





Abstract
(R)-Ketamine exhibits rapid and sustained antidepressant effects in animal models of depression. It is stereoselectively metabolized to (R)-norketamine and subsequently to (2R,6R)-hydroxynorketamine in the liver. The metabolism of ketamine to hydroxynorketamine was recently demonstrated to be essential for ketamine’s antidepressant actions. However, no study has compared the antidepressant effects of these 3 compounds in animal models of depression.
The effects of a single i.p. injection of (R)-ketamine, (R)-norketamine, and (2R,6R)-hydroxynorketamine in a rat learned helplessness model were examined.
A single dose of (R)-ketamine (20 mg/kg) showed an antidepressant effect in the rat learned helplessness model. In contrast, neither (R)-norketamine (20 mg/kg) nor (2R,6R)-hydroxynorketamine (20 and 40 mg/kg) did so.
Unlike (R)-ketamine, its metabolite (2R,6R)-hydroxynorketamine did not show antidepressant actions in the rat learned helplessness model. Therefore, it is unlikely that the metabolism of ketamine to hydroxynorketamine is essential for ketamine’s antidepressant actions.
The rapid and sustained antidepressant effects of ketamine in patients with treatment-resistant depression are the most important discovery in the field of depression research in a half-century. However, the precise mechanisms underlying the antidepressant effects of ketamine remain unknown. A recent study (Zanos et al., 2016) reported that the metabolism of ketamine to hydroxynorketamine (HNK) is essential for ketamine’s antidepressant effects. In particular, (2R,6R)-HNK, a metabolite of (R)-ketamine, plays a key role in the antidepressant actions. However, here we report that, unlike (R)-ketamine, its metabolites (R)-norketamine and (2R,6R)-HNK did not elicit antidepressant effects in a rat learned helplessness model. It is, therefore, unlikely that the metabolism of ketamine to HNK is necessary for ketamine’s antidepressant actions.
Introduction
Recently conducted meta-analyses revealed that the N-methyl-D-aspartate receptor antagonist ketamine exhibits rapid and sustained antidepressant effects in patients with treatment-resistant depression (Newport et al, 2015; Kishimoto et al, 2016). Thus, ketamine is the most attractive antidepressant for the treatment of treatment-resistant depression (Monteggia and Zarate, 2015; Duman et al., 2016; Hashimoto, 2016b), although the precise mechanisms underlying its antidepressant actions remain unknown.
(R,S)-Ketamine is a racemic mixture containing equal parts of (R)-ketamine and (S)-ketamine. (S)-Ketamine shows approximately 3- to 4-fold greater anesthetic potency and greater undesirable psychotomimetic side effects than (R)-ketamine (Domino et al., 2010). Several groups including our own have demonstrated that (R)-ketamine showed greater potency and longer-lasting antidepressant effects than (S)-ketamine in animal models of depression (Zhang et al., 2014; Yang et al., 2015, 2017, 2018; Zanos et al., 2016; Fukumoto et al., 2017). Unlike (S)-ketamine, (R)-ketamine does not induce psychotomimetic side effects or exhibit abuse potential in rodents (Yang et al., 2015). Furthermore, single or repeated intermittent administration of (S)-ketamine, but not of (R)-ketamine, resulted in the loss of parvalbumin immunoreactivity in the prefrontal cortex and hippocampus (Yang et al., 2015, 2016). Moreover, with the results using [11C]raclopride and positron emission tomography, we reported a marked reduction of dopamine D2/3 receptor binding in the conscious monkey striatum after a single infusion of (S)-ketamine, but not of (R)-ketamine (Hashimoto et al., 2017). These findings suggest that (S)-ketamine, but not (R)-ketamine, can cause a marked release of dopamine from presynaptic terminals, which is associated with acute psychotomimetic effects (Hashimoto et al., 2017). Taking these findings together, (R)-ketamine could be a potentially safer antidepressant without detrimental side effects in humans than (S)-ketamine (Hashimoto, 2016a, 2016b, 2017).
It is well known that ketamine is rapidly metabolized into norketamine and subsequently into hydroxynorketamine (HNK) by microsomal cytochrome P450 enzymes (through N-demethylation and hydroxylation) in the liver (Figure 1) (Turfus et al., 2009; Zhao et al., 2012; Zanos et al., 2016; Hashimoto, 2017). The metabolism of ketamine to HNK was also recently demonstrated to be essential for the antidepressant actions of ketamine (Zanos et al., 2016). In particular, (2R,6R)-HNK, a metabolite from (R)-ketamine, plays a key role in the antidepressant actions (Zanos et al., 2016). However, increasing attention has been drawn to the antidepressant actions of (2R,6R)-HNK (Abdallah, 2017). In the present study, we examined the effects of a single systemic injection of (R)-ketamine and its two major metabolites, (R)-norketamine and (2R,6R)-HNK, in a rat learned helplessness (LH) model.
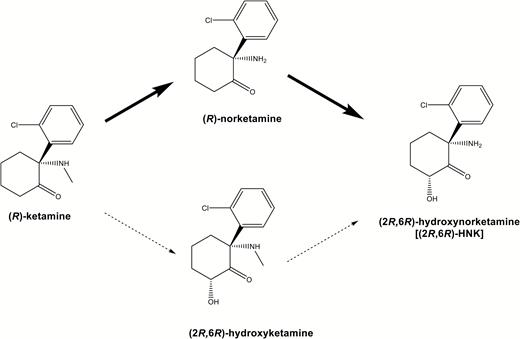
Metabolism of (R)-ketamine in the liver. In the liver, (R)-ketamine is metabolized to (R)-norketamine (major pathway) and (2R,6R)-hydroxyketamine (minor pathway), subsequently (2R,6R)-hydroxynorketamine (HNK).
Methods and Materials
Animals
Male Sprague-Dawley rats (200–230 g, 7 weeks old; Charles-River Japan) were used. The animals were housed under a 12-h-light/-dark cycle with free access to food and water. The protocol was approved by the Chiba University Institutional Animal Care and Use Committee (permission no: 28–394 and 29–328). All efforts were made to minimize suffering.
Drugs
(R)-Ketamine hydrochloride was prepared by recrystallization of (R,S)-ketamine (Ketalar, ketamine hydrochloride, Daiichi Sankyo Pharmaceutical Ltd) and D-(-)-tartaric acid, as described previously (Zhang et al., 2014). (R)-Norketamine hydrochloride was prepared as described previously (Zanos et al., 2016). The purity of these stereoisomers was determined by a high-performance liquid chromatography (CHIRALPAK IA, column size: 250x4.6 mm, mobile phase: n-hexane/dichloromethane/diethylamine (75/25/0.1), Daicel Corporation). (2R,6R)-HNK hydrochloride was provided from Taisho Pharmaceutical Ltd as reported previously (Zanos et al., 2016). (R)-Ketamine, (R)-norketamine, and (2R,6R)-HNK were dissolved in 0.9% NaCl. Other compounds were purchased commercially. The doses of (R)-ketamine and its metabolites were selected as previously reported (Yang et al., 2015, 2017; Zanos et al., 2016).
Stress Paradigm (LH Model)
To create an LH paradigm, the animals are initially exposed to uncontrollable stress. When the animal is later placed in a situation where the shock is controllable (escapable), the animal not only fails to acquire the escape response but also often makes no efforts to escape the shock at all. The LH behavioral tests were performed using the Gemini Avoidance System (San Diego Instruments) (Shirayama et al., 2015, 2017). This apparatus is divided into 2 compartments by a retractable door. On days 1 and 2, the rats were subjected to 30 inescapable electric foot-shocks (0.65 mA, 30-second duration, administered at random intervals averaging 18–42 seconds) (Figure 2A, C). On day 3, a 2-way conditioned avoidance test was performed as a post-shock test to determine whether the rats would exhibit the predicted escape deficits (Figure 2A, C). This screening session consisted of 30 trials in which electric foot-shocks (0.65 mA, 6-second duration, administered at random intervals with a mean of 30 seconds) were preceded by a 3-second conditioned stimulus tone that remained on until the shock was terminated. Rats with more than 25 escape failures among the 30 trials were regarded as having reached the LH criterion and were used in further experiments. Approximately 65% of the rats met this criterion.
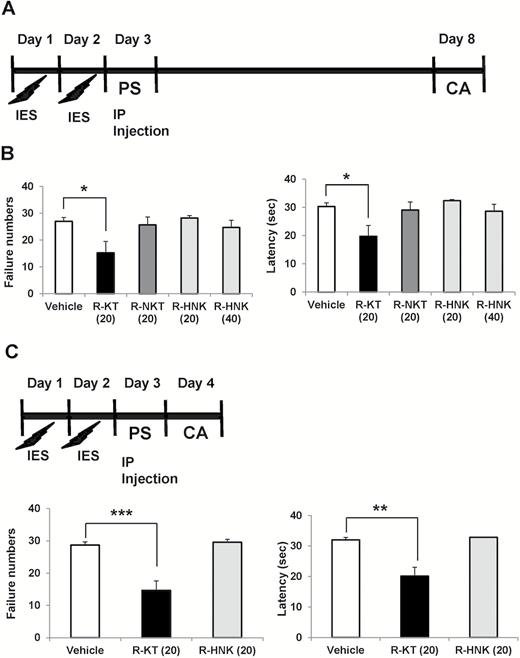
Effects of a single injection of (R)-ketamine, (R)-norketamine, and (2R,6R)- HNK in a rat LH model. (A) Rats received inescapable electric shock (IES) treatments on 2 days (days 1 and 2), passed a post-shock test (PS) on day 3, and were designated as learned helplessness (LH) rats with depression-like phenotype. On day 3, vehicle (saline: 2 mL/kg), (R)-ketamine (20 mg/kg), (R)-norketamine (20 mg/kg), or (2R,6R)- HNK (20 and 40 mg/kg) was administered i.p. into LH rats. On day 8 (5 days after a single injection), conditioned avoidance (CA) test to study the antidepressant effect was performed. (B) The failure number of LH (1-way ANOVA: F4,24 = 3.755, P = .0167). The escape latency of LH (1-way ANOVA: F4,24 = 3.973, P = .013). Data are shown as mean ± SEM (n = 5–8). The number in the parenthesis is the dose (mg/kg). *P < .05 compared with vehicle-treated group. (C) Rats received IES treatments on 2 days (days 1 and 2), passed a PS on day 3, and were designated as LH rats with depression-like phenotype. On day 3, either vehicle (saline: 2 mL/kg), (R)-ketamine (20 mg/kg), or (2R,6R)- HNK (20 mg/kg) was administered i.p. into LH rats. CA test was performed on day 4 (24 hours after a single injection). (D) The failure number of LH (1-way ANOVA: F2,14 = 13.52, P < .0001). The escape latency of LH (1-way ANOVA: F2,14 = 14.73, P = .0004). Data are shown as mean ± SEM (n = 5 or 6). The number in the parenthesis is the dose (mg/kg). **P < .01, ***P < .001 compared with vehicle-treated group. R-KT: (R)-ketamine, R-NKT: (R)-norketamine, R-HNK: (2R,6R)-hydroxynorketamine.
In the experiment 1, on day 3, the rats received i.p. injection of saline (2 mL/kg), (R)-ketamine (20 mg/kg), (R)-norketamine (20 mg/kg), or (2R,6R)-HNK (20 and 40 mg/kg) (Figure 2A). On day 8 (5 days after a single injection), a 2-way conditioned avoidance test was performed (Figure 2A). In the experiment 2, on day 3, the rats received i.p. injection of saline (2 mL/kg), (R)-ketamine (20 mg/kg), or (2R,6R)-HNK (20mg/kg) (Figure 2C). On day 4 (24 hours after a single injection), a 2-way conditioned avoidance test was performed (Figure 2C). This test session consisted of 30 trials in which electric foot-shocks (0.65 mA, 30-second duration, administered at random intervals with a mean of 30 seconds) were preceded by a 3-second conditioned stimulus tone that remained on until the shock was terminated. The number of escape failures and the latency until escape for each of the 30 trials were recorded by the Gemini Avoidance System.
Statistical Analysis
The data are shown as the mean ± SEM. The analyses were performed using GraphPad Prism 5 (GraphPad Software Inc). The data were analyzed using 1-way ANOVA, followed by posthoc Tukey test. The criterion for significance was P<.05.
Results
Effects of a Single Intraperitoneal Injection of (R)-Ketamine, (R)-Norketamine, and (2R,6R)-HNK in LH Rats
Experiment 1
To examine the antidepressant effects of (R)-ketamine and its 2 metabolites in a LH model, saline (2 mL/kg), (R)-ketamine (20 mg/kg), (R)-norketamine (20 mg/kg), or (2R,6R)-HNK (20 and 40 mg/kg) was administered i.p. into the LH rats. The LH rats that received a single injection of (R)-ketamine (20 mg/kg, 5 days after a single injection) exhibited significant improvements in their conditioned avoidance test results, relative to vehicle-treated LH rats (Figure 2A,B). In contrast, a single administration of (R)-norketamine (20 mg/kg), or (2R,6R)-HNK (20 and 40 mg/kg) did not improve their conditioned avoidance test results in LH rats (Figure 2A,B).
Experiment 2
The LH rats that received a single injection of (R)-ketamine (20 mg/kg, 24 hours after a single injection) exhibited significant improvements in their conditioned avoidance test results relative to vehicle-treated LH rats (Figure 2C,D). In contrast, (2R,6R)-HNK (20 mg/kg, 24 hours after a single injection) did not improve their conditioned avoidance test results in LH rats (Figure 2C,D).
Discussion
In the present study, we established that a single systemic (R)-ketamine injection (24 hours and 5 days after a single injection) showed antidepressant effects in a rat LH model of depression, whereas a single systemic injection of (R)-norketamine or (2R,6R)-HNK did not. It should be noted that a higher dose (40 mg/kg) of (2R,6R)-HNK did not have antidepressant effects. We have also recently reported the potent and longer-lasting antidepressant effects of (R)-ketamine in a social defeat stress model of depression, although (2R,6R)-HNK did not have antidepressant effects (Yang et al., 2017). Collectively, it seems that unlike (R)-ketamine, (2R,6R)-HNK does not have an antidepressant effect in rodent models of depression, inconsistent with the findings by Zanos et al. (2016).
Zanos et al. (2016) reported more potent antidepressant effects of (2R,6R)-HNK, which is exclusively derived from (R)-ketamine. A single injection of (2R,6R)-HNK (10 or 20 mg/kg) reversed chronic corticosterone-induced anhedonia assessed with the sucrose preference and female urine sniffing behavioral tasks as well as social avoidance induced by chronic social defeat stress (Zanos et al., 2016). They reported sustained (24 hours) antidepressant effects of (2R,6R)-HNK in LH model (Zanos et al, 2016). However, we could not find sustained (24 hours) antidepressant effects of (2R,6R)-HNK (20 mg/kg) in the LH model, although (R)-ketamine (20 mg/kg) showed sustained (24 hours) antidepressant effects in the same model (Figure 2D). Thus, we were unable to detect antidepressant activity induced by (2R,6R)-HNK in any of our 3 models (inflammation, social defeat stress, and LH), although rapid and sustained antidepressant effects were detected for (R)-ketamine (Yang et al., 2017; this study). The reasons for this discrepancy (Zanos et al., 2016, vs Yang et al., 2017, and this study) remain unknown. Nonetheless, our negative findings regarding the lack of an antidepressant effect of (2R,6R)-HNK in rodents with depression-like phenotype need to be replicated by other groups in future studies.
It is well known that (R)-ketamine is stereoselectively N-demethylated by liver microsomal cytochrome P450 into (R)-norketamine (Hijazi and Boulieu, 2002; Desta et al., 2012; Zanos et al., 2016) (Figure 1). (R)-Norketamine is further metabolized to (2R,6R)-HNK arising from hydroxylation of the cyclohexanone ring (Figure 1). In addition to N-demethylation, (R)-ketamine is also metabolized by the hydroxylation of the cyclohexanone ring to produce (2R,6R)-hydroxyketamine. (2R,6R)-HNK is also prepared by the N-demethylation of (2R,6R)-hydroxyketamine (Desta et al., 2012; Zanos et al., 2016) (Figure 1). A study showed that the plasma levels of norketamine and HNK were higher after a single injection of ketamine, although plasma levels of hydroxyketamine were very low (Zanos et al., 2016), suggesting that the metabolism of (R)-ketamine to (2R,6R)-HNK via (R)-norketamine is the major pathway of (R)-ketamine in mice (Figure 1). Therefore, it is noteworthy that (R)-norketamine and (2R,6R)-HNK, the two major metabolites from (R)-ketamine, did not exert antidepressant effects, although antidepressant effects of (R)-ketamine were detected in the same model.
We have recently reported that a single bilateral infusion of (R)-ketamine into the infralimbic (IL) area of the medial prefrontal cortex (mPFC) and the DG and CA3 of the hippocampus exerted antidepressant effects in LH rats (Shirayama and Hashimoto, 2017). Furthermore, a study showed that neuronal inactivation of the IL of mPFC completely blocked the antidepressant effects of (R,S)-ketamine, and that microinfusion of (R,S)-ketamine into IL of mPFC produced an antidepressant effect in control unstressed rats (Fuchikami et al., 2015). These findings suggest a crucial role for the IL area of the mPFC, DG, and CA3 in the antidepressant action of (R)-ketamine itself (NOT metabolite) in a rat LH model, since (R)-ketamine (or (R,S)-ketamine) might not be metabolized in the brain. Given the key role of hepatic cytochrome P450 enzymes in ketamine metabolism (Turfus et al., 2009; Zhao et al., 2012; Zanos et al., 2016), it is unlikely that the metabolism of (R)-ketamine in the liver plays a role in the antidepressant actions of (R)-ketamine.
In conclusion, unlike (R)-ketamine, neither (R)-norketamine nor (2R,6R)-HNK elicited antidepressant effects in LH model, suggesting that the metabolism of (R)-ketamine might not play a key role in its robust antidepressant action.
Statement of Interest
Dr. Shirayama has received research support from Eli Lilly, Eisai, MSD, Pfizer, and Mitsubishi-Tanabe. Dr. Hashimoto has received research support from Dainippon-Sumitomo, Mochida, Otsuka, and Taisho. Dr. Hashimoto is an inventor on a filed patent application on “The use of (R)-ketamine in the treatment of psychiatric diseases” by Chiba University.
Acknowledgments
We thank Dr. Shigeyuki Chaki (Taisho Pharmaceutical Co, Ltd) for providing (2R,6R)-HNK. We also thank Yuko Fujita (Chiba University) for her technical assistance.
This study was supported by Strategic Research Program for Brain Sciences, AMED, Japan (to K.H.).
References

{kind=link}
{kind=link}