” appearance in comparison to healthy hop bines (Figs. 7 and 8). Chlorotic, stunted bines that fall away from strings are also a symptom of secondary inoculum. Foliar lesions are present on the leaves of both basal and areal spikes. Lesions are sections of leaf cells that form angled brown spots on the underside of leaves (Fig. 9). On severely infected spikes, the entire underside of leaves will be covered in brown sporangia producing more zoospores (Mahaffee et al. 2009). This hop-specific disease reduces hop moisture content and impacts the appearance of hop cones which directly reduces the quality of hops delivered to brewers. Hop cones infected with downy mildew are prone to early ripening, browning, uneven drying in the oast, a shorter shelf life, and less desirable brewing characteristics, all of which affect the marketability of the product (Skotland and Johnson 1983, Mahaffee et al. 2009). Fungicide sprays can cause phytotoxicity evidenced by spotting on leaves (Fig. 10) in hot and humid weather conditions. Severe fungicide burn can result in plant desiccation. Scouting observations indicate that certain hop cultivars are more susceptible to copper-based fungicides than others.

Hop downy mildew areal spike with short internodes and chlorosis.

Hop downy mildew foliar lesions on the underside of a hop leaf.

Phytotoxicity from application of copper-based fungicide.
Management
It is difficult to reduce disease in an already infested hop yard. A combination of resistant cultivars, hop yard sanitation, and fungicides are used to reduce infestation levels of hop downy mildew. No hop cultivar is currently 100% resistant. When the more susceptible cultivars contract the disease, the resistant cultivars are exposed and remain subject to economic losses from reduced yield and cone quality. The most resistant cultivars include Fuggle, Newport, Perle, Spalter, Wye Challenger, Hallertauer Gold, Hallertauer Magnum, and Hallertauer Tradition (Gent et al. 2009b).
“Crowning” or spring pruning is an important spring management practice where late winter or early spring crown growth is mechanically or chemically removed. In the Pacific Northwest, Gent et al. (2012) provides evidence for the reduction in hop downy mildew infection. Hop yards that were pruned twice showed the lowest disease severity and growers were able to apply one fewer fungicide application. Crowning date and number influenced yield of ‘Willamette’ in this study (Gent et al. 2012). Research is needed on the impact of crowning on harvest date and yield for the shorter growing season of the northeast region. Later maturing cultivars in Europe have reduced yield when cut back at later dates (Goenia and Micibski 1972). After crowning, scouting for basal spikes should take place. If basal spikes are present they should be removed from the hop yard immediately. Sanitation of hop yard clippers and pruning tools is a critical and easy way to reduce the risk of disease spread (Skotland and Johnson 1983, Gent et al. 2012).
Pruning must be paired with fungicide applications for hop downy mildew management (Hunger and Horner 1982, Mahaffee et al. 2009). A variety of products and application times are currently exercised in northeastern hop yards in hopes of controlling this disease. According to our 2014 pesticide survey, which included grower responses from Vermont, New York, and Massachusetts, the most commonly sprayed fungicides are copper based. A mean of four fungicide applications are sprayed per season with some farms 12 applications of a copper-based fungicide per season. In Europe, 10–16 fungicide applications are common practice. It is clear from research in Washington and Oregon that making the timing of fungicide sprays more precise can reduce the number of fungicide applications per season. Using a forecasting system to calculate emergence of the first systemically infected spikes of downy mildew reduced the amount of fungicide sprayed in the Pacific Northwest without increasing disease severity (Gent et al. 2010).
Natural Enemy Arthropods of Importance
Parasitoid Wasps
Parasitic hymenopterans are very small wasps abundant in diverse agricultural landscapes. Several species are known parasitoids of potato leafhopper and hop aphid and their presence in hop yards has been documented (Wright and James 2001, Grasswitz and James 2009). Anagrus spp. are known for their parasitism of potato leafhopper eggs (Lovinger et al. 2000) while a suite of additional parasitoid species play a role in landscape biocontrol. Assemblages of several parasitoid species have been shown to keep aphid populations in check (Sigsgaard 2002). The effectiveness of parasitoids as biological control agents in hops has not been studied (Grasswitz and James 2009). However, Wright and James (2001) reared 802 parasitoids from 83 hop aphids collected on Prunus spp. (alternative hop aphid host) in Washington State. The most abundant species reared was Lysiphlebus testaceipes (Cresson). Grasswitz and James (2011) identified nine generalist parasitoid species in Pacific Northwest hop yards when assessing parasitism on hop looper (Hypena humuli Harris), a minor lepidopteran pest of hops. The parasitoids found in this study were not identified to species.
Spiders and Predatory Mites
There are over 30,000 species of spider. This diverse group of arthropods primarily feeds on other arthropods, making them effective members of predatory guilds (Riechert and Lockley 1984). A diversity of these generalist predators are present in healthy agricultural ecosystems. In alfalfa, spiders are highlighted as predators of potato leafhopper (Harwood and Obrycki 2007). Predatory mites (Acari: Phytoseiidae) including Phytoseiulus persimilis Athias-Henriot, Neoseiulus californicus McGregor, Neoseiulus fallacis Garman, Amblyseius andersoni (Chant), Galendromus occidentalis Nesbitt have controlled twospotted spider mite populations in Pacific Northwest (James 2002,,James et al. 2003) and United Kingdom (Barbour et al. 2003) hop yards. N. fallacis has been shown to manage spider mites in eastern orchard systems (Metzger 2001). Similar to spiders, predatory mites and spider mites can be seen with the naked eye. Additional steps should be taken to measure predatory mite presence and the potential impact they may have on spider mite populations in nontraditional hop-growing regions.
Predatory True Bugs and Flies
In the minute pirate bug family (Hemiptera: Anthocoridae), the Orius genus is reported as an effective generalist predator of hop aphids (Lorenzana and Hermoso 2010). According to studies in alfalfa, minute pirate bugs are effective predators of potato leafhopper (Wieser Erlandson and Obrycki 2010). Damsel bugs (Nabidae) and big eyed bugs (Geocoridae) are present in Pacific Northwest hop yards (Grasswitz and James 2011) and are observed occasionally in northeastern hop yards. Spined soldier bugs (Pentatomidae: Podisus spp.) have been spotted in northeastern hop yards and have been shown to be effective generalist biocontrol agents against Lepidopteran pests of cotton (Lopez et al. 1976) and tomato (De Clercq et al. 1998). Syrphid fly larvae are specifically known as effective biocontrol agents against aphids (Diptera: Syrphidae) (Bugg 1992). Finally, species of predatory midge, Aphidoletes spp. (Diptera: Cecidomyiidae), are shown to be effective members of the aphid biological control community in temperate apple orchards. Aphid midges thrive in cool, moist conditions and are potentially abundant in northeastern hop yards (Wyss et al. 1999, Minarro et al. 2005).
Lacewings
Green lacewings are effective consumers of soft-bodied insects in Pacific Northwest hop yards (James 2006, Lorenzana et al. 2013). James (2006) demonstrated that goldeneyed lacewing (Chrysopa oculata Say) populations were increased with the placement of methyl salicylate-baited sticky cards. Lacewing larvae were observed to feed on four potato leafhoppers per day at high pest densities under lab conditions (Weiser Erlandson and Obrycki 2010).
Lady Beetles
The Coccinellidae family of beetles includes many generalist predators. Hippodamia convergens Guérin-Méneville, Harmonia axyridis Pallas, and the native Coleomegilla maculata De Geer are three predatory lady beetle species found in hop yards (James 2003a, Campbell and Cone 1994). Lady beetles are well known for their consumption of soft-bodied arthropod pests, and certain species are mass reared for release in some farm landscapes. Lady beetle larvae have a voracious appetite for aphids and will consume more individual pests than the adult life stage (Koch 2003). H. axyridis has been reported to feed on twospotted spider mites and hop aphids in hop yards (James 2003b). C. maculata are specifically reported to feed on potato leafhoppers in alfalfa systems. The Stethorus punctum picipes (Casey), commonly known as the spider mite destroyer, is a small lady beetle that feeds on spider mites in the west (James 2003a). Stethorus punctum punctum (LeConte) is the spider mite destroyer of the eastern United States and is conserved for biological control in apple production (Felland and Hull 1996).
Arthropod Scouting Methodology
Seven hop yards in Vermont were scouted every other week June–August for 3 yr (2012–2014; Fig. 11). In 2012, there were few hop yards in Vermont and those selected for this survey were growers willing to participate. Hop yards varied in acreage, cultivar diversity, and management practices. As more farmers started growing hops in Vermont the number of hop yards scouted increased in 2013 and 2014 (Table 2).
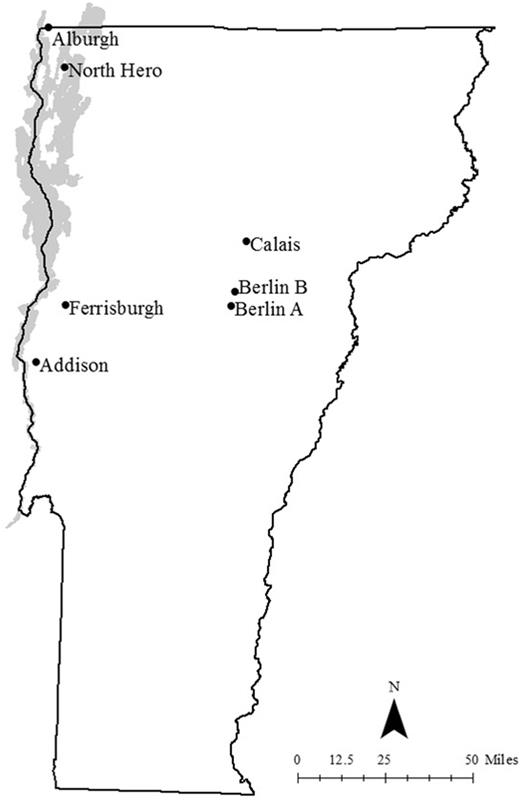
Hop yard scouting sites in Vermont. The “North” micro region includes Alburgh and North Hero yards, “West” includes Addison and Ferrisburgh yards, and “East” includes Calais, Berlin A, and Berlin B yards.
Characteristics of hop yards scouted in Vermont
| Farm | Years scouted | No. of cultivars | Farm sizea | Micro region | Management |
|---|---|---|---|---|---|
| Addison | 2012, 2013, 2014 | 7 | Large | West | Organicb |
| Alburgh | 2012, 2013, 2014 | 24 | Large | North | Organic |
| Berlin A | 2012, 2013, 2014 | 3 | Medium | East | Nonorganicc |
| Calais | 2013, 2014 | 1 | Small | East | Organic practiced |
| North Hero | 2013, 2014 | 4 | Small | North | Organic practice |
| Berlin B | 2014 | 5 | Medium | East | Nonorganic |
| Ferrisburgh | 2014 | 5 | Large | West | Nonorganic |
| Farm | Years scouted | No. of cultivars | Farm sizea | Micro region | Management |
|---|---|---|---|---|---|
| Addison | 2012, 2013, 2014 | 7 | Large | West | Organicb |
| Alburgh | 2012, 2013, 2014 | 24 | Large | North | Organic |
| Berlin A | 2012, 2013, 2014 | 3 | Medium | East | Nonorganicc |
| Calais | 2013, 2014 | 1 | Small | East | Organic practiced |
| North Hero | 2013, 2014 | 4 | Small | North | Organic practice |
| Berlin B | 2014 | 5 | Medium | East | Nonorganic |
| Ferrisburgh | 2014 | 5 | Large | West | Nonorganic |
a Small yards had <60 hills, medium yards had 60–200 hills, and large yards had >200 hills.
b Hop yards certified organic by Vermont Organic Farmers, LLC, Richmond, VT were considered “Organic.”
c Hop yards that were not certified organic were considered “Nonorganic.”
d Hop yards that were not certified but follow organic management were considered “Organic Practice.”
Characteristics of hop yards scouted in Vermont
| Farm | Years scouted | No. of cultivars | Farm sizea | Micro region | Management |
|---|---|---|---|---|---|
| Addison | 2012, 2013, 2014 | 7 | Large | West | Organicb |
| Alburgh | 2012, 2013, 2014 | 24 | Large | North | Organic |
| Berlin A | 2012, 2013, 2014 | 3 | Medium | East | Nonorganicc |
| Calais | 2013, 2014 | 1 | Small | East | Organic practiced |
| North Hero | 2013, 2014 | 4 | Small | North | Organic practice |
| Berlin B | 2014 | 5 | Medium | East | Nonorganic |
| Ferrisburgh | 2014 | 5 | Large | West | Nonorganic |
| Farm | Years scouted | No. of cultivars | Farm sizea | Micro region | Management |
|---|---|---|---|---|---|
| Addison | 2012, 2013, 2014 | 7 | Large | West | Organicb |
| Alburgh | 2012, 2013, 2014 | 24 | Large | North | Organic |
| Berlin A | 2012, 2013, 2014 | 3 | Medium | East | Nonorganicc |
| Calais | 2013, 2014 | 1 | Small | East | Organic practiced |
| North Hero | 2013, 2014 | 4 | Small | North | Organic practice |
| Berlin B | 2014 | 5 | Medium | East | Nonorganic |
| Ferrisburgh | 2014 | 5 | Large | West | Nonorganic |
a Small yards had <60 hills, medium yards had 60–200 hills, and large yards had >200 hills.
b Hop yards certified organic by Vermont Organic Farmers, LLC, Richmond, VT were considered “Organic.”
c Hop yards that were not certified organic were considered “Nonorganic.”
d Hop yards that were not certified but follow organic management were considered “Organic Practice.”
Scouting took place every other week for a total of six collection dates at each location annually. A group of five to seven plants was considered a plot. Three leaves on one plant in each plot were sampled during each visit between ground level and 2 m. Both top and bottom leaf surfaces were visually examined with Optivisor lenses (Donegan Optical Company Inc., Lenexa, KS). Arthropods were identified and counted in the field. Pests were identified to species level, while natural enemies were identified to the level at which an ecological role could be assigned (Table 1). Parasitoid wasps and predatory mites were not reported. Statistical comparisons were not presented because it was the goal to show community variation by location and management practices. The data presented are a representation of the arthropod community found in northeastern hop yards. Season sums and peak date means were calculated for arthropod taxons.
Beyond farm location, variables impacting arthropod presence including farm size, management practice, and microclimate region were assigned to each farm. Small, medium, or large size was also assigned to each farm. Small yards had <60 hills (<120 plants), medium yards had 60–200 hills (120–400 plants), and large yards had >200 hills (>400 plants). “Organic” yards were certified organic (Vermont Organic Farmers, LLC, Richmond, VT) while “Nonorganic” yards practiced a range of management tactics between organic but not certified, and conventional and were not certified organic. The “North” micro region included Alburgh and North Hero farms, “West” included Addison and Ferrisburgh farms, and “East” includes Calais, Berlin A, and Berlin B farms.
Scouting Results
Larger yards had a higher total number of pest and natural enemy individuals. However, there were hot spots of pests present on small- and medium-sized yards. With higher numbers of pests, the natural enemy community increased in abundance. Aphid and potato leafhopper natural enemy assemblages were similarly composed of generalist predators. The “East” micro region had fewer growing degree-days each year of the study. Locations with higher elevations included Calais (338 m) and Berlin (268 m). Alburgh, North Hero, Addison, and Ferrisburgh are at elevations between 27 m and 32 m along the large, freshwater Lake Champlain. Overall, Calais and Berlin yards had lower pest levels, yet they are small- and medium-sized yards.
Twospotted Spider Mite
Twospotted spider mite populations were highest in 2012 and 2014. The number of spider mite generations cannot be concluded from these data, but there is an overall late season population increase in all hop yards. In 2012, the spider mite peak date was in early August in all locations. North and West locations had a similar early August peak date in 2014 while East locations exhibited twospotted spider mite peaks in late August (Table 3). Peak dates in Table 6 indicate that spider mite destroyer populations mirrored or lagged behind twospotted spider mite populations. This pattern was especially clear in large yards and in years with high twospotted spider mite populations (Table 4). In Addison, the decreased ratio of twospotted spider mites to spider mite destroyers from 2012 to 2014 indicates that this predator population can become established. Without considering other yard variables, spider mite destroyers appear to have reduced the 2014 twospotted spider mite population in that location (Table 4, Fig. 12). Addison, being a large yard, had fewer twospotted spider mites than Berlin B, a medium-sized yard. Ferrisburgh had the highest total number of twospotted spider mites and low natural enemy abundance in 2014.
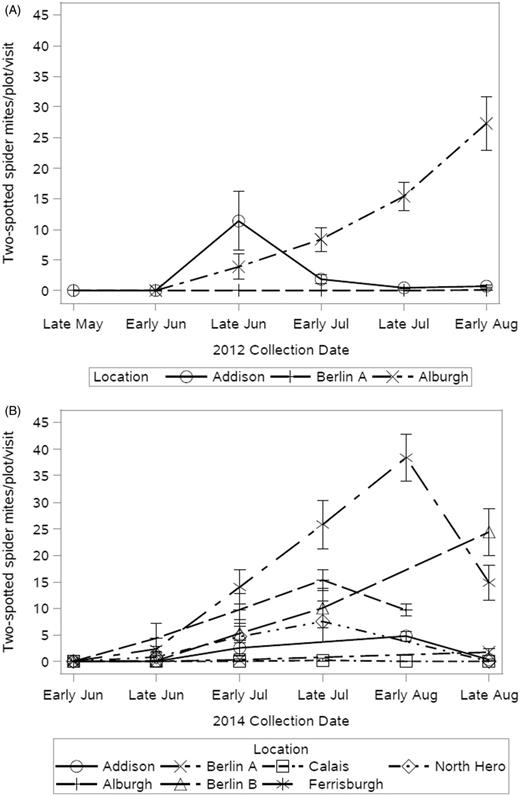
Mean number of twospotted spider mites counted per plot, per yard visit in 2012 (A) and 2014 (B). Three farms were scouted in 2012 and seven in 2014. No spider mites were observed in 2014.
Twospotted spider mite (TSSM) season sum, peak date, and peak date mean (± SEM) by year and farm
| TSSM | Farm | Farm size | Micro region | Season sum | Peak date | Peak date mean (± SEM) |
|---|---|---|---|---|---|---|
| 2012 | Alburgh | Large | North | 3,207 | 8-Aug. | 27.32 (± 4.38) |
| Addison | Large | West | 809 | 6-Aug. | 0.70 (± 0.17) | |
| Berlin A | Medium | East | 4 | 7-Aug. | 0.13 (± 0.13) | |
| 2013 | Alburgh | Large | North | 145 | 12-Aug. | 2.00 (± 0.34) |
| North Hero | Small | North | 0 | N/A | 0.00 | |
| Addison | Large | West | 172 | 15-July | 0.91 (± 0.23) | |
| Calais | Small | East | 0 | N/A | 0.00 | |
| Berlin A | Medium | East | 0 | N/A | 0.00 | |
| 2014 | Alburgh | Large | North | 3,206 | 4-Aug. | 14.58 (± 1.82) |
| North Hero | Small | North | 209 | 21-July | 7.50 (± 3.85) | |
| Addison | Large | West | 623 | 6-Aug. | 4.77 (± 0.90) | |
| Ferrisburgh | Large | West | 3,609 | 6-Aug. | 38.39 (± 4.34) | |
| Calais | Small | East | 2 | 25-July | 0.25 (± 0.25) | |
| Berlin A | Medium | East | 63 | 22-Aug. | 1.80 (± 0.69) | |
| Berlin B | Medium | East | 1,046 | 22-Aug. | 24.38 (± 4.50) |
| TSSM | Farm | Farm size | Micro region | Season sum | Peak date | Peak date mean (± SEM) |
|---|---|---|---|---|---|---|
| 2012 | Alburgh | Large | North | 3,207 | 8-Aug. | 27.32 (± 4.38) |
| Addison | Large | West | 809 | 6-Aug. | 0.70 (± 0.17) | |
| Berlin A | Medium | East | 4 | 7-Aug. | 0.13 (± 0.13) | |
| 2013 | Alburgh | Large | North | 145 | 12-Aug. | 2.00 (± 0.34) |
| North Hero | Small | North | 0 | N/A | 0.00 | |
| Addison | Large | West | 172 | 15-July | 0.91 (± 0.23) | |
| Calais | Small | East | 0 | N/A | 0.00 | |
| Berlin A | Medium | East | 0 | N/A | 0.00 | |
| 2014 | Alburgh | Large | North | 3,206 | 4-Aug. | 14.58 (± 1.82) |
| North Hero | Small | North | 209 | 21-July | 7.50 (± 3.85) | |
| Addison | Large | West | 623 | 6-Aug. | 4.77 (± 0.90) | |
| Ferrisburgh | Large | West | 3,609 | 6-Aug. | 38.39 (± 4.34) | |
| Calais | Small | East | 2 | 25-July | 0.25 (± 0.25) | |
| Berlin A | Medium | East | 63 | 22-Aug. | 1.80 (± 0.69) | |
| Berlin B | Medium | East | 1,046 | 22-Aug. | 24.38 (± 4.50) |
Twospotted spider mite (TSSM) season sum, peak date, and peak date mean (± SEM) by year and farm
| TSSM | Farm | Farm size | Micro region | Season sum | Peak date | Peak date mean (± SEM) |
|---|---|---|---|---|---|---|
| 2012 | Alburgh | Large | North | 3,207 | 8-Aug. | 27.32 (± 4.38) |
| Addison | Large | West | 809 | 6-Aug. | 0.70 (± 0.17) | |
| Berlin A | Medium | East | 4 | 7-Aug. | 0.13 (± 0.13) | |
| 2013 | Alburgh | Large | North | 145 | 12-Aug. | 2.00 (± 0.34) |
| North Hero | Small | North | 0 | N/A | 0.00 | |
| Addison | Large | West | 172 | 15-July | 0.91 (± 0.23) | |
| Calais | Small | East | 0 | N/A | 0.00 | |
| Berlin A | Medium | East | 0 | N/A | 0.00 | |
| 2014 | Alburgh | Large | North | 3,206 | 4-Aug. | 14.58 (± 1.82) |
| North Hero | Small | North | 209 | 21-July | 7.50 (± 3.85) | |
| Addison | Large | West | 623 | 6-Aug. | 4.77 (± 0.90) | |
| Ferrisburgh | Large | West | 3,609 | 6-Aug. | 38.39 (± 4.34) | |
| Calais | Small | East | 2 | 25-July | 0.25 (± 0.25) | |
| Berlin A | Medium | East | 63 | 22-Aug. | 1.80 (± 0.69) | |
| Berlin B | Medium | East | 1,046 | 22-Aug. | 24.38 (± 4.50) |
| TSSM | Farm | Farm size | Micro region | Season sum | Peak date | Peak date mean (± SEM) |
|---|---|---|---|---|---|---|
| 2012 | Alburgh | Large | North | 3,207 | 8-Aug. | 27.32 (± 4.38) |
| Addison | Large | West | 809 | 6-Aug. | 0.70 (± 0.17) | |
| Berlin A | Medium | East | 4 | 7-Aug. | 0.13 (± 0.13) | |
| 2013 | Alburgh | Large | North | 145 | 12-Aug. | 2.00 (± 0.34) |
| North Hero | Small | North | 0 | N/A | 0.00 | |
| Addison | Large | West | 172 | 15-July | 0.91 (± 0.23) | |
| Calais | Small | East | 0 | N/A | 0.00 | |
| Berlin A | Medium | East | 0 | N/A | 0.00 | |
| 2014 | Alburgh | Large | North | 3,206 | 4-Aug. | 14.58 (± 1.82) |
| North Hero | Small | North | 209 | 21-July | 7.50 (± 3.85) | |
| Addison | Large | West | 623 | 6-Aug. | 4.77 (± 0.90) | |
| Ferrisburgh | Large | West | 3,609 | 6-Aug. | 38.39 (± 4.34) | |
| Calais | Small | East | 2 | 25-July | 0.25 (± 0.25) | |
| Berlin A | Medium | East | 63 | 22-Aug. | 1.80 (± 0.69) | |
| Berlin B | Medium | East | 1,046 | 22-Aug. | 24.38 (± 4.50) |
Twospotted spider mite (TSSM) and spider mite destroyer (SMD) adult and larvae combined season totals and peak dates
| Year | Farm | Season total | Peak date | ||
|---|---|---|---|---|---|
| TSSM | SMD | TSSM | SMD | ||
| 2012 | Alburgh | 3,207 | 121 | 8-Aug. | 8-Aug. |
| Addison | 809 | 21 | 6-Aug. | 24-July | |
| Berlin A | 4 | 1 | 7-Aug. | 24-July | |
| 2013 | Alburgh | 145 | 2 | 12-Aug. | 24-July |
| North Hero | 0 | 0 | N/A | N/A | |
| Addison | 172 | 28 | 15-July | 15-July, 29-Julya | |
| Calais | 0 | 0 | N/A | N/A | |
| Berlin A | 0 | 2 | N/A | N/A | |
| 2014 | Alburgh | 3,206 | 163 | 4-Aug. | 11-Aug. |
| North Hero | 209 | 35 | 21-July | 18-Aug. | |
| Addison | 623 | 52 | 6-Aug. | 6-Aug. | |
| Ferrisburgh | 3,609 | 36 | 6-Aug. | 6-Aug. | |
| Calais | 2 | 0 | 25-July | N/A | |
| Berlin A | 63 | 0 | 22-Aug. | N/A | |
| Berlin B | 1,046 | 0 | 22-Aug. | N/A | |
| Year | Farm | Season total | Peak date | ||
|---|---|---|---|---|---|
| TSSM | SMD | TSSM | SMD | ||
| 2012 | Alburgh | 3,207 | 121 | 8-Aug. | 8-Aug. |
| Addison | 809 | 21 | 6-Aug. | 24-July | |
| Berlin A | 4 | 1 | 7-Aug. | 24-July | |
| 2013 | Alburgh | 145 | 2 | 12-Aug. | 24-July |
| North Hero | 0 | 0 | N/A | N/A | |
| Addison | 172 | 28 | 15-July | 15-July, 29-Julya | |
| Calais | 0 | 0 | N/A | N/A | |
| Berlin A | 0 | 2 | N/A | N/A | |
| 2014 | Alburgh | 3,206 | 163 | 4-Aug. | 11-Aug. |
| North Hero | 209 | 35 | 21-July | 18-Aug. | |
| Addison | 623 | 52 | 6-Aug. | 6-Aug. | |
| Ferrisburgh | 3,609 | 36 | 6-Aug. | 6-Aug. | |
| Calais | 2 | 0 | 25-July | N/A | |
| Berlin A | 63 | 0 | 22-Aug. | N/A | |
| Berlin B | 1,046 | 0 | 22-Aug. | N/A | |
a SMD was equally high on both dates in Addison.
Twospotted spider mite (TSSM) and spider mite destroyer (SMD) adult and larvae combined season totals and peak dates
| Year | Farm | Season total | Peak date | ||
|---|---|---|---|---|---|
| TSSM | SMD | TSSM | SMD | ||
| 2012 | Alburgh | 3,207 | 121 | 8-Aug. | 8-Aug. |
| Addison | 809 | 21 | 6-Aug. | 24-July | |
| Berlin A | 4 | 1 | 7-Aug. | 24-July | |
| 2013 | Alburgh | 145 | 2 | 12-Aug. | 24-July |
| North Hero | 0 | 0 | N/A | N/A | |
| Addison | 172 | 28 | 15-July | 15-July, 29-Julya | |
| Calais | 0 | 0 | N/A | N/A | |
| Berlin A | 0 | 2 | N/A | N/A | |
| 2014 | Alburgh | 3,206 | 163 | 4-Aug. | 11-Aug. |
| North Hero | 209 | 35 | 21-July | 18-Aug. | |
| Addison | 623 | 52 | 6-Aug. | 6-Aug. | |
| Ferrisburgh | 3,609 | 36 | 6-Aug. | 6-Aug. | |
| Calais | 2 | 0 | 25-July | N/A | |
| Berlin A | 63 | 0 | 22-Aug. | N/A | |
| Berlin B | 1,046 | 0 | 22-Aug. | N/A | |
| Year | Farm | Season total | Peak date | ||
|---|---|---|---|---|---|
| TSSM | SMD | TSSM | SMD | ||
| 2012 | Alburgh | 3,207 | 121 | 8-Aug. | 8-Aug. |
| Addison | 809 | 21 | 6-Aug. | 24-July | |
| Berlin A | 4 | 1 | 7-Aug. | 24-July | |
| 2013 | Alburgh | 145 | 2 | 12-Aug. | 24-July |
| North Hero | 0 | 0 | N/A | N/A | |
| Addison | 172 | 28 | 15-July | 15-July, 29-Julya | |
| Calais | 0 | 0 | N/A | N/A | |
| Berlin A | 0 | 2 | N/A | N/A | |
| 2014 | Alburgh | 3,206 | 163 | 4-Aug. | 11-Aug. |
| North Hero | 209 | 35 | 21-July | 18-Aug. | |
| Addison | 623 | 52 | 6-Aug. | 6-Aug. | |
| Ferrisburgh | 3,609 | 36 | 6-Aug. | 6-Aug. | |
| Calais | 2 | 0 | 25-July | N/A | |
| Berlin A | 63 | 0 | 22-Aug. | N/A | |
| Berlin B | 1,046 | 0 | 22-Aug. | N/A | |
a SMD was equally high on both dates in Addison.
Hop Aphid
Hop aphid populations were highest in 2014. The wet spring of 2013 and continual precipitation throughout 2014 allowed hop aphids to flourish (Table 5, Fig. 13). Two hop aphid cycles were observed in 2014. In years of high population levels, hop aphid peak date ranged from early to late August depending on geographic location. As expected, locations with high total pest numbers had the highest peak date means. Natural enemy counts were highest in locations and years with high hop aphid populations. This indicates that increased natural enemy presence could aid in pest population management. Where predators became established there were lower hop aphid season sums. Specifically in Alburgh, the hop aphid population almost doubled from 2013 to 2014. In both 2013 and 2014, Addison had high aphid populations. In contrast, Ferrisburgh had the highest hop aphid population of all yards with low natural enemy abundance (Table 5). Again, it is difficult to compare hop aphid to potato leafhopper control by natural enemies because these assemblages are very similar (Table 1). Although spiders are not included in the hop aphid natural enemy assemblage, it is possible that these generalist predators feed on hop aphids when other prey is not available. Alburgh had the highest spider count of all locations in 2012 and 2014 when the highest number of aphids was observed.
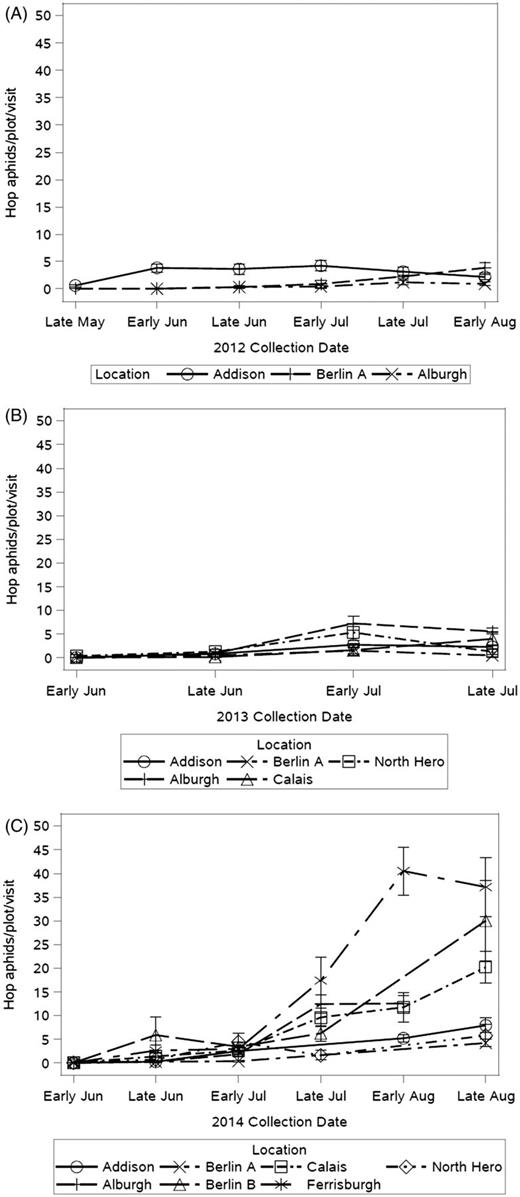
Mean number of hop aphids counted per plot, per yard visit in 2012 (A), 2013 (B), and 2014 (C). Three farms were scouted in 2012, five in 2013, and seven in 2014.
Hop aphid (HA) season sum, peak date, and peak date mean (± SEM) by year and farm
| HA | Farm | Farm size | Micro region | Season sum | Peak date | Peak date mean (± SEM) |
|---|---|---|---|---|---|---|
| 2012 | Alburgh | Large | North | 172 | 25-July | 1.22 (± 0.55) |
| Addison | Large | West | 984 | 24-July | 3.18 (± 0.74) | |
| Berlin A | Medium | East | 224 | 7-Aug. | 3.86 (± 0.95) | |
| 2013 | Alburgh | Large | North | 1562 | 12-Aug. | 12.93 (± 1.60) |
| North Hero | Small | North | 204 | 9-July | 5.35 (± 1.73) | |
| Addison | Large | West | 522 | 29-July | 3.00 (± 0.54) | |
| Calais | Small | East | 93 | 6-Aug. | 5.88 (± 2.13) | |
| Berlin A | Medium | East | 97 | 6-Aug. | 0.80 (± 0.28) | |
| 2014 | Alburgh | Large | North | 2864 | 4-Aug. | 16.93 (± 3.20) |
| North Hero | Small | North | 201 | 18-Aug. | 5.81 (± 2.30) | |
| Addison | Large | West | 1080 | 27-Aug. | 7.96 (± 1.51) | |
| Ferrisburgh | Large | West | 3708 | 6-Aug. | 40.53 (± 5.06) | |
| Calais | Small | East | 365 | 24-Aug. | 20.25 (± 3.39) | |
| Berlin A | Medium | East | 149 | 22-Aug. | 4.27 (± 0.74) | |
| Berlin B | Medium | East | 1131 | 22-Aug. | 30.04 (± 8.50) |
| HA | Farm | Farm size | Micro region | Season sum | Peak date | Peak date mean (± SEM) |
|---|---|---|---|---|---|---|
| 2012 | Alburgh | Large | North | 172 | 25-July | 1.22 (± 0.55) |
| Addison | Large | West | 984 | 24-July | 3.18 (± 0.74) | |
| Berlin A | Medium | East | 224 | 7-Aug. | 3.86 (± 0.95) | |
| 2013 | Alburgh | Large | North | 1562 | 12-Aug. | 12.93 (± 1.60) |
| North Hero | Small | North | 204 | 9-July | 5.35 (± 1.73) | |
| Addison | Large | West | 522 | 29-July | 3.00 (± 0.54) | |
| Calais | Small | East | 93 | 6-Aug. | 5.88 (± 2.13) | |
| Berlin A | Medium | East | 97 | 6-Aug. | 0.80 (± 0.28) | |
| 2014 | Alburgh | Large | North | 2864 | 4-Aug. | 16.93 (± 3.20) |
| North Hero | Small | North | 201 | 18-Aug. | 5.81 (± 2.30) | |
| Addison | Large | West | 1080 | 27-Aug. | 7.96 (± 1.51) | |
| Ferrisburgh | Large | West | 3708 | 6-Aug. | 40.53 (± 5.06) | |
| Calais | Small | East | 365 | 24-Aug. | 20.25 (± 3.39) | |
| Berlin A | Medium | East | 149 | 22-Aug. | 4.27 (± 0.74) | |
| Berlin B | Medium | East | 1131 | 22-Aug. | 30.04 (± 8.50) |
Hop aphid (HA) season sum, peak date, and peak date mean (± SEM) by year and farm
| HA | Farm | Farm size | Micro region | Season sum | Peak date | Peak date mean (± SEM) |
|---|---|---|---|---|---|---|
| 2012 | Alburgh | Large | North | 172 | 25-July | 1.22 (± 0.55) |
| Addison | Large | West | 984 | 24-July | 3.18 (± 0.74) | |
| Berlin A | Medium | East | 224 | 7-Aug. | 3.86 (± 0.95) | |
| 2013 | Alburgh | Large | North | 1562 | 12-Aug. | 12.93 (± 1.60) |
| North Hero | Small | North | 204 | 9-July | 5.35 (± 1.73) | |
| Addison | Large | West | 522 | 29-July | 3.00 (± 0.54) | |
| Calais | Small | East | 93 | 6-Aug. | 5.88 (± 2.13) | |
| Berlin A | Medium | East | 97 | 6-Aug. | 0.80 (± 0.28) | |
| 2014 | Alburgh | Large | North | 2864 | 4-Aug. | 16.93 (± 3.20) |
| North Hero | Small | North | 201 | 18-Aug. | 5.81 (± 2.30) | |
| Addison | Large | West | 1080 | 27-Aug. | 7.96 (± 1.51) | |
| Ferrisburgh | Large | West | 3708 | 6-Aug. | 40.53 (± 5.06) | |
| Calais | Small | East | 365 | 24-Aug. | 20.25 (± 3.39) | |
| Berlin A | Medium | East | 149 | 22-Aug. | 4.27 (± 0.74) | |
| Berlin B | Medium | East | 1131 | 22-Aug. | 30.04 (± 8.50) |
| HA | Farm | Farm size | Micro region | Season sum | Peak date | Peak date mean (± SEM) |
|---|---|---|---|---|---|---|
| 2012 | Alburgh | Large | North | 172 | 25-July | 1.22 (± 0.55) |
| Addison | Large | West | 984 | 24-July | 3.18 (± 0.74) | |
| Berlin A | Medium | East | 224 | 7-Aug. | 3.86 (± 0.95) | |
| 2013 | Alburgh | Large | North | 1562 | 12-Aug. | 12.93 (± 1.60) |
| North Hero | Small | North | 204 | 9-July | 5.35 (± 1.73) | |
| Addison | Large | West | 522 | 29-July | 3.00 (± 0.54) | |
| Calais | Small | East | 93 | 6-Aug. | 5.88 (± 2.13) | |
| Berlin A | Medium | East | 97 | 6-Aug. | 0.80 (± 0.28) | |
| 2014 | Alburgh | Large | North | 2864 | 4-Aug. | 16.93 (± 3.20) |
| North Hero | Small | North | 201 | 18-Aug. | 5.81 (± 2.30) | |
| Addison | Large | West | 1080 | 27-Aug. | 7.96 (± 1.51) | |
| Ferrisburgh | Large | West | 3708 | 6-Aug. | 40.53 (± 5.06) | |
| Calais | Small | East | 365 | 24-Aug. | 20.25 (± 3.39) | |
| Berlin A | Medium | East | 149 | 22-Aug. | 4.27 (± 0.74) | |
| Berlin B | Medium | East | 1131 | 22-Aug. | 30.04 (± 8.50) |
Potato leafhopper (PLH) season sum, peak date, and peak date mean (± SEM) by year and farm
| PLH | Farm | Farm size | Micro region | Season sum | Peak date | Peak date mean (± SEM) |
|---|---|---|---|---|---|---|
| 2012 | Alburgh | Large | North | 730 | 29-June | 6.01 (± 0.60) |
| Addison | Large | West | 588 | 28-June | 6.50 (± 0.97) | |
| Berlin A | Medium | East | 573 | 30-June | 14.70 (± 1.33) | |
| 2013 | Alburgh | Large | North | 349 | 24-June | 3.11 (± 0.65) |
| North Hero | Small | North | 291 | 24-June | 6.75 (± 1.73) | |
| Addison | Large | West | 945 | 18-June | 5.65 (± 0.79) | |
| Calais | Small | East | 69 | 9-July | 4.38 (± 1.44) | |
| Berlin A | Medium | East | 284 | 9-July | 5.67 (± 0.84) | |
| 2014 | Alburgh | Large | North | 113 | 8-July | 0.59 (± 0.14) |
| North Hero | Small | North | 34 | 21-July | 1.12 (± 0.34) | |
| Addison | Large | West | 183 | 11-July | 0.83 (± 0.17) | |
| Ferrisburgh | Large | West | 62 | 9-July | 0.65 (± 0.18) | |
| Calais | Small | East | 3 | 25-July | 0.38 (± 0.26) | |
| Berlin A | Medium | East | 6 | 11-July | 0.17 (± 0.10) | |
| Berlin B | Medium | East | 19 | 25-July | 0.33 (± 0.12) |
| PLH | Farm | Farm size | Micro region | Season sum | Peak date | Peak date mean (± SEM) |
|---|---|---|---|---|---|---|
| 2012 | Alburgh | Large | North | 730 | 29-June | 6.01 (± 0.60) |
| Addison | Large | West | 588 | 28-June | 6.50 (± 0.97) | |
| Berlin A | Medium | East | 573 | 30-June | 14.70 (± 1.33) | |
| 2013 | Alburgh | Large | North | 349 | 24-June | 3.11 (± 0.65) |
| North Hero | Small | North | 291 | 24-June | 6.75 (± 1.73) | |
| Addison | Large | West | 945 | 18-June | 5.65 (± 0.79) | |
| Calais | Small | East | 69 | 9-July | 4.38 (± 1.44) | |
| Berlin A | Medium | East | 284 | 9-July | 5.67 (± 0.84) | |
| 2014 | Alburgh | Large | North | 113 | 8-July | 0.59 (± 0.14) |
| North Hero | Small | North | 34 | 21-July | 1.12 (± 0.34) | |
| Addison | Large | West | 183 | 11-July | 0.83 (± 0.17) | |
| Ferrisburgh | Large | West | 62 | 9-July | 0.65 (± 0.18) | |
| Calais | Small | East | 3 | 25-July | 0.38 (± 0.26) | |
| Berlin A | Medium | East | 6 | 11-July | 0.17 (± 0.10) | |
| Berlin B | Medium | East | 19 | 25-July | 0.33 (± 0.12) |
Potato leafhopper (PLH) season sum, peak date, and peak date mean (± SEM) by year and farm
| PLH | Farm | Farm size | Micro region | Season sum | Peak date | Peak date mean (± SEM) |
|---|---|---|---|---|---|---|
| 2012 | Alburgh | Large | North | 730 | 29-June | 6.01 (± 0.60) |
| Addison | Large | West | 588 | 28-June | 6.50 (± 0.97) | |
| Berlin A | Medium | East | 573 | 30-June | 14.70 (± 1.33) | |
| 2013 | Alburgh | Large | North | 349 | 24-June | 3.11 (± 0.65) |
| North Hero | Small | North | 291 | 24-June | 6.75 (± 1.73) | |
| Addison | Large | West | 945 | 18-June | 5.65 (± 0.79) | |
| Calais | Small | East | 69 | 9-July | 4.38 (± 1.44) | |
| Berlin A | Medium | East | 284 | 9-July | 5.67 (± 0.84) | |
| 2014 | Alburgh | Large | North | 113 | 8-July | 0.59 (± 0.14) |
| North Hero | Small | North | 34 | 21-July | 1.12 (± 0.34) | |
| Addison | Large | West | 183 | 11-July | 0.83 (± 0.17) | |
| Ferrisburgh | Large | West | 62 | 9-July | 0.65 (± 0.18) | |
| Calais | Small | East | 3 | 25-July | 0.38 (± 0.26) | |
| Berlin A | Medium | East | 6 | 11-July | 0.17 (± 0.10) | |
| Berlin B | Medium | East | 19 | 25-July | 0.33 (± 0.12) |
| PLH | Farm | Farm size | Micro region | Season sum | Peak date | Peak date mean (± SEM) |
|---|---|---|---|---|---|---|
| 2012 | Alburgh | Large | North | 730 | 29-June | 6.01 (± 0.60) |
| Addison | Large | West | 588 | 28-June | 6.50 (± 0.97) | |
| Berlin A | Medium | East | 573 | 30-June | 14.70 (± 1.33) | |
| 2013 | Alburgh | Large | North | 349 | 24-June | 3.11 (± 0.65) |
| North Hero | Small | North | 291 | 24-June | 6.75 (± 1.73) | |
| Addison | Large | West | 945 | 18-June | 5.65 (± 0.79) | |
| Calais | Small | East | 69 | 9-July | 4.38 (± 1.44) | |
| Berlin A | Medium | East | 284 | 9-July | 5.67 (± 0.84) | |
| 2014 | Alburgh | Large | North | 113 | 8-July | 0.59 (± 0.14) |
| North Hero | Small | North | 34 | 21-July | 1.12 (± 0.34) | |
| Addison | Large | West | 183 | 11-July | 0.83 (± 0.17) | |
| Ferrisburgh | Large | West | 62 | 9-July | 0.65 (± 0.18) | |
| Calais | Small | East | 3 | 25-July | 0.38 (± 0.26) | |
| Berlin A | Medium | East | 6 | 11-July | 0.17 (± 0.10) | |
| Berlin B | Medium | East | 19 | 25-July | 0.33 (± 0.12) |
Potato Leafhopper
Potato leafhopper populations were highest in 2012 and 2013 across all locations with peak dates in late June and early July (Fig. 14). Two generations of potato leafhopper were observed in 2012 and 2013 on multiple farms. This migratory pest arrived late to the northeast in 2014. A low, later population was therefore observed in 2014 with peak dates throughout July. Eastern sites had very few individuals in the 2014 season. Addison was the location with the highest potato leafhopper pressure, yet it is also the location with the highest natural enemy abundance. Medium-sized Berlin A had similar potato leafhopper abundance to Addison in 2012 (Table 6). Both locations exhibited natural enemy presence. In 2013, Berlin A potato leafhopper pest abundance dropped and natural enemy abundance increased, potentially indicating the impact of natural enemies. Spiders were a prevalent potato leafhopper natural enemy in all locations. Potato leafhopper was typically an early season pest, attacking new growth. However, given the unpredictable migration of this pest, females could arrive before natural enemy populations become established in the spring. Potato leafhopper population numbers were less variable by location or farm size than hop aphids or twospotted spider mites.
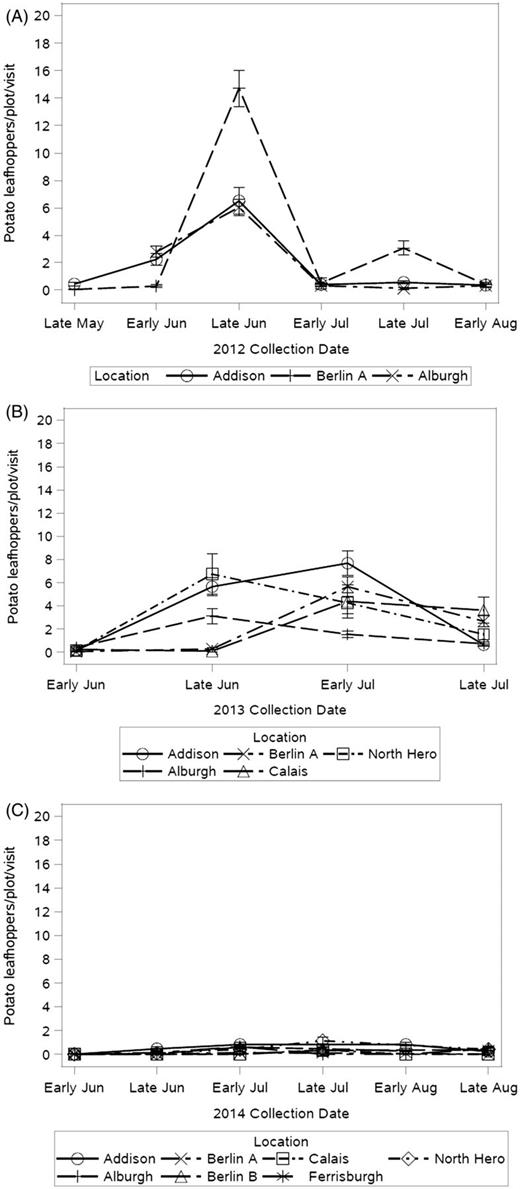
Mean number of potato leafhoppers counted per plot, per yard visit in 2012 (A), 2013 (B), and 2014 (C). Three farms were scouted in 2012, five in 2013, and seven in 2014.
Minor Pests
Eastern comma, Polygonia comma Harris (Lepidoptera: Nymphalidae), adults and larva (Fig. 15) were abundant only in 2012 and have not been reported as a problem since. Japanese beetle, Popillia japonica Newman (Coleoptera: Scarabaeidae) (Fig. 16), was reported as a problem on hops in southern parts of the northeast region. Low numbers of this pest were observed in 2012. A Japanese beetle hot spot was observed at the Berlin A location in late July through early August of 2013. Hop looper was spotted on all scouted hop yards during at least 1 yr of the survey at low abundance (Table 7). The foliar chewing damage that Japanese beetle and eastern comma larvae incur to hop plants is visually shocking. However, because Japanese beetles are foliar feeders and become a problem in late July or early August, their damage is unlikely to reduce hop yield.

Eastern comma larva, a minor hop pest.

Japanese beetle adults and defoliation. This is a minor pest in northern hop yards yet a more serious pest in southern locations.
Minor pests including Eastern comma (Polygonia comma Harris) adult and larvae, hop looper (Hypena humuli Harris) larvae, and Japanese beetle (Popillia japonica Newman) adult season sums are reported
| Year | Farm | Farm size | Micro region | Season sum | ||
|---|---|---|---|---|---|---|
| Eastern comma | Hop looper | Japanese beetle | ||||
| 2012 | Alburgh | Large | North | 55 | 7 | 1 |
| Addison | Large | West | 321 | 7 | 5 | |
| Berlin A | Medium | East | 100 | 3 | 4 | |
| 2013 | Alburgh | Large | North | 5 | 1 | 1 |
| North Hero | Small | North | 0 | 0 | 1 | |
| Addison | Large | West | 0 | 2 | 7 | |
| Calais | Small | East | 0 | 2 | 0 | |
| Berlin A | Medium | East | 1 | 7 | 93 | |
| 2014 | Alburgh | Large | North | 0 | 10 | 1 |
| North Hero | Small | North | 0 | 5 | 0 | |
| Addison | Large | West | 0 | 8 | 0 | |
| Ferrisburgh | Large | West | 0 | 7 | 0 | |
| Calais | Small | East | 0 | 5 | 0 | |
| Berlin A | Medium | East | 0 | 7 | 0 | |
| Berlin B | Medium | East | 0 | 12 | 0 | |
| Year | Farm | Farm size | Micro region | Season sum | ||
|---|---|---|---|---|---|---|
| Eastern comma | Hop looper | Japanese beetle | ||||
| 2012 | Alburgh | Large | North | 55 | 7 | 1 |
| Addison | Large | West | 321 | 7 | 5 | |
| Berlin A | Medium | East | 100 | 3 | 4 | |
| 2013 | Alburgh | Large | North | 5 | 1 | 1 |
| North Hero | Small | North | 0 | 0 | 1 | |
| Addison | Large | West | 0 | 2 | 7 | |
| Calais | Small | East | 0 | 2 | 0 | |
| Berlin A | Medium | East | 1 | 7 | 93 | |
| 2014 | Alburgh | Large | North | 0 | 10 | 1 |
| North Hero | Small | North | 0 | 5 | 0 | |
| Addison | Large | West | 0 | 8 | 0 | |
| Ferrisburgh | Large | West | 0 | 7 | 0 | |
| Calais | Small | East | 0 | 5 | 0 | |
| Berlin A | Medium | East | 0 | 7 | 0 | |
| Berlin B | Medium | East | 0 | 12 | 0 | |
Minor pests including Eastern comma (Polygonia comma Harris) adult and larvae, hop looper (Hypena humuli Harris) larvae, and Japanese beetle (Popillia japonica Newman) adult season sums are reported
| Year | Farm | Farm size | Micro region | Season sum | ||
|---|---|---|---|---|---|---|
| Eastern comma | Hop looper | Japanese beetle | ||||
| 2012 | Alburgh | Large | North | 55 | 7 | 1 |
| Addison | Large | West | 321 | 7 | 5 | |
| Berlin A | Medium | East | 100 | 3 | 4 | |
| 2013 | Alburgh | Large | North | 5 | 1 | 1 |
| North Hero | Small | North | 0 | 0 | 1 | |
| Addison | Large | West | 0 | 2 | 7 | |
| Calais | Small | East | 0 | 2 | 0 | |
| Berlin A | Medium | East | 1 | 7 | 93 | |
| 2014 | Alburgh | Large | North | 0 | 10 | 1 |
| North Hero | Small | North | 0 | 5 | 0 | |
| Addison | Large | West | 0 | 8 | 0 | |
| Ferrisburgh | Large | West | 0 | 7 | 0 | |
| Calais | Small | East | 0 | 5 | 0 | |
| Berlin A | Medium | East | 0 | 7 | 0 | |
| Berlin B | Medium | East | 0 | 12 | 0 | |
| Year | Farm | Farm size | Micro region | Season sum | ||
|---|---|---|---|---|---|---|
| Eastern comma | Hop looper | Japanese beetle | ||||
| 2012 | Alburgh | Large | North | 55 | 7 | 1 |
| Addison | Large | West | 321 | 7 | 5 | |
| Berlin A | Medium | East | 100 | 3 | 4 | |
| 2013 | Alburgh | Large | North | 5 | 1 | 1 |
| North Hero | Small | North | 0 | 0 | 1 | |
| Addison | Large | West | 0 | 2 | 7 | |
| Calais | Small | East | 0 | 2 | 0 | |
| Berlin A | Medium | East | 1 | 7 | 93 | |
| 2014 | Alburgh | Large | North | 0 | 10 | 1 |
| North Hero | Small | North | 0 | 5 | 0 | |
| Addison | Large | West | 0 | 8 | 0 | |
| Ferrisburgh | Large | West | 0 | 7 | 0 | |
| Calais | Small | East | 0 | 5 | 0 | |
| Berlin A | Medium | East | 0 | 7 | 0 | |
| Berlin B | Medium | East | 0 | 12 | 0 | |
Generalist Predators
Because the arthropod community was disturbed as plants were approached, parasitoid wasps were seldom observed and the magnitude of generalist predators may not be represented. Across farms, lady beetles and spiders were the most abundant generalist predators. Lacewings were the next most abundant natural enemy group followed by syrphid flies and finally minute pirate bugs. As expected, large farms had the highest natural enemy abundance. However, over the course of 3 yr the smaller North Hero and Calais farms exhibited an increase in the number of total generalist predators. Hop yards that were sampled all 3 yr (Alburgh, Addison, and Berlin A) had the highest generalist natural enemy abundance in 2012 (Table 8). These generalist natural enemy populations coincide with high pest abundance in 2012. Hop aphids and potato leafhoppers totaled 2,201 in 2012, 1,680 in 2013, and 356 in 2014 in the yards scouted for 3 yr.
Season sums by farm and year for generalist predators including lady beetle (Coccinellidae) adults and larva, spiders, lacewings (Chrysopidae and Hemerobiidae) adult and larva, syrphid fly (Syrphidae) adults, and minute pirate bug (Orius spp.) adults
| Year | Farm | Farm size | Micro region | Season sum | ||||
|---|---|---|---|---|---|---|---|---|
| Lady beetles | Spiders | Lacewings | Syrphid flies | Minute pirate bugs | ||||
| 2012 | Alburgh | Large | North | 56 | 39 | 70 | 0 | 0 |
| Addison | Large | West | 109 | 32 | 31 | 2 | 0 | |
| Berlin A | Medium | East | 33 | 9 | 9 | 0 | 0 | |
| 2013 | Alburgh | Large | North | 4 | 6 | 3 | 7 | 0 |
| North Hero | Small | North | 3 | 17 | 5 | 0 | 0 | |
| Addison | Large | West | 45 | 43 | 17 | 11 | 1 | |
| Calais | Small | East | 0 | 12 | 0 | 0 | 0 | |
| Berlin A | Medium | East | 5 | 29 | 8 | 1 | 0 | |
| 2014 | Alburgh | Large | North | 8 | 56 | 1 | 0 | 1 |
| North Hero | Small | North | 15 | 23 | 0 | 0 | 0 | |
| Addison | Large | West | 34 | 26 | 3 | 0 | 0 | |
| Ferrisburgh | Large | West | 41 | 28 | 7 | 3 | 0 | |
| Calais | Small | East | 18 | 9 | 0 | 0 | 0 | |
| Berlin A | Medium | East | 6 | 26 | 1 | 0 | 0 | |
| Berlin B | Medium | East | 17 | 11 | 1 | 0 | 0 | |
| Year | Farm | Farm size | Micro region | Season sum | ||||
|---|---|---|---|---|---|---|---|---|
| Lady beetles | Spiders | Lacewings | Syrphid flies | Minute pirate bugs | ||||
| 2012 | Alburgh | Large | North | 56 | 39 | 70 | 0 | 0 |
| Addison | Large | West | 109 | 32 | 31 | 2 | 0 | |
| Berlin A | Medium | East | 33 | 9 | 9 | 0 | 0 | |
| 2013 | Alburgh | Large | North | 4 | 6 | 3 | 7 | 0 |
| North Hero | Small | North | 3 | 17 | 5 | 0 | 0 | |
| Addison | Large | West | 45 | 43 | 17 | 11 | 1 | |
| Calais | Small | East | 0 | 12 | 0 | 0 | 0 | |
| Berlin A | Medium | East | 5 | 29 | 8 | 1 | 0 | |
| 2014 | Alburgh | Large | North | 8 | 56 | 1 | 0 | 1 |
| North Hero | Small | North | 15 | 23 | 0 | 0 | 0 | |
| Addison | Large | West | 34 | 26 | 3 | 0 | 0 | |
| Ferrisburgh | Large | West | 41 | 28 | 7 | 3 | 0 | |
| Calais | Small | East | 18 | 9 | 0 | 0 | 0 | |
| Berlin A | Medium | East | 6 | 26 | 1 | 0 | 0 | |
| Berlin B | Medium | East | 17 | 11 | 1 | 0 | 0 | |
Season sums by farm and year for generalist predators including lady beetle (Coccinellidae) adults and larva, spiders, lacewings (Chrysopidae and Hemerobiidae) adult and larva, syrphid fly (Syrphidae) adults, and minute pirate bug (Orius spp.) adults
| Year | Farm | Farm size | Micro region | Season sum | ||||
|---|---|---|---|---|---|---|---|---|
| Lady beetles | Spiders | Lacewings | Syrphid flies | Minute pirate bugs | ||||
| 2012 | Alburgh | Large | North | 56 | 39 | 70 | 0 | 0 |
| Addison | Large | West | 109 | 32 | 31 | 2 | 0 | |
| Berlin A | Medium | East | 33 | 9 | 9 | 0 | 0 | |
| 2013 | Alburgh | Large | North | 4 | 6 | 3 | 7 | 0 |
| North Hero | Small | North | 3 | 17 | 5 | 0 | 0 | |
| Addison | Large | West | 45 | 43 | 17 | 11 | 1 | |
| Calais | Small | East | 0 | 12 | 0 | 0 | 0 | |
| Berlin A | Medium | East | 5 | 29 | 8 | 1 | 0 | |
| 2014 | Alburgh | Large | North | 8 | 56 | 1 | 0 | 1 |
| North Hero | Small | North | 15 | 23 | 0 | 0 | 0 | |
| Addison | Large | West | 34 | 26 | 3 | 0 | 0 | |
| Ferrisburgh | Large | West | 41 | 28 | 7 | 3 | 0 | |
| Calais | Small | East | 18 | 9 | 0 | 0 | 0 | |
| Berlin A | Medium | East | 6 | 26 | 1 | 0 | 0 | |
| Berlin B | Medium | East | 17 | 11 | 1 | 0 | 0 | |
| Year | Farm | Farm size | Micro region | Season sum | ||||
|---|---|---|---|---|---|---|---|---|
| Lady beetles | Spiders | Lacewings | Syrphid flies | Minute pirate bugs | ||||
| 2012 | Alburgh | Large | North | 56 | 39 | 70 | 0 | 0 |
| Addison | Large | West | 109 | 32 | 31 | 2 | 0 | |
| Berlin A | Medium | East | 33 | 9 | 9 | 0 | 0 | |
| 2013 | Alburgh | Large | North | 4 | 6 | 3 | 7 | 0 |
| North Hero | Small | North | 3 | 17 | 5 | 0 | 0 | |
| Addison | Large | West | 45 | 43 | 17 | 11 | 1 | |
| Calais | Small | East | 0 | 12 | 0 | 0 | 0 | |
| Berlin A | Medium | East | 5 | 29 | 8 | 1 | 0 | |
| 2014 | Alburgh | Large | North | 8 | 56 | 1 | 0 | 1 |
| North Hero | Small | North | 15 | 23 | 0 | 0 | 0 | |
| Addison | Large | West | 34 | 26 | 3 | 0 | 0 | |
| Ferrisburgh | Large | West | 41 | 28 | 7 | 3 | 0 | |
| Calais | Small | East | 18 | 9 | 0 | 0 | 0 | |
| Berlin A | Medium | East | 6 | 26 | 1 | 0 | 0 | |
| Berlin B | Medium | East | 17 | 11 | 1 | 0 | 0 | |
Specific Farm Factors
Season totals and graphical phenology of pest and peak date means by location provide insight into northeast hop yard arthropod patterns. However, the effects of cultivar genetic variation, specific management practices, surrounding habitat, previous crop, and hop yard age on the arthropod community were not accounted for in this survey. There are specific management practices that we believe to have influenced certain pest populations. Alburgh exhibited almost the same total number of spider mites in 2012 and 2014. Pyganic, a pyrethrin broad-spectrum insecticide, was sprayed multiple times at this location for management of potato leafhopper in 2012. This likely caused the late season secondary outbreak of twospotted spider mites at this site. Hop quality and yield were negatively affected by this outbreak in 2012. Insecticides have not been applied in the Alburgh hop yard since. The same total number of spider mites in 2014 did not have an impact on hop quality or yield in the unsprayed yard.
Moving Forward
The patterns observed in this survey were the first published documentation of northeastern pests and natural enemies since the 1940s (Magie 1944). We consider the hop arthropod pests of concern in the northeast to be twospotted spider mite, hop aphid, and potato leafhopper. We observed similar pest phenology to other hop-growing regions for twospotted spider mites and hop aphids.
Arthropod communities are known to change based on climate, landscape, and management practices (Schweiger et al. 2005). Microclimate and hop yard size factors reported here observationally influenced pest abundance. As expected, where high pest populations were observed, natural enemies were more abundant. Habitat diversity and surrounding landscape, not reported, are well-known variables that increase natural enemy presence (Landis et al. 2000, Rusch et al. 2010).
Hop cones grow throughout the canopy of plants. Although logistically challenging, we suggest that mid and late season pest monitoring could be improved by sampling above eye level in the hop canopy. Unbaited sticky traps can be pinned to trellis wire to monitor flying pests and natural enemies. Growers should visually inspect the underside of leaves on all cultivars as weekly scouting. Parasitoid wasps, not reported, are important and numerous members of hop aphid and potato leafhopper natural enemy assemblages (unpublished data).
Weather was an indicator of pest abundance in northeastern hops. In a dry, hot year, we expect to see high twospotted spider mite populations, yet with cooler temperatures and early or continued precipitation, we expect to see high aphid populations throughout August. Potato leafhopper population prediction will rely on southern reports of presence on other crops, such as alfalfa, that are routinely monitored.
Economic threshold levels have not been developed for these pests in the northeast. When making pest control decisions, eastern Extension professionals reference Pacific Northwestern and European economic threshold ranges for twospotted spider mites (10–100 mites per leaf; Wright and Cone 1999, Weihrauch 2004) and hop aphid (5–70 aphids per leaf; Lorenzana et al. 2009). The literature suggests that thresholds for these pests vary considerably both locally and regionally (Weihrauch 2004, Lorenzana et al. 2009). The research hop yard in Alburgh, VT, has not exceeded a yard mean of 60 twospotted spider mites per leaf or a yard mean of 10 hop aphids per leaf during peak pest abundance. These levels have, however, been reached in pockets of the Alburgh hop yard.
This survey provided evidence that natural enemies and insecticide applications impact pest populations. Natural enemies were present to varying degrees in northeast hop yards, and once established, they appeared to maintain pest populations. This was particularly clear in the Addison hop yard. Insecticides have the opposite effect, as they kill natural enemies and increase the risk of a twospotted spider mite outbreak. This was observed in Alburgh (2012) and Ferrisburgh (2014) yards.
The work presented was the first step toward developing appropriate IPM tactics for modern day northeastern hop production. As evidenced, it is always important to consider site-specific factors that may influence arthropod populations. Northeastern hop arthropod and disease IPM requires further research. Based on this survey, economic thresholds for the three major arthropod pests should be developed. Although some research on natural enemy populations in hop yards has been conducted in the Pacific Northwest, development of conservation and augmentative biological control protocols would reduce pesticide use and provide management options for organic growers. Finally, research on hop downy mildew and weed management will be required in order to produce high quality northeastern hops.
Acknowledgments
We continue to appreciate the growers who opened their farms to our scouting efforts and have participated in our grower surveys. We thank the permanent and seasonal employees of the UVM Extension Northwest Crops and Soils Team for their time spent collecting data. This research would not have been possible without financial support from the Northeast Sustainable Agriculture Research and Education (SARE) program (GNE12-033) and the United States Department of Agriculture-National Institute of Food and Agriculture (USDA-NIFA) Northeastern IPM Center (027269).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}