





Abstract
Quorum sensing (QS) is the regulation of gene expression in response to the concentration of small signal molecules, and its inactivation has been suggested to have great potential to attenuate microbial virulence. It is assumed that unlike antimicrobials, inhibition of QS should cause less Darwinian selection pressure for bacterial resistance. Using the opportunistic pathogen Pseudomonas aeruginosa, we demonstrate here that bacterial resistance arises rapidly to the best-characterized compound that inhibits QS (brominated furanone C-30) due to mutations that increase the efflux of C-30. Critically, the C-30-resistant mutant mexR was more pathogenic to Caenorhabditis elegans in the presence of C-30, and the same mutation arises in bacteria responsible for chronic cystic fibrosis infections. Therefore, bacteria may evolve resistance to many new pharmaceuticals thought impervious to resistance.
Introduction
Bacteria have been identified that are resistant to all known antibiotics (Defoirdt et al., 2010), and infectious diseases remain the leading cause of death (Rasko and Sperandio, 2010); hence, it is important to develop new antimicrobials. Indeed, it has been decreed that we live in a post-antibiotic era and heralded that anti-quorum sensing (QS)/antivirulence methods hold great promise for treating bacterial infections (Rasko and Sperandio, 2010). One of the most attractive features of this approach is that by interrupting cell signaling, these approaches do not impose harsh or direct selective pressure like antibiotics (Bjarnsholt et al., 2010), so there is less evolutionary pressure to develop resistance to antivirulence compounds (Bjarnsholt et al., 2010; Rasko and Sperandio, 2010).
The best-characterized of the antivirulence compounds are the brominated furanones, which are secreted by the seaweed Delisea pulchra to prevent biofilms from inhibiting its photosynthesis; to date, no bacteria have been identified that are resistant to them. The natural compound (5Z)-4-bromo-5-(bromomethylene)-3-butyl-2(5H)-furanone from D. pulchra inhibits both acyl-homoserine lactone-based and autoinducer 2-based QS (Ren et al., 2001), and the synthetic furanone C-30 (Figure 1a inset) has been shown to decrease acyl-homoserine lactone-based signaling as well as decrease the virulence of P. aeruginosa in a mouse pulmonary infection model (Hentzer et al., 2003). These brominated furanones interrupt QS by interacting with transcriptional regulators that propagate the QS response (Defoirdt et al., 2007), and they do not affect bacterial growth in rich medium (Gram et al., 1996; Ren et al., 2001; Hentzer et al., 2003). However, growth of pathogens in the host during infections is more likely to involve non-robust carbon sources and may involve compounds whose utilization depends on QS (Defoirdt et al., 2010). Under these conditions, there will be selection pressure to evolve resistance against the compounds that block QS, and it has been theorized, but not shown, that resistance may arise (Defoirdt et al., 2010). Note that previously mutations to the QS response regulator LuxR were generated in Escherichia coli, which altered both the binding of the natural ligand as well as that of quorum quenching (QQ) compounds, but resistance was not investigated and was predicted not to occur (Koch et al., 2005).
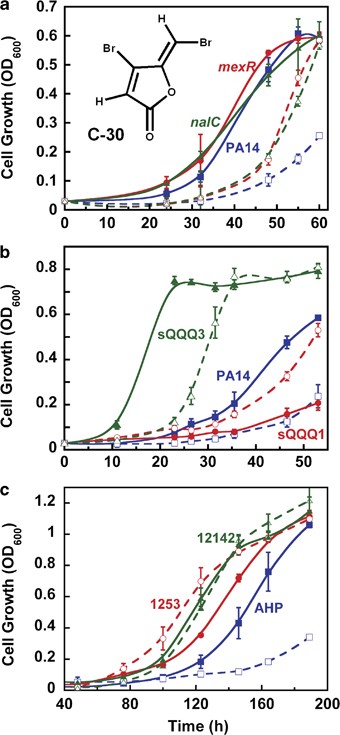
The mexR and nalC mutations render P. aeruginosa less sensitive to QQ compound C-30 during growth in adenosine minimal medium. (a) Growth (at 37 °C) of wild-type P. aeruginosa PA14 (blue squares), transposon mutant mexR (red circles) and transposon mutant nalC (green triangles). (b) Growth of spontaneous quorum quenching quandary mutants (sQQQ) 1 (red circles) and sQQQ3 (green triangles) vs wild-type PA14 (blue squares). (c) Growth of CF clinical isolate 1253 (red circles) and CF Liverpool epidemic strain 12142 (green triangles) with enhanced mexA expression vs control strain AHP (blue squares). Open symbols indicate the presence of 50 μm C-30, whereas closed symbols indicate the absence of C-30. Data represent the mean±s.d.; n=3.
To investigate whether QQ-resistant bacterial mutants may arise in the presence of antivirulence compounds, we utilized the best studied bacterium for QS, P. aeruginosa, which is an opportunistic pathogen that is responsible for many infections, including those of ventilator-associated pneumonia, urinary and peritoneal dialysis, catheter infections, bacterial keratitis, otitis externa, burns, wound infections and those of the lung (Macé et al., 2008). Wild-type P. aeruginosa PA14 was used instead of PAO1 because PA14 is more virulent than PAO1 in diverse infection models (Harrison et al., 2010) and because of the availability of the complete mutant library (Liberati et al., 2006). Our strategy was to utilize a minimal medium (so it resembles more closely clinical situations) using a growth compound whose assimilation requires QS; therefore, QS and growth were inhibited by the antivirulence compound C-30, which has become the gold standard for antivirulence compounds. Growth on adenosine by P. aeruginosa depends on the degradative enzyme, nucleoside hydrolase, which is positively controlled by LasR (Heurlier et al., 2005). LasR is the transcriptional regulator that mediates acyl-homoserine lactone-based QS in this strain via N-3-oxododecanoyl homoserine lactone (3OC12-HSL). In addition, furanone derivatives including C-30 have been predicted to bind LasR at the 3OC12-HSL binding site to inactivate LasR (Kim et al., 2008; Yang et al., 2009). Hence, growth on adenosine in minimal medium depends on a functional LasR QS pathway (Heurlier et al., 2005), and furanone probably inhibits this growth via LasR. Therefore, we reasoned that mutations that confer resistance to furanone C-30 will disrupt the ability of furanone to inhibit LasR-mediated QS in this strain, and these mutants may be identified easily based on our selection method (faster growth).
It was also advantageous to utilize growth on adenosine as our model system because adenosine is released into the human intestinal tissues and lumen during surgical injury, ischemia and inflammation (Patel et al., 2007; Zaborin et al., 2009), and adenosine induces the P. aeruginosa virulence factor PA-I lectin/adhesin, which leads to lethal gut-derived sepsis (Patel et al., 2007). Note the extracellular concentration of adenosine, which is generated by breakdown of secreted ATP (Crane et al., 2002), can reach 5 mm (Kimura et al., 2005) and increase 109-fold in human intestinal epithelial cells after exposure to hypoxia (Patel et al., 2007). Because adenosine is secreted by the host as a cytoprotective compound that tightens epithelial cell junctions, P. aeruginosa probably metabolizes adenosine to increase its ability to act as a pathogen (Patel et al., 2007). Furthermore, although P. aeruginosa is better known as a respiratory pathogen rather than an intestinal pathogen, this strain is one of the most common opportunistic pathogens in the normal gastrointestinal tract (Alverdy et al., 2000), and in critically-ill and immunocompromised patients where stress levels are high, Pseudomonas sp. counts have been shown to increase by as much as 100-fold (Shimizu et al., 2006) leading to the expression of virulence determinants (Alverdy et al., 2000) that are controlled by QS (Hegde et al., 2009) (just the presence of P. aeruginosa in the gastrointestinal tract of critically-ill surgical patients has been associated with nearly 70% mortality (Alverdy et al., 2000)). Hence, growth on this carbon source is physiologically relevant and P. aeruginosa infections in the gastrointestinal tract are pertinent.
Using the novel adenosine screen, we found that P. aeruginosa cells may evolve resistance to the QQ compound C-30. The mechanism for this resistance to QQ was determined to be due to mexR and nalC mutations, which encode repressors of the mexAB-oprM multi-drug resistance operon; hence, the mutants have enhanced efflux of C-30. In addition, we demonstrate that the mutants are resistant to C-30 in an animal model. We also show that P. aeruginosa isolates with mutations in mexR and nalC that arise in chronic cystic fibrosis (CF) patients after antibiotic therapy have evolved resistance to the QQ compound C-30, illustrating that these mutants are physiologically relevant.
Materials and methods
Strains and growth conditions
All experiments were conducted at 37 °C and included at least three independent cultures. The bacterial strains used in this study are listed in Table 1. Wild-type P. aeruginosa PA14 from Dr Frederick Ausubel (Liberati et al., 2006) was utilized. The brominated furanone C-30 was synthesized (Jones et al., 2005), confirmed by 1H-NMR and GC-MS (Jones et al., 2005) and dissolved in methanol; 50 μm was used for all experiments.
Bacterial strains used in this study
| Strain | Genotype/relevant characteristics | Source |
|---|---|---|
| PA14 | Pseudomonas aeruginosa wild-type | Liberati et al. (2006) |
| mexR | PA14 mexR Ω miniTn5 luxAB-Tet, TcR | This study |
| nalC | PA14 nalC Ω miniTn5 luxAB-Tet, TcR | This study |
| sQQQ1 | Spontaneous C-30 resistant PA14 mutant | This study |
| sQQQ3 | Spontaneous C-30 resistant PA14 mutant | This study |
| phzM | PA14 phzM Ω Mar2xT7, GmR | Liberati et al. (2006) |
| rhlR | PA14 rhlR Ω Mar2xT7, GmR | Liberati et al. (2006) |
| lasB | PA14 lasB Ω Mar2xT7, GmR | Liberati et al. (2006) |
| AHP | mexR (V126E), nalC (G71E, A78T, S209R) | Tomás et al. (2010) |
| 12142a | mexR (R83C), nalC (G71E), nalD (D187H) | Tomás et al. (2010) |
| 1253a | mexR (V126E), nalC (G71E, A186T), nalD (D187H) | Tomás et al. (2010) |
| 14169a | mexR (R83C), nalC (G71E), nalD (D187H) | Tomás et al. (2010) |
| 1681a | mexR (R83C), nalC (G71E), nalD (D187H) | Tomás et al. (2010) |
| 3149a | nalC (G71E), nalD (D187H, L201P) | Tomás et al. (2010) |
| 1248a | mexR (R83C), nalC (G71E), qnalD (D187H, L201P) | Tomás et al. (2010) |
| 12102a | mexR (V126E), nalC (G71E), nalD (D187H) | Tomás et al. (2010) |
| 1665a | mexR (R83C), nalC (G71E), nalD (D187H, L201P) | Tomás et al. (2010) |
| 1712a | mexR (R83C), nalC (G71E, S209R), nalD (D187H, L201P) | Tomás et al. (2010) |
| OP50 | Escherichia coli B strain (uracil auxotroph) | Brenner (1974) |
| Strain | Genotype/relevant characteristics | Source |
|---|---|---|
| PA14 | Pseudomonas aeruginosa wild-type | Liberati et al. (2006) |
| mexR | PA14 mexR Ω miniTn5 luxAB-Tet, TcR | This study |
| nalC | PA14 nalC Ω miniTn5 luxAB-Tet, TcR | This study |
| sQQQ1 | Spontaneous C-30 resistant PA14 mutant | This study |
| sQQQ3 | Spontaneous C-30 resistant PA14 mutant | This study |
| phzM | PA14 phzM Ω Mar2xT7, GmR | Liberati et al. (2006) |
| rhlR | PA14 rhlR Ω Mar2xT7, GmR | Liberati et al. (2006) |
| lasB | PA14 lasB Ω Mar2xT7, GmR | Liberati et al. (2006) |
| AHP | mexR (V126E), nalC (G71E, A78T, S209R) | Tomás et al. (2010) |
| 12142a | mexR (R83C), nalC (G71E), nalD (D187H) | Tomás et al. (2010) |
| 1253a | mexR (V126E), nalC (G71E, A186T), nalD (D187H) | Tomás et al. (2010) |
| 14169a | mexR (R83C), nalC (G71E), nalD (D187H) | Tomás et al. (2010) |
| 1681a | mexR (R83C), nalC (G71E), nalD (D187H) | Tomás et al. (2010) |
| 3149a | nalC (G71E), nalD (D187H, L201P) | Tomás et al. (2010) |
| 1248a | mexR (R83C), nalC (G71E), qnalD (D187H, L201P) | Tomás et al. (2010) |
| 12102a | mexR (V126E), nalC (G71E), nalD (D187H) | Tomás et al. (2010) |
| 1665a | mexR (R83C), nalC (G71E), nalD (D187H, L201P) | Tomás et al. (2010) |
| 1712a | mexR (R83C), nalC (G71E, S209R), nalD (D187H, L201P) | Tomás et al. (2010) |
| OP50 | Escherichia coli B strain (uracil auxotroph) | Brenner (1974) |
Amino acid replacements encoded by the mutations in the CF isolates and their control are shown in parenthesis (note there may be other mutations in these strains).
TcR and GmR indicate tetracycline and gentamicin resistance, respectively.
Indicates clinical CF isolate.
Bacterial strains used in this study
| Strain | Genotype/relevant characteristics | Source |
|---|---|---|
| PA14 | Pseudomonas aeruginosa wild-type | Liberati et al. (2006) |
| mexR | PA14 mexR Ω miniTn5 luxAB-Tet, TcR | This study |
| nalC | PA14 nalC Ω miniTn5 luxAB-Tet, TcR | This study |
| sQQQ1 | Spontaneous C-30 resistant PA14 mutant | This study |
| sQQQ3 | Spontaneous C-30 resistant PA14 mutant | This study |
| phzM | PA14 phzM Ω Mar2xT7, GmR | Liberati et al. (2006) |
| rhlR | PA14 rhlR Ω Mar2xT7, GmR | Liberati et al. (2006) |
| lasB | PA14 lasB Ω Mar2xT7, GmR | Liberati et al. (2006) |
| AHP | mexR (V126E), nalC (G71E, A78T, S209R) | Tomás et al. (2010) |
| 12142a | mexR (R83C), nalC (G71E), nalD (D187H) | Tomás et al. (2010) |
| 1253a | mexR (V126E), nalC (G71E, A186T), nalD (D187H) | Tomás et al. (2010) |
| 14169a | mexR (R83C), nalC (G71E), nalD (D187H) | Tomás et al. (2010) |
| 1681a | mexR (R83C), nalC (G71E), nalD (D187H) | Tomás et al. (2010) |
| 3149a | nalC (G71E), nalD (D187H, L201P) | Tomás et al. (2010) |
| 1248a | mexR (R83C), nalC (G71E), qnalD (D187H, L201P) | Tomás et al. (2010) |
| 12102a | mexR (V126E), nalC (G71E), nalD (D187H) | Tomás et al. (2010) |
| 1665a | mexR (R83C), nalC (G71E), nalD (D187H, L201P) | Tomás et al. (2010) |
| 1712a | mexR (R83C), nalC (G71E, S209R), nalD (D187H, L201P) | Tomás et al. (2010) |
| OP50 | Escherichia coli B strain (uracil auxotroph) | Brenner (1974) |
| Strain | Genotype/relevant characteristics | Source |
|---|---|---|
| PA14 | Pseudomonas aeruginosa wild-type | Liberati et al. (2006) |
| mexR | PA14 mexR Ω miniTn5 luxAB-Tet, TcR | This study |
| nalC | PA14 nalC Ω miniTn5 luxAB-Tet, TcR | This study |
| sQQQ1 | Spontaneous C-30 resistant PA14 mutant | This study |
| sQQQ3 | Spontaneous C-30 resistant PA14 mutant | This study |
| phzM | PA14 phzM Ω Mar2xT7, GmR | Liberati et al. (2006) |
| rhlR | PA14 rhlR Ω Mar2xT7, GmR | Liberati et al. (2006) |
| lasB | PA14 lasB Ω Mar2xT7, GmR | Liberati et al. (2006) |
| AHP | mexR (V126E), nalC (G71E, A78T, S209R) | Tomás et al. (2010) |
| 12142a | mexR (R83C), nalC (G71E), nalD (D187H) | Tomás et al. (2010) |
| 1253a | mexR (V126E), nalC (G71E, A186T), nalD (D187H) | Tomás et al. (2010) |
| 14169a | mexR (R83C), nalC (G71E), nalD (D187H) | Tomás et al. (2010) |
| 1681a | mexR (R83C), nalC (G71E), nalD (D187H) | Tomás et al. (2010) |
| 3149a | nalC (G71E), nalD (D187H, L201P) | Tomás et al. (2010) |
| 1248a | mexR (R83C), nalC (G71E), qnalD (D187H, L201P) | Tomás et al. (2010) |
| 12102a | mexR (V126E), nalC (G71E), nalD (D187H) | Tomás et al. (2010) |
| 1665a | mexR (R83C), nalC (G71E), nalD (D187H, L201P) | Tomás et al. (2010) |
| 1712a | mexR (R83C), nalC (G71E, S209R), nalD (D187H, L201P) | Tomás et al. (2010) |
| OP50 | Escherichia coli B strain (uracil auxotroph) | Brenner (1974) |
Amino acid replacements encoded by the mutations in the CF isolates and their control are shown in parenthesis (note there may be other mutations in these strains).
TcR and GmR indicate tetracycline and gentamicin resistance, respectively.
Indicates clinical CF isolate.
For growth with C-30, overnight cultures in Luria-Bertani (LB) medium were re-inoculated into 20 ml of oxygen-sensing minimal medium containing 0.1% (w/v) adenosine (Heurlier et al., 2005) with or without C-30, and the mixture was incubated with shaking (250 r.p.m.); all cultures had the same initial turbidity at 600 nm. Cell turbidity was measured at 600 nm with a spectrophotometer. The MIC50 value for chloramphenicol was determined with an initial inoculum of 5 × 105 cells ml−1 using the broth dilution method (Wiegand et al., 2008) with the modifications of using shaking (250 r.p.m.) and LB medium.
Quorum-sensing-related assays
Five quorum-sensing-related assays were performed to investigate the impact of C-30 with P. aeruginosa. Pyoverdine production was assayed using overnight cell cultures (100 μl) that were re-inoculated into minimal medium with succinate (Ren et al., 2005) with or without C-30 and incubated for 7 h, then the pyoverdine in the supernatant (20-times diluted) was measured spectrophotometrically at 370 nm (Ren et al., 2005). Pyocyanin production was assayed spectrophotometrically after incubation in LB medium for 9 h and after extracting with chloroform and 0.2 N HCl (Lee et al., 2009) using phzM as a negative control (Ueda et al., 2009). Elastase was assayed using elastin-Congo red as a substrate and lasB as a negative control (Ueda et al., 2009). Swarming was assayed using BM2-swarming medium after incubation for 24 h (Lee et al., 2009), and rhlR as a negative control (Ueda et al., 2009). Biofilm formation was assayed in polystyrene 96-well plates with minimal medium with succinate or glucose by staining with crystal violet (Ueda and Wood, 2009).
Isolation of mutants less sensitive to C-30
A P. aeruginosa PA14 transposon mutant library was generated (Ueda and Wood, 2009) using conjugation with E. coli S17-1 (λpir)/pUT-miniTn5 luxAB-TcR, which delivered the Tn5-luxAB transposon. After conjugation, cells (100 μl) were added to 10 ml of oxygen-sensing minimal medium containing 0.1% (w/v) adenosine (Heurlier et al., 2005) supplemented with C-30, 75 μg ml−1 of tetracycline (to select P. aeruginosa transposon mutants) and 20 μg ml−1 of nalidixic acid (to kill the donor E. coli cells because P. aeruginosa is naturally resistant to this antibiotic). The mixture was incubated with shaking for 2 days, then the cell culture (100 μl) was re-inoculated into the same medium and incubation continued for four passages. The most abundant transposon mutants were selected by streaking an aliquot of the cell culture on LB agar plates containing 75 μg ml−1 of tetracycline and 20 μg ml−1 of nalidixic acid. Single colonies were picked, and their resistance to C-30 on growth was confirmed. To identify the position of the insertion of the transposon, genomic DNA from the transposon mutants was sequenced using two-step PCR (Ueda and Wood, 2009). The mexR mutant with the insertion position at bp 387 (Ala129) from the start of the coding region and the nalC mutant with the insertion at bp 156 (Tyr52) were used for further experiments. Spontaneous mutants that were less sensitive to C-30 were obtained using the same sequential dilution method (without tetracycline).
C. elegans fast-killing assay
To investigate the virulence effect of PA14 strains, the C. elegans fast-killing assay was performed (Mahajan-Miklos et al., 1999). An overnight LB culture was used to spread each of the bacterial cultures (roughly 5 × 107 cells in 5–10 μl) on 3.5-cm diameter peptone-glucose-sorbitol (PGS) agar plates and incubated at 37 °C for 24 h. C-30 was included in both the overnight culture and in the agar plates. After 6–8 h at room temperature, bacterial lawns were heat killed by incubating at 65 °C for 30 min. After cooling to room temperature, each plate was seeded with 20 early to mid-L4 stage hermaphrodite worms (wild-type Bristol N2 strain, Caenorhabditis Genetics Center), and three to four replicates were used for each independent culture. Plates were incubated at 25 °C and scored for live worms. E. coli OP50 was used as a negative control.
Efflux assay
Efflux of C-30 was assayed by measuring the concentration of C-30 exported from cells in minimal succinate medium using high-performance liquid chromatography (4 × 50 mm Inertisil ODS column, Shimadzu SPD-10AVP instrument (Kyoto, Japan) with a UV–Vis detector at 285 nm, and 40% acetonitrile as the mobile phase). Overnight cultures were resuspended in minimal succinate medium to a turbidity of 1 at 600 nm and were allowed to import C-30 for 30 min; the uptake activity of C-30 was determined by measuring the C-30 concentration in the supernatant. The cell pellets were resuspended in minimal succinate medium to a turbidity of 1 at 600 nm and the amount of C-30 effluxed to the supernatant was detected both at 0 h and after 15 h. M9 buffer instead of the minimal succinate medium was used as a negative control.
Whole-transcriptome analysis
The P. aeruginosa genome array (Affymetrix, Santa Clara, CA, USA; P/N 510596) was used to investigate differential gene expression in planktonic cells between PA14 and the mexR mutant in the presence of C-30 (Ueda and Wood, 2009). When the culture grew to the mid-exponential phase (turbidity of 0.25), C-30 was added for 2 h. Total RNA was isolated as described previously (Ren et al., 2004) using a bead beater (Biospec, Bartlesville, OK, USA) and RNAlater buffer (Applied Biosystems, Foster City, CA, USA) to stabilize the RNA. cDNA synthesis, fragmentation, hybridizations and data analysis were as described previously (Lee et al., 2009). To ensure the reliability of the induced/repressed gene list, genes were identified as differentially expressed if the P-value was <0.05 and if the expression ratio was >2 (as the s.d. for the expression ratio for all the genes was 1.7 (Ueda and Wood, 2009)). The whole-transcriptome data were deposited in the NCBI Gene Expression Omnibus (GSE24262).
Quantitative real-time reverse transcription PCR
Quantitative real-time reverse transcription PCR was performed using the StepOnePlus Real-Time PCR System (Applied Biosystems). Expression of nuh gene was determined using the total RNA isolated from two independent cultures of PA14 and the mexR mutant. When the culture reached the mid-exponential phase (turbidity of 0.25), C-30 was added for 30 min. Total RNA was isolated in the same manner as described above for the whole-transcriptome analysis. The primers for quantitative real-time reverse transcription PCR are 5′-GGACTACCTGATCCGTACC-3′ and 5′-GATCTTGTGGGTCACGTC-3′. The housekeeping gene rplU (Kuchma et al., 2007) was used to normalize the gene expression data.
Results and discussion
Growth of P. aeruginosa on adenosine in minimal medium was determined in the presence of the QQ compound C-30. As expected, the growth yield at 48 h of P. aeruginosa in adenosine (0.1% w/v) minimal medium was decreased by 5±1-fold with 50 μm C-30 (Figure 1a), whereas there was no growth inhibition due to C-30 in LB medium (μ=1.4±0.1 h−1) as found previously (Hentzer et al., 2003). Hence, we successfully created a method by which there is selective pressure to evolve resistance to the antivirulence compound C-30 as it represses QS.
To confirm that C-30 represses QS phenotypes with P. aeruginosa; that is, it acts as a QQ compound, we chose five phenotypes that have been linked to QS with this strain: pyocyanin production (Ueda et al., 2009), pyoverdine production (Hentzer et al., 2003), elastase activity (Hentzer et al., 2003), swarming (Ueda et al., 2009) and biofilm formation (Ueda et al., 2009). Swarming was abolished by the addition of C-30 (Figure 2a), and biofilm formation was reduced by 3.7±0.7-fold with C-30 in glucose minimal medium and 2.3±0.5-fold in succinate minimal medium. Also, pyocyanin production was reduced by 1.49±0.04-fold upon addition of C-30 (Figure 2b), and as shown previously with C-30 (Hentzer et al., 2003), pyoverdine production was decreased by 2.4±0.4-fold and elastase was decreased by 8±2-fold. Hence, the synthesized C-30 was active and inhibited QS-related phenotypes with P. aeruginosa.
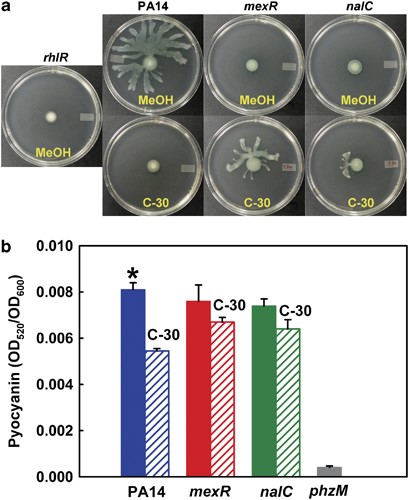
The mexR and nalC mutations decrease C-30 inhibition of P. aeruginosa QS phenotypes. (a) Swarming motility at 37 °C after 24 h using rhlR as a negative control. Three independent cultures were used for each strain, and two swarming plates were used for each culture. Representative plates are shown. (b) Pyocyanin production after incubation in LB medium for 9 h. Mutant phzM was used as a negative control. C-30 was used at 50 μm. Data represent the mean±s.d.; n=3. *P<0.01 by Student's t-test compared with the value in the presence of C-30.
To facilitate the evolution of P. aeruginosa for resistance to C-30, random transposon mutagenesis was performed. In this way, cells with mutations that allow for faster growth with C-30 could be identified rapidly by DNA sequencing from the transposon. Using minimal medium containing adenosine and C-30, mutants less sensitive to C-30 were identified by their faster growth in sequential shake flask cultures. After four sequential dilutions, the difference in the growth rates allowed putative C-30 mutants to be purified. After performing this procedure several times, for eight mutants, the insertion position of Tn5-luxAB transposon was determined to be in mexR, the gene for the repressor of the locus that encodes the MexAB-OprM multi-drug resistance efflux pump (Chen et al., 2010). Corroborating these results, four mutants had the insertion in nalC, which also encodes a negative regulator of mexAB-oprM. Note mexR was previously found to be induced by C-30 (Hentzer et al., 2003).
We also obtained two types of spontaneous mutants resistant to C-30 that did not rely on transposon mutagenesis using the same selection method. After 13 sequential dilutions, two types of spontaneous QQ quandary mutants (sQQQ) were obtained (Figure 1b): C-30 enhances growth with sQQQ1, and sQQQ3 grows more rapidly with and without C-30 than the PA14 wild-type strain.
To confirm that the mexR and nalC mutants were more resistant to furanone C-30, growth was checked in minimal adenosine medium. Although C-30 still inhibited growth of the mexR and nalC strains, the growth yield of these two strains was two- to three-fold higher than the wild-type strain after 50–60 h (Figure 1a). In addition, C-30 had little effect on the QS-related phenotypes of the mexR and nalC strains because its addition did not alter pyocyanin formation (Figure 2b). Also, the addition of C-30 enhanced the swarming motility of mexR and nalC mutants rather than inhibiting it as for the wild-type strain (Figure 2a). Hence, the mexR and nalC mutations render the cells less sensitive to the anti-QS effects of the brominated furanone C-30.
To compare the impact of C-30 on PA14 and mexR, a whole-transcriptome analysis was performed with planktonic cells of PA14 and mexR grown in adenosine minimal medium with C-30. C-30 was previously shown to repress the majority of QS-induced genes in P. aeruginosa (Hentzer et al., 2003). Addition of C-30 to mexR-induced 23 QS genes (Hentzer et al., 2003; Schuster et al., 2003; Wagner et al., 2003) relative to the wild-type strain with C-30 (Table 2); hence, the ability of C-30 to repress QS was reduced in the mexR mutant. These QS genes include lasR and rhlR that encode the two transcriptional regulators of the acyl-homoserine lactone system of P. aeruginosa (Schuster et al., 2003). In our whole-transcriptome analysis, the nuh gene, which encodes the nucleoside hydrolase that is required to assimilate adenosine (Heurlier et al., 2005), was slightly induced (1.2-fold). Because mexR strain grows better in adenosine medium than the wild-type in the presence of C-30, nuh should show elevated expression. To explore this further, we conducted quantitative real-time reverse transcription PCR and found that nuh was induced 6±3-fold in the mexR mutant relative to wild-type PA14 after addition of C-30 to minimal adenosine medium for 30 min. Therefore, these whole-transcriptome results are consistent with our hypothesis that the mexR mutation renders P. aeruginosa less sensitive to the QQ compound C-30.
List of induced quorum-sensing genes for planktonic cells of mexR vs PA14 wild-type
| PAO1 ID | PA14 ID | Gene name | Fold change | Description | References |
|---|---|---|---|---|---|
| Multidrug efflux operon mexAB-oprM | |||||
| PA0425 | PA14_05530 | mexA | 1.7 | Resistance-nodulation-cell division (RND) multidrug efflux membrane fusion protein | |
| PA0426 | PA14_05540 | mexB | 1.5 | RND multidrug efflux transporter | |
| PA0427 | PA14_05550 | oprM | 1.6 | Multiple antibiotic resistance efflux outer membrane protein | |
| Quorum-induced genes identified by previous studies | |||||
| PA1430 | PA14_45960 | lasR | 2.0 | Transcriptional regulator LasR | |
| PA3477 | PA14_19120 | rhlR | 3.2 | Transcriptional regulator RhlR | a, b, c |
| PA0105 | pA14_01290 | coxB | 2.0 | Cytochrome c oxidase, subunit II | b |
| PA0106 | pA14_01300 | coxA | 3.0 | Cytochrome c oxidase, subunit I | b |
| PA0107 | pA14_01310 | 2.1 | Probable cytochrome c assembly protein | b, c | |
| PA0108 | pA14_01320 | coIII | 2.3 | Cytochrome c oxidase, subunit III | b |
| PA1174 | PA14_49250 | napA | 2.0 | Periplasmic nitrate reductase protein NapA | |
| PA1175 | PA14_49230 | napD | 2.0 | Protein of periplasmic nitrate reductase | b |
| PA1176 | PA14_49220 | napF | 2.0 | Ferredoxin protein NapF | b |
| PA1245 | PA14_48140 | 2.0 | Hypothetical protein | b | |
| PA1247 | PA14_48100 | aprE | 2.1 | Alkaline protease secretion protein AprE | a, b |
| PA1887 | PA14_40110 | 2.0 | Hypothetical protein | ||
| PA1888 | PA14_40100 | 2.3 | Hypothetical protein | b | |
| PA2303 | PA14_34830 | 2.6 | Putative regulatory protein | a, b, c | |
| PA2306 | PA14_34800 | 2.1 | Putative transporter, LysE family | c | |
| PA2591 | PA14_30580 | 2.0 | Probable transcriptional regulator, LuxR family | a, b, c | |
| PA2747 | PA14_28600 | 2.0 | Hypothetical protein | a, b | |
| PA3032 | PA14_24860 | snr1 | 2.1 | Cytochrome c | b, c |
| PA3904 | PA14_13390 | 2.8 | Hypothetical protein | a, b, c | |
| PA4139 | PA14_10380 | 2.0 | Hypothetical protein | a, b | |
| PA4496 | PA14_58350 | 2.1 | Probable binding protein component of ABC transporter | c | |
| PA4677 | PA14_61870 | 2.3 | Hypothetical protein | a, b, c | |
| PA4738 | PA14_62680 | 2.0 | Hypothetical protein | a, b | |
| PA4739 | PA14_62690 | 2.6 | Hypothetical protein | a, b | |
| PA5482 | PA14_72370 | 2.1 | Hypothetical protein | a, b | |
| PAO1 ID | PA14 ID | Gene name | Fold change | Description | References |
|---|---|---|---|---|---|
| Multidrug efflux operon mexAB-oprM | |||||
| PA0425 | PA14_05530 | mexA | 1.7 | Resistance-nodulation-cell division (RND) multidrug efflux membrane fusion protein | |
| PA0426 | PA14_05540 | mexB | 1.5 | RND multidrug efflux transporter | |
| PA0427 | PA14_05550 | oprM | 1.6 | Multiple antibiotic resistance efflux outer membrane protein | |
| Quorum-induced genes identified by previous studies | |||||
| PA1430 | PA14_45960 | lasR | 2.0 | Transcriptional regulator LasR | |
| PA3477 | PA14_19120 | rhlR | 3.2 | Transcriptional regulator RhlR | a, b, c |
| PA0105 | pA14_01290 | coxB | 2.0 | Cytochrome c oxidase, subunit II | b |
| PA0106 | pA14_01300 | coxA | 3.0 | Cytochrome c oxidase, subunit I | b |
| PA0107 | pA14_01310 | 2.1 | Probable cytochrome c assembly protein | b, c | |
| PA0108 | pA14_01320 | coIII | 2.3 | Cytochrome c oxidase, subunit III | b |
| PA1174 | PA14_49250 | napA | 2.0 | Periplasmic nitrate reductase protein NapA | |
| PA1175 | PA14_49230 | napD | 2.0 | Protein of periplasmic nitrate reductase | b |
| PA1176 | PA14_49220 | napF | 2.0 | Ferredoxin protein NapF | b |
| PA1245 | PA14_48140 | 2.0 | Hypothetical protein | b | |
| PA1247 | PA14_48100 | aprE | 2.1 | Alkaline protease secretion protein AprE | a, b |
| PA1887 | PA14_40110 | 2.0 | Hypothetical protein | ||
| PA1888 | PA14_40100 | 2.3 | Hypothetical protein | b | |
| PA2303 | PA14_34830 | 2.6 | Putative regulatory protein | a, b, c | |
| PA2306 | PA14_34800 | 2.1 | Putative transporter, LysE family | c | |
| PA2591 | PA14_30580 | 2.0 | Probable transcriptional regulator, LuxR family | a, b, c | |
| PA2747 | PA14_28600 | 2.0 | Hypothetical protein | a, b | |
| PA3032 | PA14_24860 | snr1 | 2.1 | Cytochrome c | b, c |
| PA3904 | PA14_13390 | 2.8 | Hypothetical protein | a, b, c | |
| PA4139 | PA14_10380 | 2.0 | Hypothetical protein | a, b | |
| PA4496 | PA14_58350 | 2.1 | Probable binding protein component of ABC transporter | c | |
| PA4677 | PA14_61870 | 2.3 | Hypothetical protein | a, b, c | |
| PA4738 | PA14_62680 | 2.0 | Hypothetical protein | a, b | |
| PA4739 | PA14_62690 | 2.6 | Hypothetical protein | a, b | |
| PA5482 | PA14_72370 | 2.1 | Hypothetical protein | a, b | |
Cells were incubated in adenosine minimal medium containing 50 μm C-30. Raw data for the two DNA microarrays are available using GEO series accession number GSE24262.
a, (Hentzer et al., 2003); b, (Schuster et al., 2003); c, (Wagner et al., 2003).
List of induced quorum-sensing genes for planktonic cells of mexR vs PA14 wild-type
| PAO1 ID | PA14 ID | Gene name | Fold change | Description | References |
|---|---|---|---|---|---|
| Multidrug efflux operon mexAB-oprM | |||||
| PA0425 | PA14_05530 | mexA | 1.7 | Resistance-nodulation-cell division (RND) multidrug efflux membrane fusion protein | |
| PA0426 | PA14_05540 | mexB | 1.5 | RND multidrug efflux transporter | |
| PA0427 | PA14_05550 | oprM | 1.6 | Multiple antibiotic resistance efflux outer membrane protein | |
| Quorum-induced genes identified by previous studies | |||||
| PA1430 | PA14_45960 | lasR | 2.0 | Transcriptional regulator LasR | |
| PA3477 | PA14_19120 | rhlR | 3.2 | Transcriptional regulator RhlR | a, b, c |
| PA0105 | pA14_01290 | coxB | 2.0 | Cytochrome c oxidase, subunit II | b |
| PA0106 | pA14_01300 | coxA | 3.0 | Cytochrome c oxidase, subunit I | b |
| PA0107 | pA14_01310 | 2.1 | Probable cytochrome c assembly protein | b, c | |
| PA0108 | pA14_01320 | coIII | 2.3 | Cytochrome c oxidase, subunit III | b |
| PA1174 | PA14_49250 | napA | 2.0 | Periplasmic nitrate reductase protein NapA | |
| PA1175 | PA14_49230 | napD | 2.0 | Protein of periplasmic nitrate reductase | b |
| PA1176 | PA14_49220 | napF | 2.0 | Ferredoxin protein NapF | b |
| PA1245 | PA14_48140 | 2.0 | Hypothetical protein | b | |
| PA1247 | PA14_48100 | aprE | 2.1 | Alkaline protease secretion protein AprE | a, b |
| PA1887 | PA14_40110 | 2.0 | Hypothetical protein | ||
| PA1888 | PA14_40100 | 2.3 | Hypothetical protein | b | |
| PA2303 | PA14_34830 | 2.6 | Putative regulatory protein | a, b, c | |
| PA2306 | PA14_34800 | 2.1 | Putative transporter, LysE family | c | |
| PA2591 | PA14_30580 | 2.0 | Probable transcriptional regulator, LuxR family | a, b, c | |
| PA2747 | PA14_28600 | 2.0 | Hypothetical protein | a, b | |
| PA3032 | PA14_24860 | snr1 | 2.1 | Cytochrome c | b, c |
| PA3904 | PA14_13390 | 2.8 | Hypothetical protein | a, b, c | |
| PA4139 | PA14_10380 | 2.0 | Hypothetical protein | a, b | |
| PA4496 | PA14_58350 | 2.1 | Probable binding protein component of ABC transporter | c | |
| PA4677 | PA14_61870 | 2.3 | Hypothetical protein | a, b, c | |
| PA4738 | PA14_62680 | 2.0 | Hypothetical protein | a, b | |
| PA4739 | PA14_62690 | 2.6 | Hypothetical protein | a, b | |
| PA5482 | PA14_72370 | 2.1 | Hypothetical protein | a, b | |
| PAO1 ID | PA14 ID | Gene name | Fold change | Description | References |
|---|---|---|---|---|---|
| Multidrug efflux operon mexAB-oprM | |||||
| PA0425 | PA14_05530 | mexA | 1.7 | Resistance-nodulation-cell division (RND) multidrug efflux membrane fusion protein | |
| PA0426 | PA14_05540 | mexB | 1.5 | RND multidrug efflux transporter | |
| PA0427 | PA14_05550 | oprM | 1.6 | Multiple antibiotic resistance efflux outer membrane protein | |
| Quorum-induced genes identified by previous studies | |||||
| PA1430 | PA14_45960 | lasR | 2.0 | Transcriptional regulator LasR | |
| PA3477 | PA14_19120 | rhlR | 3.2 | Transcriptional regulator RhlR | a, b, c |
| PA0105 | pA14_01290 | coxB | 2.0 | Cytochrome c oxidase, subunit II | b |
| PA0106 | pA14_01300 | coxA | 3.0 | Cytochrome c oxidase, subunit I | b |
| PA0107 | pA14_01310 | 2.1 | Probable cytochrome c assembly protein | b, c | |
| PA0108 | pA14_01320 | coIII | 2.3 | Cytochrome c oxidase, subunit III | b |
| PA1174 | PA14_49250 | napA | 2.0 | Periplasmic nitrate reductase protein NapA | |
| PA1175 | PA14_49230 | napD | 2.0 | Protein of periplasmic nitrate reductase | b |
| PA1176 | PA14_49220 | napF | 2.0 | Ferredoxin protein NapF | b |
| PA1245 | PA14_48140 | 2.0 | Hypothetical protein | b | |
| PA1247 | PA14_48100 | aprE | 2.1 | Alkaline protease secretion protein AprE | a, b |
| PA1887 | PA14_40110 | 2.0 | Hypothetical protein | ||
| PA1888 | PA14_40100 | 2.3 | Hypothetical protein | b | |
| PA2303 | PA14_34830 | 2.6 | Putative regulatory protein | a, b, c | |
| PA2306 | PA14_34800 | 2.1 | Putative transporter, LysE family | c | |
| PA2591 | PA14_30580 | 2.0 | Probable transcriptional regulator, LuxR family | a, b, c | |
| PA2747 | PA14_28600 | 2.0 | Hypothetical protein | a, b | |
| PA3032 | PA14_24860 | snr1 | 2.1 | Cytochrome c | b, c |
| PA3904 | PA14_13390 | 2.8 | Hypothetical protein | a, b, c | |
| PA4139 | PA14_10380 | 2.0 | Hypothetical protein | a, b | |
| PA4496 | PA14_58350 | 2.1 | Probable binding protein component of ABC transporter | c | |
| PA4677 | PA14_61870 | 2.3 | Hypothetical protein | a, b, c | |
| PA4738 | PA14_62680 | 2.0 | Hypothetical protein | a, b | |
| PA4739 | PA14_62690 | 2.6 | Hypothetical protein | a, b | |
| PA5482 | PA14_72370 | 2.1 | Hypothetical protein | a, b | |
Cells were incubated in adenosine minimal medium containing 50 μm C-30. Raw data for the two DNA microarrays are available using GEO series accession number GSE24262.
a, (Hentzer et al., 2003); b, (Schuster et al., 2003); c, (Wagner et al., 2003).
To further characterize the mexR mutant, we investigated its pathogenicity using Caenorhabditis elegans (Mahajan-Miklos et al., 1999) as the animal model. It has been shown that the C. elegans fast-killing model is mediated by pyocyanin and other toxins, which are also required for pathogenesis in plants and mice (Mahajan-Miklos et al., 1999). In addition, because pyocyanin is QS controlled and inhibited by C-30 in wild-type PA14, this model is related to QS. Our goals were to determine if the mexR mutant could still be pathogenic as well as whether it was more virulent, compared with the wild-type strain, in the presence of the QS inhibitor C-30. L4 stage hermaphrodite worms were exposed to lawns of PA14 and mexR grown on PGS agar plates with or without C-30. A mexA mutant with reduced MexAB-OprM efflux pump activity was found previously to be less virulent to C. elegans in a similar assay (Mahajan-Miklos et al., 1999), and our results indicate that the mexR mutant with enhanced MexAB-OprM efflux pump activity is as virulent as the wild-type strain: 50∼60% of the worms were killed within 10 h for both strains (Figure 3). Critically, C-30 reduced wild-type PA14 virulence by 2.8±0.5-fold whereas it had little effect on the mexR mutant. In contrast, worms exposed to E. coli OP50 on the same medium plates showed 100% viability. These results indicate that the mexR mutant remains pathogenic under the conditions we tested and that it is much more virulent in the presence of the QS inhibitor than the wild-type strain.
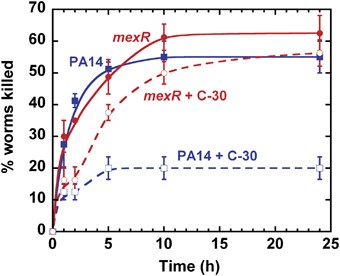
The mexR mutation decreases C-30 inhibition of killing of C. elegans by P. aeruginosa. L4 stage hermaphrodite worms were exposed to wild-type P. aeruginosa (blue squares) and mexR (red circles) grown on PGS medium with (open symbols) or without (closed symbols) 50 μm C-30. Worms exposed to E. coli OP50 on the same medium plates were used as the negative control, which showed 100% viability. Data represent the mean±s.d.; n=4 plates, 20 worms per plate.
To corroborate these C. elegans results with the C-30-resistant mexR and nalC mutants obtained from growth on adenosine, clinical isolates from patients with chronic CF infections (Tomás et al., 2010) were investigated for their resistance to C-30. These P. aeruginosa strains have mutations that include mexR and nalC (Tomás et al., 2010); such mutations arise frequently in chronic but not in acute infections (Oliver and Mena, 2010). For the nine strains we checked (Table 1), strain 12142 with the highest mexA induction (15.9-fold) showed no growth defect in the presence of C-30 (Figure 1c), and strain 1253 had increased growth in the presence of C-30. In contrast, growth yield of the control strain AHP (Tomás et al., 2010) with reduced mexA expression (8.3-fold) was reduced 4.2±0.7-fold by C-30. Hence, natural isolates in CF patients have obtained mutations, which include mexR and nalC, and these natural CF isolates are also resistant to the QQ compound C-30.
MexR is the primary negative regulator of the MexAB–OprM complex, which exports diverse antibiotics, including quinolones, β-lactams, tetracycline, chloramphenicol, novobiocin, macrolides and biocides (Chen et al., 2010); this pump also exports the 3OC12-HSL of the P. aeruginosa Las QS system (Pearson et al., 1999). As a possible mechanism for the resistance of the mexR mutant to the QS quenching C-30, we hypothesized that inactivation of the mexR gene led to increased efflux of C-30 by the P. aeruginosa cells; thereby, the inefficiency of C-30 as an antivirulence compound in the mexR mutant may be because of it being exported, which limits its effect on the QS pathways. Indeed, the three genes in mexAB-oprM operon were induced slightly in the mexR mutant upon addition of C-30 (Table 2). More significantly, the minimum inhibitory concentration of chloramphenicol, an antibiotic known to be effluxed by the MexAB–OprM complex (Chen et al., 2010), was increased 3.5±0.4-fold for the C-30-resistant, mexR mutant. Furthermore, both the mexR transposon mutant and the wild-type strain had similar uptake activities of C-30 (229±7 nmol mg−1 and 250±13 nmol mg−1 protein, respectively) after 30 min; however, only the mexR mutant effluxed C-30 in minimal succinate medium (3±2 nmol h−1 mg−1 protein) as there was no efflux found with the wild-type strain (20±10% of the C-30 imported into the cells was exported by the mexR mutant after 15 h). To avoid the possibility of nonspecific binding of C-30 to the cells, the concentration of C-30 was measured after washing and no C-30 was detected for either strain. In addition, there was no measurable efflux by either strain in buffer, which served as a negative control for non-active release. Therefore, the mexR mutation renders the cells more resistant to C-30 because of its increased export.
It was not anticipated that the antibiotic efflux pump MexAB-OprM may export a QQ compound. Although the MexAB-OprM pump is able to export the 3OC12-HSL signal (Pearson et al., 1999), it does not export the C4-HSL signal (Pearson et al., 1999), which has a shorter N-acyl side chain than 3OC12-HSL, and the shorter C4-HSL signal resembles to C-30 more than 3OC12-HSL. Hence, the discovery of the QQ-resistance mechanism found here because of increased efflux activity is novel.
In conclusion, our results show that bacteria can obtain resistance to antivirulence compounds such as the best-studied brominated furanones. The overlap of the resistance to the antivirulence compounds and the resistance to antibiotics for the C-30-resistant mutants indicates that these kinds of mutants are easily obtained. These mutants with increased MexAB-OprM efflux activities would evolve not only under adenosine-usage selective pressure, but also under antibiotic treatment, which constitute a very strong selective pressure. Hence, treatments with antibiotics may induce resistance to antivirulence compounds (and perhaps vice versa). Corroborating this hypothesis, we also demonstrated that natural isolates from chronic infections also are resistant to the QQ compound C-30 as we showed that some of the isolates that arise in CF patients (those with the mexR and nalC mutations) are resistant to C-30. Hence, these results are important for the development of rational strategies for the utilization of antivirulence compounds for the clinical treatment of infectious diseases in that clinicians should take into account the possibility of a rapid selection and dissemination of resistant bacteria by diverse resistance mechanisms.
Acknowledgements
This work was supported by the NIH (R01 GM089999). We thank Dr N Woodford and Dr D Livermore from the Centre for Infections, Health Protection Agency, United Kingdom, for their help with the clinical CF isolates. TM is grateful for the Excellent Young Researchers Overseas Visit Program Fellowship of the Japan Society for the Promotion of Science. RG-C was supported by the SEP/CONACyT grant 152794. TW is the T Michael O’Connor II Endowed Chair and Professor at the Texas A&M University.
Data availability
Accession codes
Accessions
Gene Expression Omnibus
References
Author notes
Toshinari Maeda, Rodolfo García-Contreras and Mingming Pu: These authors contributed equally to this work.

{kind=link}
{kind=link}
{kind=link}