





Abstract
Global warming is enabling tropical invaders to expand into regions, where unsuitable climate conditions used to prevent the survival or reproduction. Here, using the occurrence data of 22 Lessepsian fish in the Mediterranean Sea, we aimed at (1) gaining a deeper understanding of temperature match dynamics over the spatio-temporal spread of each invasion event and (2) testing the hypothesis that successful invasions require a temperature match, as assumed in several bioinvasion risk assessments. For each species, we identified the areas of the Mediterranean lacking a temperature match with native ranges over the past six decades. We found that some species, particularly recent invaders, have spread far beyond temperature matching conditions. The populations at the expanding edge of the range are often found in sites that have experienced increases in temperature. Temperature match positively correlates with species spread rate and after 1990, higher temperatures in the Mediterranean provided analogous conditions for a growing number of non-indigenous species. Our results warn against the general use of temperature-based climate matching in bioinvasion risk and provide guidance for the application of species distribution models that incorporate temperature to marine bioinvasion research.
Introduction
Global warming has already significantly altered the distribution of Earth's biota (Pecl et al., 2017) by enabling a large number of non-indigenous species to expand into regions where they previously could not survive and/or reproduce (Walther et al., 2009; Bellard et al., 2013; Rius et al., 2014). In particular, the spread of warm-adapted species is facilitated, whose ranges are forecast to become broader and impacts to be increasingly apparent in the next few decades (McInerney et al., 2021). Intense work is being conducted to understand the relationship between species distribution and climate with a view to predicting the potential spread of biological invaders, and the corresponding risk, in a warming world (Hobbs, 2000). Most risk assessment approaches rely on climate matching—which evaluates whether the climatic conditions of the recipient region resemble (i.e. are analogous to) those of the species’ native range (Hayes and Barry, 2008; Abellán et al., 2017)—as a key predictor of establishment success (Kumschick and Richardson, 2013) and potential invasiveness (Vilizzi et al., 2021). A climate match has been shown to correlate positively with the success or the invaded range size of a variety of non-indigenous bird, reptile, amphibian, and freshwater fish taxa (Hayes and Barry, 2008; Bomford et al., 2009; Bomford et al., 2010; Mahoney et al., 2015; Broennimann et al., 2021), and the major role of climate change as a driver for future alien species distribution has been recently reaffirmed (Bellard et al., 2018). However, recent studies have found that some species invade areas with different (non-analogous) climatic conditions compared with those of their native range (Border et al., 2018). It would thus be useful to identify the cases in which climate matching fails to predict the risk of bioinvasion of areas outside the known natural distribution of a species (Abellán et al., 2017).
Climate matching also plays an important role when fitting species distribution models (Elith and Leathwick, 2009; Guisan et al., 2017) for non-indigenous species because it may determine extrapolation. Extrapolation occurs when model predictions are produced not in the range of the sample data used for calibration, due to a lack of analogous conditions and it can cause biased results (Mesgaran et al., 2014). The same consideration applies to future conditions, which may result in the emergence of non-analogue climate (Mahony et al., 2017). In such cases, species face novel conditions, a situation that has been denominated “niche pioneering” (Atwater et al., 2018). Recently, niche pioneering has been explained as reflecting the fact that portions of a species’ fundamental ecological niche can be observed only in invaded areas, i.e. as a simple distributional expansion within the constraints of the niche (Qiao et al., 2017). Accordingly, niche pioneering can bias the results of those species distribution models that are based only on the known distribution in the realized niche.
Despite its significance and implications, to the best of our knowledge climate matching has not been tested in the marine environment. To do this, we exploited the unique situation of the Mediterranean Sea, a hotspot of climate change and biological invasions (MedECC, 2020) which in the past century has undergone major biotic transformations. The opening of the Suez Canal in 1869 allowed hundreds of species to pass from the Red Sea into the Mediterranean basin, a phenomenon known as Lessepsian migration. The more than 100 Lessepsian fish recorded so far (Golani, 2021) account for no less than 22% of the total number of fish species in the basin (Kovačić et al., 2021). This makes the Mediterranean the most heavily invaded marine region in the world (Edelist et al., 2013) as well as a unique and serendipitous testbed for bioinvasion research. These invaders are warm-adapted species, many of which have generated large populations in the easternmost (and warmest) area of the Mediterranean, known as Levantine Basin (e.g. Edelist et al., 2013; Galil et al., 2014). Such species, which have had dramatic impacts on native biota (Arndt et al., 2018), habitats (Sala et al., 2011; Vergés et al., 2014), and ecosystem services (Katsanevakis et al., 2014), are believed to require favourable climatic conditions to spread further into new Mediterranean sectors (e.g. Hiddink et al., 2012; Azzurro et al., 2019; D'Amen and Azzurro, 2020a).
Lessepsian species thriving in previously unencountered environmental conditions have been reported in niche shift studies. Notably, Parravicini et al. (2015) have described a tendency of several Lessepsian fish to expand their climatic niche. More recently, D'Amen and Azzurro (2020b), in their analysis of the climatic niche of the dusky spinefoot, Siganus luridus, described a niche pioneering event in relation to two temperature-related variables and stressed the need for a greater knowledge of each axis of the Grinnellian niche.
Based on this last evidence, and because temperature is considered the main key-factor driving colonization and establishment for non-indigenous species, especially under warming conditions (e.g. Raitsos et al., 2010; Sorte et al., 2010; Dijkstra et al., 2018), here we tested the importance of a temperature-match for the successful establishment of Lessepsian fish. The study has two main goals. The first was to gain a deeper understanding of temperature match dynamics over the spatio-temporal spread of each invasion event, i.e. to investigate how much more “invader-friendly” the Mediterranean has become for Red Sea species over time. This aspect was addressed by analysing the records of 22 Lessepsian fish species over six decades in a case study involving the entire Mediterranean Sea. The second objective was to verify whether Lessepsian invaders require a temperature match, as assumed in several bioinvasion risk assessments (e.g. Iacarella et al., 2015; Patoka et al., 2017). This we did by comparing the temperature records in the invaded and the native ranges of the species.
Material and methods
Occurrence data
The current and historical distribution of Lessepsian fish species was reconstructed by mining a highly exhaustive spatial dataset (the ORMEF database) that records Lessepsian fish invasion chronologically over the past 100 years compiled by D'Amen & Azzurro (2020a), updated in 2021 and available at the SEANOE repository (Azzurro et al., 2021). We extracted from this database occurrence data of 22 Lessepsian species that have invaded the Mediterranean Sea (Streftaris and Zenetos, 2006; Katsanevakis et al., 2014) and for which more than 25 records were available: Alepes djedaba, Bregmaceros nectabanus, Callionymus filamentosus, Etrumeus golanii, Fistularia commersonii, Hemiramphus far, Lagocephalus guentheri, Lagocephalus sceleratus, Parupeneus forsskali, Plotosus lineatus, Pterois miles, Sargocentron rubrum, Saurida lessepsianus, Scarus ghobban, Scomberomorus commerson, Siganus luridus, Siganus rivulatus, Sphyraena chrysotaenia, Sphyraena flavicauda, Torquigener flavimaculosus, Upeneus moluccensis, and Upeneus pori. Records in the native range (see Supplementary Appendix 1 for a description of the native ranges’ extent) were obtained from the open source datasets OBIS (OBIS, 2021) and GBIF (GBIF.org, 2021). Records were checked for reliability. Those with incomplete or inaccurate coordinates (e.g. without decimals) and those lacking the date or source information or listing unlikely points (e.g. falling within incorrect political borders) were discarded. Species distributions were compared with the AquaMaps dataset (Kaschner et al., 2010) for general geographical reliability; records not matching the distribution maps were removed. These datasets were thinned to 1° resolution to match the environmental layer grids.
For each species we assumed a stable native distribution and set the decade of its initial spread to the invaded range as a baseline condition for calculating the temperature match with the Mediterranean in the following decades. If the spread began before 1955 (25 of the 3396 records in the Mediterranean date from before 1955), we considered the temperature layers of 1955–1964, which is the earliest decade for which data are available.
Climate data
We selected the climate database which provides the longest time-series for the Mediterranean water, i.e. the World Ocean Atlas 2018 (National Oceanic and Atmospheric Administration, NOAA; Boyer et al., 2018) at a resolution of 1° for all six available decades, from 1955 to 2017. We selected surface water temperature, because it affects key biological processes, it is believed to determine the geographical distribution of marine species, especially fish (Cheung et al., 2009, 2013; Dijkstra et al., 2018) and has already been shown to be pioneered by the dusky spinefoot (D'Amen and Azzurro, 2020b). We selected the statistical mean surface temperature, the average of all unflagged interpolated values in each 1° square, which contains at least one measurement for the relevant oceanographic variable. We did not use salinity data due to poor geographical coverage for some seasons/decades.
We downloaded five temperature metrics: mean annual temperature (AT), one of the most widely used temperature variables in studies of climatic suitability, and the mean seasonal temperature in winter, spring, summer, and autumn (Supplementary Appendix 2). As regards the four seasonal variables, these were used to account for the interannual temperature variability (ITV), because it characterizes the temperate Mediterranean climate compared with the tropical climates of the native ranges of Lessepsian species. Noteworthy, we couldn't use higher resolution temperature data. Indeed, these are available only for the last few decades (from the 1980s onwards in Copernicus Marine Service) and not for the entire study period of six decades.
Statistical analysis
Preliminary test: importance of the chosen temperature variables in the native ranges
First, we verified that the chosen variables were significant predictors of species distribution in the native range in the decade when they first spread to the invaded range. We trained a multivariate Random Forest model for each species and generated pseudo-absences from the occupied area in the same number as presences (as recommended for classification techniques, Barbet-Massin et al., 2012) using the caret library (Kuhn, 2008) in R 4.0.5 (R Core Team, 2021) for each decade, considering species distribution as stable. The importance of a variable is generated by permuting the variables over all trees and then calculating the out-of-bag (OOB) error for each tree before and after permutation. Variables with a greater difference in estimated error are considered more important. The results are reported in Supplementary Appendix 3.
Analysis of spatio-temporal temperature match variation
The degree of matching temperature conditions between the native and invaded ranges of each species was calculated using the package dsmextra (Bouchet et al., 2020), an R implementation of the Extrapolation Detection (ExDet) tool proposed by Mesgaran et al. (2014), which allows conducting quantitative, spatially explicit assessments of univariate and combinatorial extrapolation using Euclidean and Mahalanobis distances (Bouchet et al., 2020). ExDet values between 0 and 1 indicate temperature conditions analogous to the native range (temperature matches), whereas values >1 or <0 indicate novel temperature conditions, non-analogous to those experienced in the native range, which can cause model extrapolation. ExDet values <0 indicate out-of-range values for any given covariate (which in a predictive model would lead to “Univariate extrapolation”); values >1 indicate novel combinations of values encountered within the univariate range of the reference covariates (which in a predictive model would induce “Combinatorial extrapolation”). The unbounded Mahalanobis metric can quantify the magnitude of the novelty in non-analogous temperature conditions: the further below 0, the greater the Univariate novelty, the further above 1, the greater the Combinatorial novelty. We applied the ExDet tool to both annual temperature alone (AT) and to the four seasons together (ITV). For ITV we identified the season making the largest contributions to the ExDet values in case of a non-analogous ITV (the “most influential covariate”). For each species, we evaluated the degree of novel AT and ITV conditions in the Mediterranean in all six decades compared with the native range temperature in the years of the initial spread of the species to the invaded range. We also calculated the longitudinal native range extension and tested if it was correlated with the temporal trends in the ExDet values.
Testing the temperature match hypothesis: is the invaded range limited by non-analogous conditions?
To establish whether species distribution in the novel range was related to matching temperature conditions and to identify potential temperature-related limitations, we extracted the AT and ITV ExDet values, calculated for the entire Mediterranean surface as described above, at the site of each record in the decade containing the year when it was detected; for records not falling in the ExDet raster layer coverage (due to missing values in the native temperature layers) we assigned the values of the closest cell. For each species, we also identified the first records indicating the species expansion (hereafter “front records”) in the Mediterranean, separately for the northern and southern coasts of the basin (D'Amen and Azzurro, 2020a). These records allowed estimating the maximum distance reached along the Mediterranean coasts and the spread rate of each species (D'Amen and Azzurro, 2020a). Finally, we tested the species pioneering behaviour by comparing the ExDet values of the entire record set with those of the front records.
Results
Spatio-temporal temperature match dynamics in the invaded range
Analysis of the six decadal temperature layers in the Mediterranean Sea highlighted some degree of novelty in AT and ITV for 16 and 21 of the 22 species, respectively (see, e.g., Figure 1, Figure 2, and Supplementary Appendix 4 for all the species), which can cause model extrapolation. Differences were large among species, while within species, the degree of novelty of AT was not significantly different across decades (p = 0.102, Kruskal–Wallis test) (Figure 2), whereas the proportion of the range showing analogous ITV conditions exhibited significant differences (p = 0.008, Kruskal–Wallis test), with analogy increasing in the final decades. The spatial pattern of the temperature matches was consistent across species, since analogous temperatures (where present) were mostly detected in the Levantine sector, whereas there was a westward gradient of increasing novelty (see, e.g., Figure 1 and Supplementary Appendix 4 for all species).
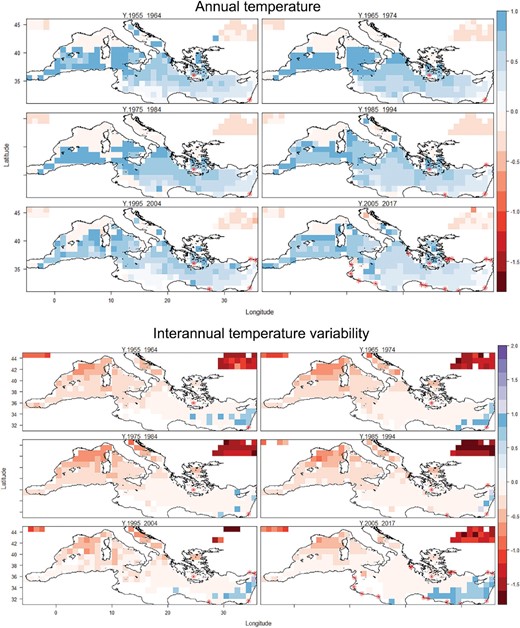
Representative temperature match estimates for a Lessepsian fish species, Alepes djedaba, across six decades. Annual temperature (AT, top) and interannual temperature variability (ITV bottom). Values from 0 to 1 indicate analogous conditions. Values <0 and >1 evidenced by vertical stripes indicate non-analogous conditions (univariate and combinatorial extrapolation, respectively). Red dots: cumulative occurrence records for each decade.
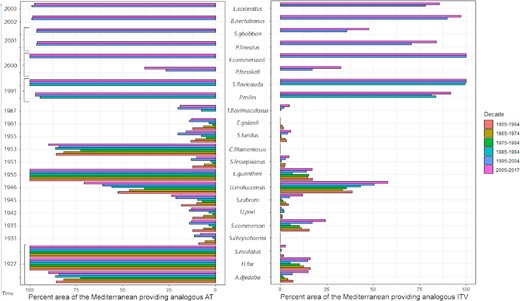
Percentage of the area of the Mediterranean basin providing temperature conditions (AT and ITV) analogous to the native ranges of the 22 fish considered. Species are listed according to time of arrival in the basin, and on the Y axis, date of the first record in the Mediterranean is reported. ExDet values are provided only for the decades when the species were present in the Mediterranean.
For 12 of the 22 species (A. djedaba, H. far, S. rivulatus, C. filamentosus, L. guentheri, F. commersonii, P. miles, S. flavicauda, P. lineatus, S. ghobban, B. nectabanus, and L. sceleratus), the AT at the time of arrival in the Mediterranean already matched the one in their native range. For these species, matching ATs were seen in about 80% of the Mediterranean Sea and this percentage has remained unchanged (or has slightly increased) up to the present (Figure 2, Supplementary Appendix 1). On the contrary, at the time of arrival U. moluccensis and P. forsskali experienced analogous conditions over 30% and 50% of the Mediterranean Sea, respectively; for U. moluccensis the area then increased up to 70%. For the other species, only a small portion of the Mediterranean (1–30%) offered matching temperature conditions; these areas were largely concentrated in the easternmost sectors of the basin.
As regards the ITV, the evidence was mixed. Six species (F. commersonii, P. miles, S. flavicauda, P. lineatus, B. nectabanus, and L. sceleratus) experienced matching ITVs in about 80% of the Mediterranean Sea. Notably, all six species entered the Mediterranean after 1990. In contrast, a small area of the Mediterranean (<10%) provided analogous ITV conditions for A. djedaba, S.rivulatus, S. chrysotaenia, U. pori, C. filamentosus, S. luridus, and E. golanii. Winter was the season providing non-analogous ITV conditions for nine species.
In the Levantine sector, 64% and 55% of species encountered matching AT and ITV conditions, respectively, at the time of their arrival. Data analysis showed that the proportion of Mediterranean Sea surface providing a temperature match with the native ranges at the time of species arrival was not related (p<>0.05i>, Spearman correlation test) to the size of the area that was eventually invaded (expressed as the maximum distance reached along the northern or the southern coasts). By contrast, it was significantly related (p<0.001, Spearman correlation test) to the species spread rate. Accordingly, species finding analogous temperature conditions at the time of their arrival in the Mediterranean spread faster, especially along the northern coast (p<<0.05i>, Wilcoxon test).
Testing the temperature match hypothesis: do the temperature conditions of the invaded range match those of the native range?
We attributed the ExDet values, calculated for AT or ITV, to each record reported in the Mediterranean according to the year of detection. There were large differences between species in the proportion of the records found in analogous AT/ITV conditions (Figure 3). Notably, the species that entered the Mediterranean after 1990 (late invaders) were largely recorded in analogous conditions, whereas the earlier invaders were often recorded at locations with non-matching temperature (Figure 3). For example, more than 75% of the records of S. luridus, E. golani, and S. chrysotaenia are at sites with non-analogous AT conditions. As regards the ITV, more than 75% of the records are at locations with non-matching conditions for nine species; notably, for two of them, S. chrysotaenia and C. filamentosus, none of the records are at sites with analogous conditions (Figure 3). This result cannot be attributed to differences in the native range longitudinal extent (as proxy of larger temperature tolerance) between early and late invaders (Wilcoxon test, p<>0.05i>).
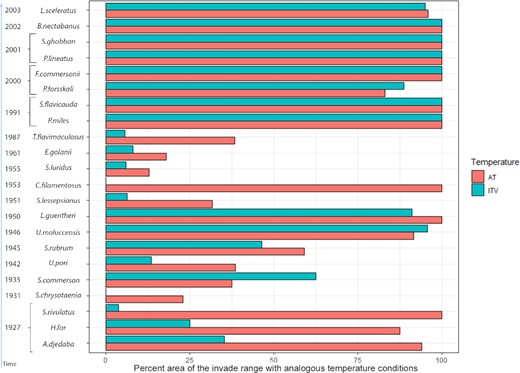
Percentage occurrence of the records of 22 Lessepsian fish species found in analogous AT/ITV conditions in the Mediterranean. Species are listed according to time of arrival in the basin, and on the Y axis date of the first record in the Mediterranean is reported.
The front records were detected at sites with non-analogous temperature conditions more often than were the other records. The difference was significant for both AT and ITV (p<0.0001, χ2 test). For most species, the ExDet values calculated for AT and ITV significantly correlated with record longitude (Supplementary Appendix 5).
For several species, the percentage of Mediterranean Sea surface providing analogous conditions often differed depending on whether AT or ITV was considered and was always greater for AT, especially for A. djedaba, H. far, S. rivulatus, S. chrysotaenia, L. guentheri, C. filamentosus, and S. ghobban (Figure 2). For such species, the winter temperature is the key determinant in the native range and the most influential covariate in determining non-analogous ITV conditions in the new range. As regards the ExDet values at the record locations (Figure 3), AT was always related to more extensive areas of analogous conditions than ITV, the sole exception being L. guentheri.
Discussion
We exploited the unique case study provided by the invasion of Lessepsian fish species into the Mediterranean Sea to analyse the invasion process according to the climate matching approach in a set of 22 marine fish for which abundant records are available, focusing on temperature variables. To the best of our knowledge, this is the first attempt to test the approach in the marine environment. Our findings highlight a widely different ability of the 22 species to pioneer novel temperature conditions, either annual or seasonal, as well as a very different establishment success before and after 1990.
Spatio-temporal temperature match dynamics
In the invasion process of all 22 species, the degree of temperature match showed a clear longitudinal gradient that was especially sharp when the spread was to the west of the basin. Given the tropical origin of Lessepsian fish and the colder water temperature in the western sectors of the Mediterranean, these results were largely expected (Pastor et al., 2018). At the time of their arrival in the eastern part of the basin, the temperature conditions in this area were similar to those of the native range of more than half of the species assessed in the study. In fact, the easternmost sector of the Mediterranean, which hosts the richest and most abundant set of Lessepsian species, is very different climatically from the central and western sectors (Pastor et al., 2018), so much so that a novel biogeographical name, the Lessepsian Province (Por, 1978), has been proposed for it.
Climate change will continue to affect the oceans. In the Mediterranean, seawater temperature has already changed over the past six decades, especially since the mid-1970s, with a rapid and continuous warming of surface waters (Volosciuk et al., 2016), with a significant seasonal warming trend varying in the different sub-basins (Shaltout and Omstedt, 2014). Our results are in line with such climatic data. In fact, they document a shrinking temperature gap, especially for the interannual temperature variability (representing seasonality), between the native and novel ranges of the 22 Lessepsian species in the last six decades.
The gap narrowed further around the 1990s. As a result, the Lessepsian fish species reaching the Mediterranean since then, encountered analogous temperatures (both AT and ITV) in a significantly higher proportion of the basin than did the earlier immigrants, records of which have often been reported in non-analogous temperature (especially ITV) conditions. This result cannot be attributed to differences in the native range longitudinal extent (as proxy of larger temperature tolerance) between early and late invaders, supporting the idea of a key role of rising temperatures in accelerating the Lessepsian phenomenon. These conclusions would reinforce the findings of earlier studies, which associated the increasing temperatures match between native and invaded ranges to the sharp increase and remarkable success of Lessepsian migrants since the 1990 (Vargas-Yanez et al., 2008; Raitsos et al., 2010). Future studies could further investigate the causes behind the increasing success of Lessepsian fish after this date and explore other possible differences between pre- and post-1990 introductions, according to the most important traits influencing their success probabilities, particularly ecological traits, environmental affinity (Belmaker et al., 2013; Arndt and Schembri, 2015), and morphological distinctness within the receiving community (Azzurro et al., 2014).
Testing the temperature match hypothesis
According to our data, Lessepsian fish often occupy areas characterized by non-matching temperature conditions, which entails a spread beyond their native temperature ranges. Such expansion was observed for species that migrated before 1990, particularly in leading-edge populations. In fact, the distributional range edges of the species are highly dynamic, thus they may be not representative of the environmental tolerance of the whole species (Jiménez-Valverde et al., 2011; Raybaud et al., 2015). Our findings seem to depict an invasion process shared by the non-indigenous species, where the individuals at the dispersion front encounter non-analogous temperatures and the rest of the population resides in core areas where the temperature conditions match those of the native range. For instance, the records of H. far show a westward pattern of analogy–novelty in seasonal temperature, where leading edge individuals moving towards the Strait of Sicily and the Aegean Sea found novel ITV conditions (Supplementary Appendix 4).
Similar pioneering events have been described in studies of non-indigenous plants, but only in 10% of the cases of niche dynamics (Atwater et al., 2018). Our results show that pioneering events along the temperature axes are common in Lessepsian fish and are not limited to the distributional range edge. For example, most of the invaded range of S. luridus and E. golani involves novel AT and ITV conditions. Notably, both species have spread beyond the Strait of Sicily, and individuals of E. golani have been recorded as far west as the Strait of Gibraltar (Tamsouri et al., 2019). Compared with the results reported by Parravicini et al. (2015) and D'Amen and Azzurro (2020b)—which are based on species niches measured according to a set of eight variables—we identified the areas lacking a temperature match both spatially and temporally and analysed their changes over time.
Several studies have attempted to explain the tremendous success of Lessepsian fish. Their observed ability to pioneer novel temperatures in recipient communities can be considered among the key factors, along with a lower biotic resistance (Belmaker et al., 2013; Azzurro et al., 2014). A rapid adaptive evolution (Bernardi et al., 2016), could also lead to novel physiological abilities and new possibilities to conquer sites providing non-analogous conditions. Gewing et al. (2019) have recently described the development of a broader temperature range resistance in non-indigenous ascidians over several decades since its entry into the Mediterranean. Similarly, the tropical seagrass Halophila stipulacea has shifted up its thermal tolerance in the Mediterranean (Wesselmann et al., 2020).
Potential limitations of our study relate to the use of a temperature range based solely on the realization of the temperature niche in the geographical space, whereas the physiological range may be different (e.g. Kellermann et al., 2012; Azzurro et al., 2014). To this end dynamic energy budgets (DEB) models, including physiological processes of individual organisms, could prove a useful tool for including this component in the assessment of functional environmental conditions (i.e. water temperature in this case) effects on the distribution and abundance these organisms (Sousa et al., 2010; Kearney and Porter, 2020). Other limitations of our study can be attributed to the coarse resolution of the data. Introduced species, including Lessepsian fish (Azzurro et al., 2016) can be subjected to variable time lags before the onset of population outbreaks, and the spatio-temporal nature of these invasions could be influenced by environmental and habitat changes, including water quality (Daskalov et al., 2007; Rilov and Galil, 2009). Including additional climate and ecological variables in future studies, would probably improve the understanding of the limiting factors shaping the distribution of these species, such as hydrological regimes, habitat suitability, shelter availability, and abundance of competitors and/or prey (Lodge, 1993; Moyle and Light, 1996; Vila-Gispert et al., 2005). Finally, the occurrence of a certain species in a certain place (presence-only records) does not necessarily entail that the area provides optimal conditions or that it host a well-established population. Probably, not all the observations included in this study can be attributed to the existence of locally established populations. Indeed, a few of them could be represented by records of vagrant individuals, or they can be related to non-sustaining sink subpopulations (Howe et al., 1991) that persist in non-analogous locations if there is a source (perhaps in analogous conditions) that provides sufficient individuals for regular “reinforcement”.
In the last decade, several studies have highlighted the importance of climate warming in making the Mediterranean a more suitable environment for Lessepsian fish (Raitsos et al., 2010). However, according to our data, the spatio-temporal nature of Lessepsian fish invasions cannot be explained only by temperature conditions, and other important factors such as life-history traits e.g. frequent reproduction, large body size and a long life span, the ability to exploit opportunistically available food resources intrinsic population dynamics and climatic/environmental changes in the recipient region are certainly worth to be considered (Vilizzi et al., 2021).
Identifying the main climatic novelties found by Lessepsian fish entering the Mediterranean has important implications for species distribution models (e.g. Albouy et al., 2014; Hattab et al., 2014) and can help to understand the cases in which bioclimatic models are informative and those where some degree of extrapolation may determine inaccuracies (Mahony et al., 2017). In the case of our 22 fish species, we show that even though the seasonal distribution of the covariates may explain species distribution in the native range, their use in projecting the invaded range can produce biased results in the Mediterranean range for most of the species recorded in the Mediterranean before 1990 because of non-analogue conditions between the donor and recipient regions. By contrast, since matching mean AT was found for a larger number of species, this seems to be a more useful variable for fitting species distribution models without causing biases in the projections. One way to reduce the uncertainty and improve predictions may be to train the models by pooling both native and invaded range data, where available (D'Amen & Azzurro, 2020a, b). In fact, the information carried by records from the invaded range can expand the modelled niche toward the fundamental niche and avoid extrapolation (Jiménez-Valverde et al., 2011).
For non-indigenous species, climate match assessment is used primarily to estimate the overall invasion risk and to prioritize management actions (Baker and Bomford, 2009). Our findings illustrate the idiosyncratic nature of each invasion and provide mixed evidence about the role of analogous temperatures in limiting the distribution of Lessepsian fish in the Mediterranean Sea. Despite seawater temperature is a major factor influencing the settlement of these tropical species (Ben-Tuvia, 1966; Ben Rais Lasram et al., 2008), our results indicate that many of them have overcome native temperature limits in their invaded range. In particular, although low winter seawater temperatures are held to be key factors limiting their survival and reproductive success of invasive Indo-Pacific fish (Rilov and Galil, 2009; Zurel et al., 2012), we found that in the Mediterranean some species (e.g. A. djedaba, H. far, S. rivulatus, and S. chrysotaenia) can tolerate lower winter temperatures. Altogether, our findings highlight the risk that assessment based only on temperature match criteria may underestimate the invasion potential of Lessepsian fish (Ben Rais Lasram et al., 2008; Ricciardi et al., 2013). This is a major challenge for managing risks from biological invasions in the marine realm. Including additional informative variables that do not cause potential extrapolation could improve our predictive ability, using species Distribution Models. For instance, salinity proven to be a key predictor for most of the considered species (D'Amen, Smeraldo and Azzurro, in prep.); thus, its inclusion could improve the prediction of suitable areas for the potential expansion of Lessepsian specie. On the other hand, often temperature is one of the few variables available, thus complementary approaches to the climate-matching one could be implemented to estimate the risk of invasion and establishment of non-indigenous species. For instance, a general and alternative approach recently proposed bases on measuring the environmental resistance of the landscape: it uses gradients of biotic similarity among native communities in the invaded region to predict the most likely routes of spread and has been shown to provide for predicting invasion risk that can be rapidly deployed (Lovell et al., 2021).
Data availability statement
The data underlying this article are available in the SEANOE Repository, at https://doi.org/10.17882/84182.
Author contributions statement
MD and EA contributed to the conception and design of the study. EA and SS organized the database. MD performed the statistical analysis and wrote the first draft of the manuscript. MD, ADF, SS, and EA contributed to manuscript revision and approved the submitted version.
Conflicts of interests
The authors have no conflicts of interest to declare.
Funding
This work was supported by the Interreg Med Programme [Grant number: 5216 | 5MED18_3.2_M23_007], 85% co-funded by the European Regional Development Fund.

{kind=link}
{kind=link}
{kind=link}