





Abstract
European hake (Merluccius merluccius) were caught alive in the northern Bay of Biscay in June/July 2002, using a codend specially designed to avoid crushing fish and also to retain water while hauling the trawl. In all, 1307 fish were tagged with anchor T-bar tags, injected with tetracycline, then released. The length range of the tagged and marked fish varied from 13 to 58 cm, and the modal size was 28 cm, at which length they were assumed to be 2–3 years old. A mean survival rate of 68.2% was estimated. Mortality was mainly caused by stress of capture and physical damage, and depended on the size of the catch and the depth of trawling. By the end of March 2003, 32 fish and three tags had been returned to the laboratory (a 2.7% return rate), the recapture data indicating that the probability of survival after release does not depend on catch depth or depth at location of release. For combined sexes, the somatic growth rate was estimated at 0.033 ± 0.019 cm day−1 (n=15). Males and females did not differ significantly in somatic growth rate, which were, respectively, 0.028±0.018 (n=6) and 0.033 ± 0.012 cm day−1 (n=6). This pilot experiment represents the first recorded mass tagging of European hake, which is known to be a very fragile species. The preliminary results indicate that it would be possible to carry out a large-scale tagging experiment of the European stocks in order to improve assessment and subsequent management decisions.
Introduction
European hake (Merluccius merluccius) are distributed widely over the northeast Atlantic shelf, from Norway to Mauritania, with density highest from the British Isles to the south of Spain (Casey and Pereiro, 1995). For several decades, the species has been targeted by the demersal fisheries of the region. It is landed as target or incidental catch by a variety of gear, including bottom trawl, net, and longline.
For all stocks assessed by the International Council for the Exploration of the Sea (ICES), total landings of European hake declined from about 120 000 t in the early 1960s to 80 000 t in the 1970s and 1980s. Catches declined further during the 1990s to the current level of 50 000 t (ICES, 2003). In recent years, assessments have raised concern about the state of the stocks. Spawning stock biomass is low and declining, and recruitment is poor, both phenomena being linked to overfishing of the population (ICES, 2003).
The stock assessment model currently used by ICES has proved to be useful in establishing diagnostics on the status of stocks. However, the approach has several limitations and shortcomings. Little is known about the growth of European hake, and no validation of age estimates used to build the catch-at-age matrix has yet been carried out. Compared with other species and stocks assessed by ICES, this is a concern, because hake otolith reading is particularly difficult. Despite considerable progress made in recent years, thanks in part to international otolith reading workshops (Piñeiro, 2000; Piñeiro et al., 2000), determining the age of hake older than 5 years is still problematic. The uncertainty about growth of larger fish leads ICES to use a plus-group at age 8, which is a source of bias in the assessment (ICES, 2003). An arbitrary constant-by-age value of 0.2 is used for natural mortality (M). Although there is no evidence for the existence of multiple populations, two stocks are considered for assessment purposes and analysed separately. The so-called “northern stock” is distributed from the northern British Isles to south of the Bay of Biscay, excluding the Spanish shelf (ICES Divisions IIIa, Sub-areas II, IV, VI, and VII, and Divisions VIIIa, b, d). The “southern stock” is distributed along the Spanish and Portuguese coasts (ICES Divisions VIIIc and IXa). No spatial structure is incorporated into the population dynamics model used to assess the stocks. For a stock that is distributed widely and exploited by numerous fleets fishing in different areas and catching different sizes of fish, such simplification of the dynamics could be a source of further uncertainty.
Although fishery data are essential for monitoring a fishery, they provide little biological information related to stock structure, movement, growth, or mortality of the fish, all of which are important in modelling stock dynamics. Furthermore, catch data alone are not sufficient to estimate stock abundance reliably, because estimated fishing mortality (F) and stock abundance have a large negative correlation (Fournier and Archibald, 1982; Deriso et al., 1985). Additional information is therefore vital. ICES already uses abundance indices from survey data. Tagging data could provide information on growth, movement, and mortality. Complex integrated stock assessment models can then employ this information (Deriso et al., 1985; Schnute and Richards, 1995). Before proceeding with a large-scale tagging experiment capable of providing such data for hake, we chose to conduct a pilot study. Our objectives were: to assess the feasibility of tagging hake, a fish reputed to be very fragile; to adapt and improve tagging techniques (gear used to catch live fish, fish handling during tagging and release); and to estimate the total number of tagged fish required in a large-scale experiment. To date, few attempts have been made to tag hake and, as a consequence, published results from hake-tagging experiments are scarce. In all European hake-tagging experiments of which we are aware (Belloc, 1935; Lucio et al., 2000), the number of tagged fish and recorded recaptures has been very small, insufficient to plan a large-scale experiment with confidence.
Star Oddi (Iceland) has recently developed a remotely controlled system for underwater tagging that can be used in a fishing trawl (US Patent 6286460). Although potentially of use for tagging hake, this new technique remains costly and requires further developments to tag fish calcified structures, such as otoliths. As an alternative, in line with what was used by Belloc (1935), Maucorps and Lefranc (1973), and Robles et al. (1975), we developed gear designed to reduce stress and physical damage during capture. In this paper, we present the methodology developed, a description of the tagging operations conducted during the pilot experiment, and preliminary results obtained so far.
Methods
The experiment was carried out in the northern Bay of Biscay, from 24 June to 10 July 2002, on board the 25 m trawler RV “Gwen Drez”. The pilot study was divided into two phases. The first was devoted to the trawl test and observation, and used two different video systems. The towed video system EROC (Engin Remorqué d'Observation des Chaluts) provided real-time observations of trawl behaviour, and the VECOC (Vidéo Embarquée de Contrôle et d'Observation des Chaluts) video system, mounted on the trawl, was used to record images inside the codend.
For the experiment, we used a 21.2 m bottom trawl equipped with rockhopper gear and a codend specially designed to avoid crushing fish and to retain water in the codend during hauling (Figure 1a). The tow periods were kept short (10–15 min) in order to limit the volume of the total catches, reduce captures of incidental species such as horse mackerel (Trachurus trachurus), and minimize the time spent by fish in the net. The trawl was hauled in slowly at an average speed of 0.5 m s−1. Attempts were made to haul gear with decompression stops, but this method did not improve fish condition.

(a) View of the codend, (b) puncturing the swimbladder, (c) the tagging process, and (d) tagged hake in a cool tank awaiting release.
Once aboard, fish were quickly transferred into a fibreglass tank continuously supplied with ambient seawater and drained with an external standpipe.
As suggested by Belloc (1935) and demonstrated by application to other species (Kohler, 1971; Keniry et al., 1996), to improve hake survival it is essential to perforate the swimbladder quickly. Once the excess gas has been vented, hake rapidly recover their natural position and are able to swim normally. The preferred zone to puncture the hake swimbladder is behind the anus in a region that does not contain essential organs. The puncture was made underwater using a hypodermic needle Terumo® 1.1 × 25 mm (Figure 1b). Fish were then kept in the holding tank for a minimum of 10 min in order to allow them to recover before further processing.
Fish were measured (total length) to the centimetre below. A T-bar tag (Floy Tag® FD-94 Anchor tag) was inserted at the base and in front of the second dorsal fin (Figure 1c). Each fish was injected with Terramycine®, a veterinary solution of oxytetracycline (OTC), at a dose of 60 mg per kg of fish. This antibiotic deposits a fluorescent mark on growing calcified structures, visible under UV light. Such marking has been used classically for age-validation studies (see Wright et al., 2002). The individual OTC dose was adjusted using the weight–length relationship W=5.13×10−6L3.0744 (Dorel, 1986). Fish dead, damaged, or weak were discarded.
After tagging, fish were held in a closed-circuit waiting tank, supplied with a continuous flow of cooled water (11–13°C; Figure 1d). Twice a day, fish were released away from the fishing zones where commercial trawlers were operating at the same time as the research vessel. Fish were released through a chute made from a PVC pipe passing through a scupper, and ending just beneath the sea surface.
For 15 days before the cruise, a large publicity effort was made (posters were mailed to fish wholesalers and Spanish scientists and displayed in auction rooms along the French Atlantic coast). In addition, several press releases informed the public that the operation could not succeed with the cruise alone, and that the return of tagged fish would be needed. A reward of 50 euros was offered for each fish returned to Ifremer laboratories.
Returned fish were stored frozen before subsequent processing. At the laboratory, thawed fish were measured to the centimetre below and weighed to the nearest decigram. For non-eviscerated fish, sex was determined.
Results
In order to ensure optimal conditions for observing trawl behaviour, nine tows (2 days) were conducted on sandy substrata in the Bay of Audierne (Figure 2a). Video observation confirmed proper functioning of the codend (Figure 3). Owing to the discharge current created by the codend's waterproof underside, fish stayed in front of the codend until it closed when hauling the trawl. A number of species were caught alive and in very good condition, so affirming the efficiency of the technique. Those species included whiting, sardine, sea bream, gurnards, dory, sprat, cephalopods, and jellyfish.
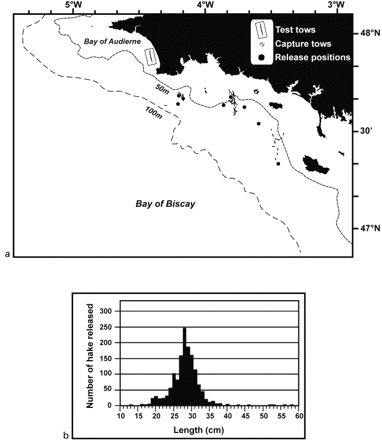
(a) Localities of test tows, capture tows, and release positions, and (b) frequency distribution of the size of fish released.
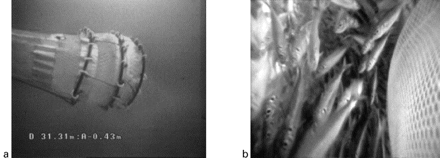
Behaviour of the codend underwater; (a) general view, EROC image, and (b) view inside the codend, VECOC image.
Following the initial test tows, 83 tows (10 days) in four different areas (Figure 2a) were devoted to hake capture. Tow areas were SW Glénans, SE Glénans, NW Groix, and NW Belle Ile, at depths varying from 35 to 99 m.
At the end of the cruise, a total of 1307 fish had been tagged, marked, and released at nine locations (Figure 2a). The size range of the tagged and marked fish varied from 13 to 58 cm (Figure 2b), with a modal size of 28 cm, corresponding to a fish age of 2–3 years, according to current age determination criteria.
A mean survival rate of 68.2±14.5% after capture and tagging was estimated from 11 tows. The mortality appeared principally to be a consequence of the capture process (average contribution of 78% to the total mortality, vs. 22% ascribable to the tagging process). A survival test was also carried out using a cooled tank: a survival rate of 63% was observed 39 h after tagging. The variability of the mortality was relatively high, but it appeared to depend on two main factors: size of by-catch, principally horse mackerel, and trawling depth (Figure 4). A regression analysis indicated that 52% of the variation in rate of mortality was explained by the trawling depth (r2=0.525, F=9.95, p=0.12). Although contributing to the survival rate, the trawling depth should not be too limiting if fish density is sufficient.
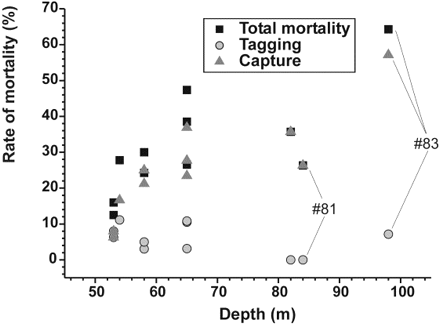
Relative contributions of capture and tagging to the immediate mortality with respect to depth of trawling. Relative contributions were measured on 11 tows. For tow #81, at 84 m depth, the total mortality rate was 26%, and was entirely due to capture. For tow #83 at 98 m depth, the mortality rate was 64%, split between 57% after capture and 7% after subsequent tagging.
By the end of March 2003, 32 tagged fish and 3 individual tags had been returned with information on capture (date, geographical position, fishing gear, fisher's name), i.e. a recapture rate of 2.7%. Table 1 lists the details of the fish recaptured.
Tag–recapture data. F, M, and U denote sex (female, male, undetermined). Gonads were immature or inactive. Distance has been calculated using the orthodromic formula between release and recapture localities.
| Characteristics at capture | Characteristics at recapture | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Fish | Date | Capture depth (m) | TL (cm) | Latitude (N) | Longitude (W) | Release depth (m) | Date | Latitude(N) | Longitude(W) | Days of freedom | Distance (nautical miles) | TL (cm) | Sex |
| 56 | 27/06/2002 | 82–85 | 31 | 47°37′ | 3°42′ | 67 | 09/09/2002 | 47°32.611′ | 3°56.312′ | 74 | 10.6 | 33 | F |
| 141 | 28/06/2002 | 35–41 | 33 | 47°37′ | 3°42′ | 65 | 16/07/2002 | 47°41.421′ | 3°47.499′ | 18 | 5.8 | 33 | F |
| 172 | 28/06/2002 | 36–40 | 38 | 47°37′ | 3°42′ | 67 | 05/07/2002 | 47°36.100′ | 3°49.250′ | 7 | 5.0 | 37 | U |
| 182 | 28/06/2002 | 36–40 | 33 | 47°37′ | 3°42′ | 67 | 24/01/2003 | 47°39.318′ | 3°20.132′ | 210 | 14.9 | No fish | |
| 275 | 29/06/2002 | 37–60 | 28 | 47°37′ | 3°41′ | 65 | 13/08/2002 | n/a | n/a | 45 | n/a | 29 | M |
| 307 | 02/07/2002 | 46–54 | 35 | 47°41′ | 3°48′ | 56 | 08/08/2002 | 47°29.880′ | 4°14.223′ | 37 | 20.9 | 34 | F |
| 354 | 02/07/2002 | 48–63 | 27 | 47°41′ | 3°48′ | 54 | 10/09/2002 | 47°31.000′ | 3°57.000′ | 70 | 11.7 | 28 | M |
| 408 | 03/07/2002 | 59–63 | 21 | 47°38′ | 3°42′ | 65 | 09/03/2003 | 47°05.000′ | 4°32.000′ | 249 | 47.3 | 37 | M |
| 452 | 04/07/2002 | 50–80 | 29 | 47°20′ | 3°27′ | 83 | 20/11/2002 | 45°58.000′ | 1°36.000′ | 139 | 111.9 | 35 | F |
| 519 | 04/07/2002 | 66–83 | 29 | 47°32′ | 3°35′ | 48 | 13/07/2002 | 47°35.032′ | 3°36.390′ | 9 | 3.2 | 29 | F |
| 594 | 05/07/2002 | 51–74 | 28 | 47°38′ | 3°42′ | 66 | 16/08/2002 | n/a | n/a | 42 | n/a | 29 | F |
| 747 | 06/07/2002 | 55–78 | 28 | 47°38′ | 3°42′ | 66 | 08/07/2002 | 47°41.421′ | 3°47.499′ | 2 | 5.0 | 28 | U |
| 762 | 06/07/2002 | 55–78 | 40 | 47°38′ | 3°42′ | 66 | 12/07/2002 | 47°28.170′ | 3°37.520′ | 6 | 10.3 | 39 | F |
| 802 | 06/07/2002 | 63–74 | 35 | 47°38′ | 3°42′ | 65 | 20/08/2002 | 47°22.600′ | 3°33.600′ | 45 | 16.4 | 32 | F |
| 809 | 06/07/2002 | 63–74 | 26 | 47°38′ | 3°42′ | 65 | 11/09/2002 | 47°32.000′ | 3°57.000′ | 67 | 11.8 | 27 | U |
| 855 | 06/07/2002 | 63–74 | 33 | 47°38′ | 3°42′ | 65 | 08/08/2002 | 47°34.000′ | 4°01.000′ | 33 | 13.4 | 32 | F |
| 859 | 06/07/2002 | 63–74 | 30 | 47°38′ | 3°42′ | 65 | 09/07/2002 | 47°41.421′ | 3°47.499′ | 3 | 5.0 | 30 | F |
| 863 | 06/07/2002 | 63–74 | 29 | 47°38′ | 3°42′ | 65 | 08/07/2002 | 47°41.421′ | 3°47.499′ | 2 | 5.0 | 29 | F |
| 880 | 06/07/2002 | 63–74 | 29 | 47°38′ | 3°42′ | 65 | 15/10/2002 | 47°31.300′ | 3°54.000′ | 101 | 10.5 | 34 | F |
| 928 | 08/07/2002 | 48–66 | 24 | 47°38′ | 3°51′ | 62 | 19/09/2002 | 47°27.284′ | 3°56.026′ | 73 | 11.2 | 26 | M |
| 968 | 08/07/2002 | 48–66 | 30 | 47°38′ | 3°51′ | 62 | 19/07/2002 | 47°26.540′ | 4°04.990′ | 11 | 14.8 | 30 | M |
| 989 | 08/07/2002 | 48–68 | 31 | 47°38′ | 3°51′ | 62 | 03/09/2002 | 47°37.120′ | 3°58.510′ | 57 | 5.1 | 32 | F |
| 1021 | 08/07/2002 | 80–99 | 32 | 47°38′ | 4°12′ | 97 | 09/07/2002 | 47°37.600′ | 4°11.800′ | 1 | 0.4 | 31 | F |
| 1032 | 08/07/2002 | 80–99 | 32 | 47°38′ | 4°12′ | 97 | 30/07/2002 | 47°20.000′ | 3°50.000′ | 22 | 23.3 | 30 | M |
| 1038 | 08/07/2002 | 80–99 | 24 | 47°38′ | 4°12′ | 97 | 30/07/2002 | 47°31.500′ | 3°59.000′ | 22 | 10.9 | 24 | M |
| 1125 | 09/07/2002 | 50–58 | 31 | 47°38′ | 3°52′ | 62 | 05/09/2002 | 47°32.915′ | 4°05.042′ | 58 | 10.2 | 32 | M |
| 1180 | 09/07/2002 | 50–58 | 25 | 47°38′ | 3°52′ | 62 | 05/09/2002 | 47°33.000′ | 4°59.000′ | 58 | 45.5 | No fish | |
| 1181 | 09/07/2002 | 50–58 | 26 | 47°38′ | 3°52′ | 62 | 01/10/2002 | 47°29.031′ | 3°33.305′ | 84 | 15.5 | 29 | F |
| 1191 | 09/07/2002 | 50–58 | 23 | 47°38′ | 3°52′ | 62 | 09/09/2002 | 47°31.719′ | 3°56.601′ | 62 | 7.0 | 28 | U |
| 1196 | 09/07/2002 | 50–58 | 25 | 47°38′ | 3°52′ | 62 | 08/08/2002 | 47°43.090′ | 3°46.300′ | 30 | 6.4 | 25 | M |
| 1210 | 09/07/2002 | 50–58 | 29 | 47°38′ | 3°52′ | 62 | 07/08/2002 | 47°34.000′ | 3°59.000′ | 29 | 6.2 | 30 | U |
| 1217 | 09/07/2002 | 50–58 | 29 | 47°38′ | 3°52′ | 62 | 27/08/2002 | 47°36.000′ | 4°08.000′ | 49 | 11.0 | 29 | F |
| 1300 | 09/07/2002 | 58–67 | 30 | 47°38′ | 3°52′ | 64 | 15/07/2002 | 47°41.421′ | 3°47.499′ | 6 | 4.6 | 30 | F |
| 1310 | 09/07/2002 | 58–67 | 25 | 47°38′ | 3°52′ | 64 | 02/08/2002 | 47°28.000′ | 3°55.000′ | 24 | 10.2 | No fish | |
| 1359 | 09/07/2002 | 58–67 | 40 | 47°38′ | 3°52′ | 64 | 21/08/2002 | 47°20.430′ | 3°46.850′ | 43 | 17.9 | 41 | M |
| Characteristics at capture | Characteristics at recapture | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Fish | Date | Capture depth (m) | TL (cm) | Latitude (N) | Longitude (W) | Release depth (m) | Date | Latitude(N) | Longitude(W) | Days of freedom | Distance (nautical miles) | TL (cm) | Sex |
| 56 | 27/06/2002 | 82–85 | 31 | 47°37′ | 3°42′ | 67 | 09/09/2002 | 47°32.611′ | 3°56.312′ | 74 | 10.6 | 33 | F |
| 141 | 28/06/2002 | 35–41 | 33 | 47°37′ | 3°42′ | 65 | 16/07/2002 | 47°41.421′ | 3°47.499′ | 18 | 5.8 | 33 | F |
| 172 | 28/06/2002 | 36–40 | 38 | 47°37′ | 3°42′ | 67 | 05/07/2002 | 47°36.100′ | 3°49.250′ | 7 | 5.0 | 37 | U |
| 182 | 28/06/2002 | 36–40 | 33 | 47°37′ | 3°42′ | 67 | 24/01/2003 | 47°39.318′ | 3°20.132′ | 210 | 14.9 | No fish | |
| 275 | 29/06/2002 | 37–60 | 28 | 47°37′ | 3°41′ | 65 | 13/08/2002 | n/a | n/a | 45 | n/a | 29 | M |
| 307 | 02/07/2002 | 46–54 | 35 | 47°41′ | 3°48′ | 56 | 08/08/2002 | 47°29.880′ | 4°14.223′ | 37 | 20.9 | 34 | F |
| 354 | 02/07/2002 | 48–63 | 27 | 47°41′ | 3°48′ | 54 | 10/09/2002 | 47°31.000′ | 3°57.000′ | 70 | 11.7 | 28 | M |
| 408 | 03/07/2002 | 59–63 | 21 | 47°38′ | 3°42′ | 65 | 09/03/2003 | 47°05.000′ | 4°32.000′ | 249 | 47.3 | 37 | M |
| 452 | 04/07/2002 | 50–80 | 29 | 47°20′ | 3°27′ | 83 | 20/11/2002 | 45°58.000′ | 1°36.000′ | 139 | 111.9 | 35 | F |
| 519 | 04/07/2002 | 66–83 | 29 | 47°32′ | 3°35′ | 48 | 13/07/2002 | 47°35.032′ | 3°36.390′ | 9 | 3.2 | 29 | F |
| 594 | 05/07/2002 | 51–74 | 28 | 47°38′ | 3°42′ | 66 | 16/08/2002 | n/a | n/a | 42 | n/a | 29 | F |
| 747 | 06/07/2002 | 55–78 | 28 | 47°38′ | 3°42′ | 66 | 08/07/2002 | 47°41.421′ | 3°47.499′ | 2 | 5.0 | 28 | U |
| 762 | 06/07/2002 | 55–78 | 40 | 47°38′ | 3°42′ | 66 | 12/07/2002 | 47°28.170′ | 3°37.520′ | 6 | 10.3 | 39 | F |
| 802 | 06/07/2002 | 63–74 | 35 | 47°38′ | 3°42′ | 65 | 20/08/2002 | 47°22.600′ | 3°33.600′ | 45 | 16.4 | 32 | F |
| 809 | 06/07/2002 | 63–74 | 26 | 47°38′ | 3°42′ | 65 | 11/09/2002 | 47°32.000′ | 3°57.000′ | 67 | 11.8 | 27 | U |
| 855 | 06/07/2002 | 63–74 | 33 | 47°38′ | 3°42′ | 65 | 08/08/2002 | 47°34.000′ | 4°01.000′ | 33 | 13.4 | 32 | F |
| 859 | 06/07/2002 | 63–74 | 30 | 47°38′ | 3°42′ | 65 | 09/07/2002 | 47°41.421′ | 3°47.499′ | 3 | 5.0 | 30 | F |
| 863 | 06/07/2002 | 63–74 | 29 | 47°38′ | 3°42′ | 65 | 08/07/2002 | 47°41.421′ | 3°47.499′ | 2 | 5.0 | 29 | F |
| 880 | 06/07/2002 | 63–74 | 29 | 47°38′ | 3°42′ | 65 | 15/10/2002 | 47°31.300′ | 3°54.000′ | 101 | 10.5 | 34 | F |
| 928 | 08/07/2002 | 48–66 | 24 | 47°38′ | 3°51′ | 62 | 19/09/2002 | 47°27.284′ | 3°56.026′ | 73 | 11.2 | 26 | M |
| 968 | 08/07/2002 | 48–66 | 30 | 47°38′ | 3°51′ | 62 | 19/07/2002 | 47°26.540′ | 4°04.990′ | 11 | 14.8 | 30 | M |
| 989 | 08/07/2002 | 48–68 | 31 | 47°38′ | 3°51′ | 62 | 03/09/2002 | 47°37.120′ | 3°58.510′ | 57 | 5.1 | 32 | F |
| 1021 | 08/07/2002 | 80–99 | 32 | 47°38′ | 4°12′ | 97 | 09/07/2002 | 47°37.600′ | 4°11.800′ | 1 | 0.4 | 31 | F |
| 1032 | 08/07/2002 | 80–99 | 32 | 47°38′ | 4°12′ | 97 | 30/07/2002 | 47°20.000′ | 3°50.000′ | 22 | 23.3 | 30 | M |
| 1038 | 08/07/2002 | 80–99 | 24 | 47°38′ | 4°12′ | 97 | 30/07/2002 | 47°31.500′ | 3°59.000′ | 22 | 10.9 | 24 | M |
| 1125 | 09/07/2002 | 50–58 | 31 | 47°38′ | 3°52′ | 62 | 05/09/2002 | 47°32.915′ | 4°05.042′ | 58 | 10.2 | 32 | M |
| 1180 | 09/07/2002 | 50–58 | 25 | 47°38′ | 3°52′ | 62 | 05/09/2002 | 47°33.000′ | 4°59.000′ | 58 | 45.5 | No fish | |
| 1181 | 09/07/2002 | 50–58 | 26 | 47°38′ | 3°52′ | 62 | 01/10/2002 | 47°29.031′ | 3°33.305′ | 84 | 15.5 | 29 | F |
| 1191 | 09/07/2002 | 50–58 | 23 | 47°38′ | 3°52′ | 62 | 09/09/2002 | 47°31.719′ | 3°56.601′ | 62 | 7.0 | 28 | U |
| 1196 | 09/07/2002 | 50–58 | 25 | 47°38′ | 3°52′ | 62 | 08/08/2002 | 47°43.090′ | 3°46.300′ | 30 | 6.4 | 25 | M |
| 1210 | 09/07/2002 | 50–58 | 29 | 47°38′ | 3°52′ | 62 | 07/08/2002 | 47°34.000′ | 3°59.000′ | 29 | 6.2 | 30 | U |
| 1217 | 09/07/2002 | 50–58 | 29 | 47°38′ | 3°52′ | 62 | 27/08/2002 | 47°36.000′ | 4°08.000′ | 49 | 11.0 | 29 | F |
| 1300 | 09/07/2002 | 58–67 | 30 | 47°38′ | 3°52′ | 64 | 15/07/2002 | 47°41.421′ | 3°47.499′ | 6 | 4.6 | 30 | F |
| 1310 | 09/07/2002 | 58–67 | 25 | 47°38′ | 3°52′ | 64 | 02/08/2002 | 47°28.000′ | 3°55.000′ | 24 | 10.2 | No fish | |
| 1359 | 09/07/2002 | 58–67 | 40 | 47°38′ | 3°52′ | 64 | 21/08/2002 | 47°20.430′ | 3°46.850′ | 43 | 17.9 | 41 | M |
Tag–recapture data. F, M, and U denote sex (female, male, undetermined). Gonads were immature or inactive. Distance has been calculated using the orthodromic formula between release and recapture localities.
| Characteristics at capture | Characteristics at recapture | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Fish | Date | Capture depth (m) | TL (cm) | Latitude (N) | Longitude (W) | Release depth (m) | Date | Latitude(N) | Longitude(W) | Days of freedom | Distance (nautical miles) | TL (cm) | Sex |
| 56 | 27/06/2002 | 82–85 | 31 | 47°37′ | 3°42′ | 67 | 09/09/2002 | 47°32.611′ | 3°56.312′ | 74 | 10.6 | 33 | F |
| 141 | 28/06/2002 | 35–41 | 33 | 47°37′ | 3°42′ | 65 | 16/07/2002 | 47°41.421′ | 3°47.499′ | 18 | 5.8 | 33 | F |
| 172 | 28/06/2002 | 36–40 | 38 | 47°37′ | 3°42′ | 67 | 05/07/2002 | 47°36.100′ | 3°49.250′ | 7 | 5.0 | 37 | U |
| 182 | 28/06/2002 | 36–40 | 33 | 47°37′ | 3°42′ | 67 | 24/01/2003 | 47°39.318′ | 3°20.132′ | 210 | 14.9 | No fish | |
| 275 | 29/06/2002 | 37–60 | 28 | 47°37′ | 3°41′ | 65 | 13/08/2002 | n/a | n/a | 45 | n/a | 29 | M |
| 307 | 02/07/2002 | 46–54 | 35 | 47°41′ | 3°48′ | 56 | 08/08/2002 | 47°29.880′ | 4°14.223′ | 37 | 20.9 | 34 | F |
| 354 | 02/07/2002 | 48–63 | 27 | 47°41′ | 3°48′ | 54 | 10/09/2002 | 47°31.000′ | 3°57.000′ | 70 | 11.7 | 28 | M |
| 408 | 03/07/2002 | 59–63 | 21 | 47°38′ | 3°42′ | 65 | 09/03/2003 | 47°05.000′ | 4°32.000′ | 249 | 47.3 | 37 | M |
| 452 | 04/07/2002 | 50–80 | 29 | 47°20′ | 3°27′ | 83 | 20/11/2002 | 45°58.000′ | 1°36.000′ | 139 | 111.9 | 35 | F |
| 519 | 04/07/2002 | 66–83 | 29 | 47°32′ | 3°35′ | 48 | 13/07/2002 | 47°35.032′ | 3°36.390′ | 9 | 3.2 | 29 | F |
| 594 | 05/07/2002 | 51–74 | 28 | 47°38′ | 3°42′ | 66 | 16/08/2002 | n/a | n/a | 42 | n/a | 29 | F |
| 747 | 06/07/2002 | 55–78 | 28 | 47°38′ | 3°42′ | 66 | 08/07/2002 | 47°41.421′ | 3°47.499′ | 2 | 5.0 | 28 | U |
| 762 | 06/07/2002 | 55–78 | 40 | 47°38′ | 3°42′ | 66 | 12/07/2002 | 47°28.170′ | 3°37.520′ | 6 | 10.3 | 39 | F |
| 802 | 06/07/2002 | 63–74 | 35 | 47°38′ | 3°42′ | 65 | 20/08/2002 | 47°22.600′ | 3°33.600′ | 45 | 16.4 | 32 | F |
| 809 | 06/07/2002 | 63–74 | 26 | 47°38′ | 3°42′ | 65 | 11/09/2002 | 47°32.000′ | 3°57.000′ | 67 | 11.8 | 27 | U |
| 855 | 06/07/2002 | 63–74 | 33 | 47°38′ | 3°42′ | 65 | 08/08/2002 | 47°34.000′ | 4°01.000′ | 33 | 13.4 | 32 | F |
| 859 | 06/07/2002 | 63–74 | 30 | 47°38′ | 3°42′ | 65 | 09/07/2002 | 47°41.421′ | 3°47.499′ | 3 | 5.0 | 30 | F |
| 863 | 06/07/2002 | 63–74 | 29 | 47°38′ | 3°42′ | 65 | 08/07/2002 | 47°41.421′ | 3°47.499′ | 2 | 5.0 | 29 | F |
| 880 | 06/07/2002 | 63–74 | 29 | 47°38′ | 3°42′ | 65 | 15/10/2002 | 47°31.300′ | 3°54.000′ | 101 | 10.5 | 34 | F |
| 928 | 08/07/2002 | 48–66 | 24 | 47°38′ | 3°51′ | 62 | 19/09/2002 | 47°27.284′ | 3°56.026′ | 73 | 11.2 | 26 | M |
| 968 | 08/07/2002 | 48–66 | 30 | 47°38′ | 3°51′ | 62 | 19/07/2002 | 47°26.540′ | 4°04.990′ | 11 | 14.8 | 30 | M |
| 989 | 08/07/2002 | 48–68 | 31 | 47°38′ | 3°51′ | 62 | 03/09/2002 | 47°37.120′ | 3°58.510′ | 57 | 5.1 | 32 | F |
| 1021 | 08/07/2002 | 80–99 | 32 | 47°38′ | 4°12′ | 97 | 09/07/2002 | 47°37.600′ | 4°11.800′ | 1 | 0.4 | 31 | F |
| 1032 | 08/07/2002 | 80–99 | 32 | 47°38′ | 4°12′ | 97 | 30/07/2002 | 47°20.000′ | 3°50.000′ | 22 | 23.3 | 30 | M |
| 1038 | 08/07/2002 | 80–99 | 24 | 47°38′ | 4°12′ | 97 | 30/07/2002 | 47°31.500′ | 3°59.000′ | 22 | 10.9 | 24 | M |
| 1125 | 09/07/2002 | 50–58 | 31 | 47°38′ | 3°52′ | 62 | 05/09/2002 | 47°32.915′ | 4°05.042′ | 58 | 10.2 | 32 | M |
| 1180 | 09/07/2002 | 50–58 | 25 | 47°38′ | 3°52′ | 62 | 05/09/2002 | 47°33.000′ | 4°59.000′ | 58 | 45.5 | No fish | |
| 1181 | 09/07/2002 | 50–58 | 26 | 47°38′ | 3°52′ | 62 | 01/10/2002 | 47°29.031′ | 3°33.305′ | 84 | 15.5 | 29 | F |
| 1191 | 09/07/2002 | 50–58 | 23 | 47°38′ | 3°52′ | 62 | 09/09/2002 | 47°31.719′ | 3°56.601′ | 62 | 7.0 | 28 | U |
| 1196 | 09/07/2002 | 50–58 | 25 | 47°38′ | 3°52′ | 62 | 08/08/2002 | 47°43.090′ | 3°46.300′ | 30 | 6.4 | 25 | M |
| 1210 | 09/07/2002 | 50–58 | 29 | 47°38′ | 3°52′ | 62 | 07/08/2002 | 47°34.000′ | 3°59.000′ | 29 | 6.2 | 30 | U |
| 1217 | 09/07/2002 | 50–58 | 29 | 47°38′ | 3°52′ | 62 | 27/08/2002 | 47°36.000′ | 4°08.000′ | 49 | 11.0 | 29 | F |
| 1300 | 09/07/2002 | 58–67 | 30 | 47°38′ | 3°52′ | 64 | 15/07/2002 | 47°41.421′ | 3°47.499′ | 6 | 4.6 | 30 | F |
| 1310 | 09/07/2002 | 58–67 | 25 | 47°38′ | 3°52′ | 64 | 02/08/2002 | 47°28.000′ | 3°55.000′ | 24 | 10.2 | No fish | |
| 1359 | 09/07/2002 | 58–67 | 40 | 47°38′ | 3°52′ | 64 | 21/08/2002 | 47°20.430′ | 3°46.850′ | 43 | 17.9 | 41 | M |
| Characteristics at capture | Characteristics at recapture | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Fish | Date | Capture depth (m) | TL (cm) | Latitude (N) | Longitude (W) | Release depth (m) | Date | Latitude(N) | Longitude(W) | Days of freedom | Distance (nautical miles) | TL (cm) | Sex |
| 56 | 27/06/2002 | 82–85 | 31 | 47°37′ | 3°42′ | 67 | 09/09/2002 | 47°32.611′ | 3°56.312′ | 74 | 10.6 | 33 | F |
| 141 | 28/06/2002 | 35–41 | 33 | 47°37′ | 3°42′ | 65 | 16/07/2002 | 47°41.421′ | 3°47.499′ | 18 | 5.8 | 33 | F |
| 172 | 28/06/2002 | 36–40 | 38 | 47°37′ | 3°42′ | 67 | 05/07/2002 | 47°36.100′ | 3°49.250′ | 7 | 5.0 | 37 | U |
| 182 | 28/06/2002 | 36–40 | 33 | 47°37′ | 3°42′ | 67 | 24/01/2003 | 47°39.318′ | 3°20.132′ | 210 | 14.9 | No fish | |
| 275 | 29/06/2002 | 37–60 | 28 | 47°37′ | 3°41′ | 65 | 13/08/2002 | n/a | n/a | 45 | n/a | 29 | M |
| 307 | 02/07/2002 | 46–54 | 35 | 47°41′ | 3°48′ | 56 | 08/08/2002 | 47°29.880′ | 4°14.223′ | 37 | 20.9 | 34 | F |
| 354 | 02/07/2002 | 48–63 | 27 | 47°41′ | 3°48′ | 54 | 10/09/2002 | 47°31.000′ | 3°57.000′ | 70 | 11.7 | 28 | M |
| 408 | 03/07/2002 | 59–63 | 21 | 47°38′ | 3°42′ | 65 | 09/03/2003 | 47°05.000′ | 4°32.000′ | 249 | 47.3 | 37 | M |
| 452 | 04/07/2002 | 50–80 | 29 | 47°20′ | 3°27′ | 83 | 20/11/2002 | 45°58.000′ | 1°36.000′ | 139 | 111.9 | 35 | F |
| 519 | 04/07/2002 | 66–83 | 29 | 47°32′ | 3°35′ | 48 | 13/07/2002 | 47°35.032′ | 3°36.390′ | 9 | 3.2 | 29 | F |
| 594 | 05/07/2002 | 51–74 | 28 | 47°38′ | 3°42′ | 66 | 16/08/2002 | n/a | n/a | 42 | n/a | 29 | F |
| 747 | 06/07/2002 | 55–78 | 28 | 47°38′ | 3°42′ | 66 | 08/07/2002 | 47°41.421′ | 3°47.499′ | 2 | 5.0 | 28 | U |
| 762 | 06/07/2002 | 55–78 | 40 | 47°38′ | 3°42′ | 66 | 12/07/2002 | 47°28.170′ | 3°37.520′ | 6 | 10.3 | 39 | F |
| 802 | 06/07/2002 | 63–74 | 35 | 47°38′ | 3°42′ | 65 | 20/08/2002 | 47°22.600′ | 3°33.600′ | 45 | 16.4 | 32 | F |
| 809 | 06/07/2002 | 63–74 | 26 | 47°38′ | 3°42′ | 65 | 11/09/2002 | 47°32.000′ | 3°57.000′ | 67 | 11.8 | 27 | U |
| 855 | 06/07/2002 | 63–74 | 33 | 47°38′ | 3°42′ | 65 | 08/08/2002 | 47°34.000′ | 4°01.000′ | 33 | 13.4 | 32 | F |
| 859 | 06/07/2002 | 63–74 | 30 | 47°38′ | 3°42′ | 65 | 09/07/2002 | 47°41.421′ | 3°47.499′ | 3 | 5.0 | 30 | F |
| 863 | 06/07/2002 | 63–74 | 29 | 47°38′ | 3°42′ | 65 | 08/07/2002 | 47°41.421′ | 3°47.499′ | 2 | 5.0 | 29 | F |
| 880 | 06/07/2002 | 63–74 | 29 | 47°38′ | 3°42′ | 65 | 15/10/2002 | 47°31.300′ | 3°54.000′ | 101 | 10.5 | 34 | F |
| 928 | 08/07/2002 | 48–66 | 24 | 47°38′ | 3°51′ | 62 | 19/09/2002 | 47°27.284′ | 3°56.026′ | 73 | 11.2 | 26 | M |
| 968 | 08/07/2002 | 48–66 | 30 | 47°38′ | 3°51′ | 62 | 19/07/2002 | 47°26.540′ | 4°04.990′ | 11 | 14.8 | 30 | M |
| 989 | 08/07/2002 | 48–68 | 31 | 47°38′ | 3°51′ | 62 | 03/09/2002 | 47°37.120′ | 3°58.510′ | 57 | 5.1 | 32 | F |
| 1021 | 08/07/2002 | 80–99 | 32 | 47°38′ | 4°12′ | 97 | 09/07/2002 | 47°37.600′ | 4°11.800′ | 1 | 0.4 | 31 | F |
| 1032 | 08/07/2002 | 80–99 | 32 | 47°38′ | 4°12′ | 97 | 30/07/2002 | 47°20.000′ | 3°50.000′ | 22 | 23.3 | 30 | M |
| 1038 | 08/07/2002 | 80–99 | 24 | 47°38′ | 4°12′ | 97 | 30/07/2002 | 47°31.500′ | 3°59.000′ | 22 | 10.9 | 24 | M |
| 1125 | 09/07/2002 | 50–58 | 31 | 47°38′ | 3°52′ | 62 | 05/09/2002 | 47°32.915′ | 4°05.042′ | 58 | 10.2 | 32 | M |
| 1180 | 09/07/2002 | 50–58 | 25 | 47°38′ | 3°52′ | 62 | 05/09/2002 | 47°33.000′ | 4°59.000′ | 58 | 45.5 | No fish | |
| 1181 | 09/07/2002 | 50–58 | 26 | 47°38′ | 3°52′ | 62 | 01/10/2002 | 47°29.031′ | 3°33.305′ | 84 | 15.5 | 29 | F |
| 1191 | 09/07/2002 | 50–58 | 23 | 47°38′ | 3°52′ | 62 | 09/09/2002 | 47°31.719′ | 3°56.601′ | 62 | 7.0 | 28 | U |
| 1196 | 09/07/2002 | 50–58 | 25 | 47°38′ | 3°52′ | 62 | 08/08/2002 | 47°43.090′ | 3°46.300′ | 30 | 6.4 | 25 | M |
| 1210 | 09/07/2002 | 50–58 | 29 | 47°38′ | 3°52′ | 62 | 07/08/2002 | 47°34.000′ | 3°59.000′ | 29 | 6.2 | 30 | U |
| 1217 | 09/07/2002 | 50–58 | 29 | 47°38′ | 3°52′ | 62 | 27/08/2002 | 47°36.000′ | 4°08.000′ | 49 | 11.0 | 29 | F |
| 1300 | 09/07/2002 | 58–67 | 30 | 47°38′ | 3°52′ | 64 | 15/07/2002 | 47°41.421′ | 3°47.499′ | 6 | 4.6 | 30 | F |
| 1310 | 09/07/2002 | 58–67 | 25 | 47°38′ | 3°52′ | 64 | 02/08/2002 | 47°28.000′ | 3°55.000′ | 24 | 10.2 | No fish | |
| 1359 | 09/07/2002 | 58–67 | 40 | 47°38′ | 3°52′ | 64 | 21/08/2002 | 47°20.430′ | 3°46.850′ | 43 | 17.9 | 41 | M |
The recapture positions indicate that fish movements were variable in both direction and distance (Figure 5a). Time at liberty varied from 1 to 249 days (Figure 5b). There is a positive correlation between time at liberty and the distance between location of release and recapture (r=0.520; p=0.001), although individual movement strategies may differ significantly. For instance, fish #452 “covered” 112 nautical miles during 139 days at liberty, whereas fish #408 was recaptured less than 50 miles from its release point after 249 days at liberty.
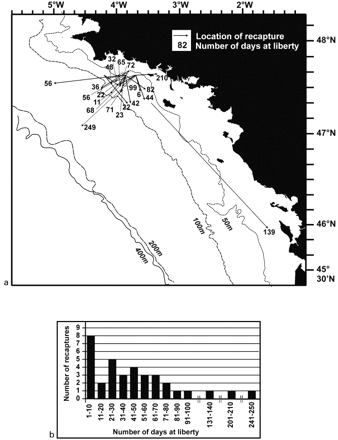
(a) Localities of recapture of tagged hake (only fish that had covered a distance greater than 10 nautical miles are shown), and (b) frequency distribution of time at liberty.
Recapture data indicate that the probability of survival after release does not depend on catch depth or depth at location of release, which were similar, because fish were caught and released in close proximity (Figure 2a). This is clearly shown in Table 2, which compares the frequency distributions of tagged and recaptured fish with respect to catch depth.
Frequency distribution of hake tagged and recaptured with respect to depth at capture.
| Depth at capture (m) | Number of fish tagged | % | Number of fish recaptured | % |
|---|---|---|---|---|
| 40–49 | 81 | 6.2 | 3 | 8.6 |
| 50–59 | 192 | 14.7 | 8 | 22.9 |
| 60–69 | 428 | 32.7 | 9 | 25.7 |
| 70–79 | 307 | 23.5 | 9 | 25.7 |
| 80–89 | 259 | 19.8 | 3 | 8.6 |
| 90–99 | 40 | 3.1 | 3 | 8.6 |
| Total | 1307 | 100 | 35 | 100 |
| Depth at capture (m) | Number of fish tagged | % | Number of fish recaptured | % |
|---|---|---|---|---|
| 40–49 | 81 | 6.2 | 3 | 8.6 |
| 50–59 | 192 | 14.7 | 8 | 22.9 |
| 60–69 | 428 | 32.7 | 9 | 25.7 |
| 70–79 | 307 | 23.5 | 9 | 25.7 |
| 80–89 | 259 | 19.8 | 3 | 8.6 |
| 90–99 | 40 | 3.1 | 3 | 8.6 |
| Total | 1307 | 100 | 35 | 100 |
Frequency distribution of hake tagged and recaptured with respect to depth at capture.
| Depth at capture (m) | Number of fish tagged | % | Number of fish recaptured | % |
|---|---|---|---|---|
| 40–49 | 81 | 6.2 | 3 | 8.6 |
| 50–59 | 192 | 14.7 | 8 | 22.9 |
| 60–69 | 428 | 32.7 | 9 | 25.7 |
| 70–79 | 307 | 23.5 | 9 | 25.7 |
| 80–89 | 259 | 19.8 | 3 | 8.6 |
| 90–99 | 40 | 3.1 | 3 | 8.6 |
| Total | 1307 | 100 | 35 | 100 |
| Depth at capture (m) | Number of fish tagged | % | Number of fish recaptured | % |
|---|---|---|---|---|
| 40–49 | 81 | 6.2 | 3 | 8.6 |
| 50–59 | 192 | 14.7 | 8 | 22.9 |
| 60–69 | 428 | 32.7 | 9 | 25.7 |
| 70–79 | 307 | 23.5 | 9 | 25.7 |
| 80–89 | 259 | 19.8 | 3 | 8.6 |
| 90–99 | 40 | 3.1 | 3 | 8.6 |
| Total | 1307 | 100 | 35 | 100 |
Regarding somatic growth, fish having spent less than 40 days at liberty had a nil or even negative somatic growth rate. Growth rates were therefore estimated only from fish for which total length differed by at least 1 cm between release and recapture. For combined sexes, the growth rate was estimated at 0.033 ± 0.019 cm day−1(mean ± 1 s.d.; n=15). The growth rates of 0.028±0.018 (n=6) and 0.033 ± 0.012 cm day−1 (n=6) for males and females, respectively, did not differ significantly (t-test, p=0.605).
Discussion
Morales-Nin (1997) wrote that “European hake rearing and capture-marking experiments are not readily applied due to the species biology and bathymetric distribution”. However, an early marking experiment did succeed with the recovery of one tagged fish after 255 days at liberty (Belloc, 1935). A more recent experiment concluded that trawling, as usually carried out, was not ideal for capturing live fish (Lucio et al., 2000). Instead, those authors used longliners working shallower than 100 m. They tagged and released 151 fish (size range, 21–70 cm) during 21 days at sea. Three recaptures were made and one fish was returned (after 24 days of freedom). One fish spent 195 days at liberty (P. Lucio, pers. comm.). Compared with that experiment, our methodology greatly increases efficiency and reduces the cost of tagging studies. The recapture rate and maximum time at liberty are slightly higher (2.7% vs. 1.9%, 249 days vs. 195 days, respectively), but, more importantly, the number of fish tagged per day was greater by a factor approaching 20. There is, however, still room for improvement through better targeting of high density areas, training of tagging personnel (336 fish were tagged during the last day of the cruise), and reduced mortality through capture and fish handling, by refining the methodology.
The recapture success clearly indicates that the method is applicable at depths up to 100 m, although the depth at which fish were captured appears to contribute to the probability of survival at capture. This result agrees with similar experiments on other species (Gitschlag and Renaud, 1994; Keniry et al., 1996; Nostvik and Pedersen, 1999). It is also a key point in developing a large-scale tagging experiment, because in most areas of interest (Celtic Sea, Iberian coast), the working depths would likely be higher than in the northern Bay of Biscay, where hake are found relatively shallow (40–60 m).
Despite the existence of a large literature base on the growth of European hake in different parts of the European Atlantic (Hickling, 1933; Belloc, 1935; Guichet et al., 1973; Decamps and Labastie, 1978; Iglesias and Dery, 1981; Goñi and Pinñeiro, 1988; Guichet, 1988; Guichet and Labastie, 1992), there is still uncertainty about interpreting the differences observed in growth data and fitted growth models. Are the differences real and linked, for instance, to environmental conditions and/or area effects on growth, or do they depend on inconsistencies in age estimation? Although to date our data are limited, they clearly show that we could address uncertainty in growth rate through adequate mark–recapture experiments. Only one recaptured specimen was badly damaged (on the caudal fin). Negative growth rates observed for some fish having spent less than 40 days at liberty may be the result of shrinkage following freezing, as observed for other species (Armstrong and Stewart, 1997; Al Hassan et al., 1999), and/or are more likely due to insufficient precision of measurement at sea (to the centimetre below). Nevertheless, fish that spend sufficient time at liberty give invaluable information on growth rate. The only direct measurement reported in the literature is that of Belloc (1935), who observed a growth rate of 0.046 cm day−1 during 255 days for a fish (sex unreported) that measured 30 cm at release. This result falls in the range of our own estimate, corroborating Belloc's conclusion that “hake is a fast growing species”. However, the great variability of the individual growth rates in this study indicates that this conclusion needs to be refined with further data. Sexual dimorphism in growth is well known in hake, and would likely appear from the third year on, in association with earlier onset of maturity in males (Hickling, 1933). This feature could not be observed in the present work because recoveries were estimated to be of fish 2–3 years old.
Recoveries are also insufficient to draw any conclusion on stock structure or movements of hake in the Bay of Biscay. However, it was not our intention to consider this point in the pilot study. Nevertheless, although most tagged fish were recaptured in the vicinity of their release location, the distance travelled by one fish was relatively great, suggesting that some exchange at a population scale is possible.
Only a large-scale tagging experiment conducted over a larger area would permit a better understanding of the structure of the population and quantify movements between areas. Several questions, which have implications for stock assessment, still need to be clarified: (1) What is the relationship between the main nursery areas (Celtic Sea, Grande Vasière) and the rest of the population? (2) What are the mixing rates between the so-called “northern” and “southern” stocks? (3) What are the levels of mixing within and between each fishing area? Several approaches may be used to analyse the data collected. The simplest is, as in the present paper, to draw arrows from release to recovery location and to plot distance travelled against time at liberty (Schaefer et al., 1961). Such an analysis, however, does not take into account the spatio-temporal distribution of fishing effort, which obviously affects the number of recoveries. To address this, several statistical methods incorporating fishing effort for estimating movement rates from tag recoveries have been developed (Ishii, 1979; Sibert, 1984; Hilborn, 1990; Kleiber and Hampton, 1994; Sibert et al., 1999).
Another aim of the present study was to help evaluate the number of tags required in a large-scale experiment to achieve the objectives of estimating growth, movement, and rate of mortality. From our results, it appears that, by improving hake survival during and just after tagging, and by increasing the efficiency of public information, we could reasonably expect a return rate of about 5%. When compared with other tagging studies with similar objectives (Kearney and Gillett, 1983; Kleiber et al., 1987; Hampton, 1997, 2000; Kaltongga, 1998; Sibert et al., 1999), returns of the order of 3000 would be a reasonable target. This would require a total release number of about 60 000 tagged hake. It would, however, be better to conduct simulations in order to better quantify the effect of the number of tagged fish releases on the precision of the parameters estimated from the tagging data collected (Bertignac, 1996).
The preliminary information on growth will be refined through rigorous analysis of otolith growth of recaptured fish, in order to evaluate the relevance of this experiment in terms of age validation. Whatever those results, the major outcome of the present study is that we should be able to get better information on hake biology and population dynamics using this method of capture and tagging. The methodology may need to be refined, but for the first time, it would be possible to carry out a large-scale tagging experiment covering the stocks exploited and managed at a European level, in order to improve assessment and subsequent management decisions.
More generally, the field of application of the new codend (patent taken out at INPI – no. 0212496) should exceed the framework of hake tagging, because it permits the capture of live fish and invertebrates in good condition.
We dedicate this work to Dr Hervé Troadec✠, with whom we thought this project could be done. We thank André Ogor (IFREMER, Lasaa, Brest), Jean Philippe Vacherot (IFREMER, TMSI, Lorient) and Jacques Labastie (IFREMER, RH, L'Houmeau), for their technical support in the project and their involvement during the cruise. The success of the cruise is largely attributable to the efficiency of the captain and crew of the “Gwen Drez”. We also thank the Spanish scientists who participated in the publicity campaign, Dr Louis Quiniou (UBO, Brest), for his help in sex determination, and Rebecca Shuford (University of South Carolina, USA), and Aisling Lannin (University of Cork, Ireland), for improving the English of the manuscript. Thanks are also due to Dave Japp (Fisheries and Oceanographic Support Services cc., Cape Town, South Africa) and an anonymous referee, for their detailed and accurate comments on this paper.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}