





Abstract
The red imported fire ant, Solenopsis invicta (Buren), is an invasive species in the southern United States and is expanding its range westward to California and eastward up the Atlantic Coast. This voracious predator can reach extremely high densities and have widespread effects once it invades an ecosystem. We conducted a 2-yr sampling study and a series of greenhouse and field experiments to document the impact of red imported fire ants on beneficial insects in cotton. We found that the densities of 12 of 13 natural enemies sampled on cotton plants in 1999, and 8 out of 8 sampled in 2000, were negatively correlated with the densities of foraging fire ant workers. Red imported fire ants reduced the survival of lady beetles (Coccinella septempunctata L., and Hippodamia convergens Guérin-Méneville) (Coleoptera: Coccinellidae) by 50% and green lacewing larvae (Chrysoperla carnea Stephens) (Neuroptera: Chrysopidae) by 38% in greenhouse experiments. Fire ants did not, however, reduce the survival of spiders (Oxyopidae, Thomisidae, and Clubionidae). We used a commercially available fire ant bait to suppress fire ant populations in cotton fields during the 2000 growing season and compared the densities of beneficial arthropods in treated versus control fields. Densities of lady beetles, spiders, and big-eyed bugs (Heteroptera: Geocoridae) were significantly higher in fields with suppressed fire ant populations than in fields with relatively large fire ant populations. The effect of fire ants on minute pirate bugs (Heteroptera: Anthocoridae) was inconsistent, and populations of damsel bugs (Heteroptera: Nabidae) and hooded beetles (Coleoptera: Anthicidae) were not affected by fire ant suppression. The results of this study suggest that red imported fire ants are major intraguild predators of important beneficial arthropods in cotton.
Intraguild predation occurs when potential competitors engage in predator-prey interactions (Polis et al. 1989, Polis and Holt 1992). Although once thought to be relatively rare, recent studies suggest that intraguild predation is common in terrestrial food webs and can be intense (Polis et al. 1989; Spiller and Schoener 1990,1994,1996; Hurd and Eisenberg 1990; Rosenheim et al. 1993,1995). For example, hemipteran predators (assassin bugs, Heteroptera: Reduviidae; damsel bugs, Heteroptera: Nabidae; and big-eyed bugs, Heteroptera: Geocoridae) kill up to 90% of green lacewings in California cotton fields (Rosenheim 2001). We investigated the potential for intraguild predation by an invasive ant on a suite of arthropod predators in an agricultural system. The goal of our study was to gain insight into factors that determine the intensity of intraguild predation and the effect of an invading intraguild predator.
The red imported fire ant, Solenopsis invicta Buren, was introduced into the southeastern United States ≈70 yr ago. After a fairly slow initial range expansion, this species spread across the southern tier of the United States to California and continues to expand its range northward along the East Coast (MacKay and Fagerlund 1997, Vinson 1997, Anonymous 1999). As well as being a direct health threat to humans and a nuisance pest of landscapes, these ants have pervasive ecological effects because of their voracious predatory habits and their ability to out-compete other surface-dwelling arthropods. They have been implicated in the decline of native invertebrates (Porter and Savignano 1990) and vertebrates (Allen et al. 1997) and may affect the survival and dispersal of native plants (Ready and Vinson 1995).
Red imported fire ants probably have pervasive effects in agroecosystems as well as natural ecosystems (Adams et al. 1976,1977,1988; Adams 1986; Morrison et al. 1997; Vinson 1997). Their biggest effect on agroecosystems may arise from their consumption of other arthropods. Red imported fire ants have been reported as significant predators of a wide variety of insect pests including boll weevil, Anthonomus grandis grandis Boheman (Sterling 1978, Jones and Sterling 1979, Sturm et al. 1990); sugarcane borer, Diatraea saccharialis (F.) (Negm and Hensley 1967; Reagan et al. 1972; Bessin and Reagan 1993); horn fly, Haematobia irritans (L.) (Lemke and Kissam 1988; Hu and Frank 1996); velvetbean caterpillar, Anticarsia gemmatalis Hübner (Lee et al. 1990), rednecked peanutworm, Stegasta bosqueella (Chambers) (Vogt et al. 2001), and even whiteflies (Homoptera: Aleyrodidae) in greenhouses (Morrill 1977). As a consequence, some growers within the southeastern United States regard fire ants as beneficial insects.
Other studies, however, raise questions about the beneficial effect of red imported fire ants in agroecosystems (Negm and Hensley 1967,1969; Sterling 1978; Sturm et al. 1990; Vinson 1994). Most notably, red imported fire ant workers are relatively indiscriminant, omnivorous predators that attack beneficial insects as well as pests (Wilson and Oliver 1969, Ricks and Vinson 1970, Morrill 1977, Lee et al. 1990, Vinson 1994). For example, red imported fire ant workers attack predators of horn flies and other pests in pastures (Hu and Frank 1996) and predators of aphids and scale insects in pecans (Bugg and Dutcher 1989, Tedders et al. 1990, Dutcher 1998). Red imported fire ants may also decrease the efficacy of parasitic wasps, primarily by consuming wasp larvae and pupae (Lopez 1982,Sturm et al. 1990, Vinson and Scarborough 1991).
Although several authors maintain that red imported fire ants are relatively benign, beneficial predators in cotton (Sterling et al. 1979; Reilly and Sterling 1983a,1983b), our experience in Alabama cotton fields suggests that fire ant workers routinely attack other predators and may interfere with biological control of cotton pests (Eubanks 2000,2001). To help elucidate the effects of red imported fire ants on insects in cotton and other agroecosystems, we conducted a series of field and greenhouse studies to quantify the effects of red imported fire ants on important beneficial arthropods commonly found in cotton. Understanding the effect of red imported fire ants on natural enemies is essential for the development of effective pest management strategies in areas already infested with fire ants and will help predict the effect of fire ants on agricultural and natural systems as this invasive species expands its range.
Materials and Methods
Sampling Data.
In 1999, three cotton fields at the E. V. Smith Substation of the Alabama Agricultural Experiment Station in Shorter were sampled throughout the growing season. The fields were ≈25 ha each and were separated by 2-5 km. Plants were sampled approximately every other week (May through August) with a 40-cm diam sweep net (12 samples per field, 25 sweeps per sample). Samples were returned to the laboratory, and arthropods were identified and counted. In 2000, we used a combination of beat cloth sampling and visual searches to sample cotton plants in four large fields at the substation. We used beat cloth sampling/visual searching to match the scouting protocols used for action thresholds and economic injury levels in southeastern cotton (Knutson and Ruberson 1996). We spread a 1-m2 cloth on the ground between two rows of cotton. Six cotton plants, three from each row, were vigorously shaken over the cloth. All insects that fell on the cloth were quickly identified and counted. We took 10 samples in each field on each sampling date (approximately weekly from May to August). Data from 1999 and 2000 were log (n + 1) transformed before analyses (Sokal and Rohlf 1995). We determined the correlation between fire ant density and the density of each natural enemy to estimate the effects of fire ants on beneficial arthropods. Separate analyses were performed for each year.
Greenhouse Experiment.
We quantified the survival of three of the most important natural enemies found in cotton in the presence or absence of foraging fire ant workers in a greenhouse experiment. We examined lady beetles (Coccinella septempunctata L., and Hippodamia convergens Guérin-Méneville), green lacewings (Chrysoperla carnea Stephens), and spiders (Araneae: Oxyopidae, Thomisidae, and Clubionidae) because of their importance as biological control agents, and because of their relative ease of collection or commercial availability, which facilitated replication of the experiment. Individual cotton plants (five-node size) were placed within cages (75 cm × 30 cm × 33 cm) constructed of fine mesh mosquito netting. Cages were attached to a 53 cm × 43 cm × 13 cm plastic pan with 0.5-cm diam plastic tubing. The plastic pan contained a small laboratory-maintained fire ant colony containing ≈500 fire ant workers. The lip of each pan was coated with liquid Teflon to prevent ants from escaping. We opened the plastic tubes to approximately half the cages. We conducted 29 replicates of lady beetle larvae without ants, 28 replicates of lady beetle larvae with ants, 34 replicates of lacewing larvae without ants, 30 replicates of lacewing larvae with ants, 30 replicates of spiders without ants and 25 replicates of spiders with ants during the late summer and early fall 2000.
Plants exposed to ant colonies were monitored to ensure that fire ant workers initiated foraging before the start of the experiment. We found that the average density of foraging workers on plants exposed to ants was 2.2 ± 1.8 individuals per visual search, well within the range observed in the field (see field results). Vinson and Scarborough (1989,1991) successfully used similar experimental protocols to document the effect of fire ants on the survival of other arthropods. At the start of the experiment, two lady beetles (one larva and one adult), two green lacewing larvae, or two spiders were placed in each cage. No food was provided for the predators.
Lady beetles and spiders were collected from the weedy edges of agricultural fields and fence lines. Green lacewing larvae and some adult lady beetles were obtained from a commercial supplier (Beneficial Insectary, Redding, CA) to supplement our field collections. Lady beetles were randomly selected for each replicate of the experiment to mimic the mix of C. septempunctata and H. convergens beetles found in cotton. Likewise, two adult spiders were randomly chosen from our collections to represent a mix of the common clubionid, oxyopid, and thomisid species abundant in cotton. The primary clubionid we used was the winter spider, Chiracanthium inclusum (Hentz). The oxyopid spiders included were the striped lynx spider, Oxyopes salticus Hentz, and the green lynx spider, Peucetia viridans (Hentz). The most common thomisid in cotton, and the species used in our experiment, was the celer crab spider, Misumenops celer (Hentz). All of these spiders have been documented as important predators of cotton insect pests (Breene et al. 1993).
Cages were monitored to ensure that the natural enemies initiated foraging on cotton plants after they were placed on the plant surface. After 24 h, we intensively searched the cotton plant, soil surface, and cage for living natural enemies. Missing natural enemies were considered killed by ants. Observations during this and preliminary experiments suggested that most insects killed by foraging fire ant workers were taken to the mound in the Teflon-lipped pan. We compared the survival number of lady beetles, lacewing larvae, and spiders in the presence and absence of ants using 2 × 3 contingency tables (0 predators alive, 1 predator alive, 2 predators alive) (Sokal and Rohlf 1995).
Field Experiment.
To experimentally estimate the effect of fire ants on beneficial arthropods in the field, we suppressed fire ant populations with a commercially available, ant-specific bait (Amdro, American Cyanamid, Wayne, NJ) and compared beneficial arthropod abundance in bait-treated cotton plots with their abundance in control plots during the 2000 growing season. Commercially available fire ant baits are excellent tools for this kind of experiment because they are composed of an inert, corn "grit" carrier impregnated with soybean oil that is very attractive to foraging fire ant workers but ignored by other insects. A toxicant, in this case hydramethylnon, is incorporated into the oil. Foraging workers find the bait and carry it back to the colony where the toxicant is spread throughout the mound, and all members of the colony are affected.
We used four large cotton fields (>20 ha each) at the E. V. Smith Substation in this experiment. We established two 1.2-ha plots at opposite ends of each field. Plots were separated by at least 100 m. The plots were randomly assigned to either the control or treatment group. Treated plots received application of fire ant bait (5.5 kg/ha) approximately 1 wk and 8 wk after planting. We sampled each field weekly throughout the growing season using the beat cloth sampling protocol described previously (10 samples per field, per week). Means of the 10 samples collected in each plot on each sampling date were log (n + 1) transformed. We used repeated measures analysis of variance (ANOVA), with field as a blocking factor, to compare the densities of beneficial arthropods among control and treated fields (SAS ProcMixed, Khattree and Naik 1999).
Results
Fire ants were very abundant at our field sites. Fire ant abundance ranged from 0 to 5.5 foraging workers per plant in 1999, and 0 to 17 foraging workers per plant in 2000. Fire ants were the third most abundant natural enemy on cotton plants in 1999 (lady beetles and big-eyed bugs were more abundant) and the most abundant natural enemy in 2000.
The abundances of 12 out of 13 natural enemy species in 1999, and 8 of 8 species in 2000, were negatively correlated with the abundance of fire ant workers on cotton plants (Table 1). These arthropods included the most abundant and economically important natural enemies in cotton such as lady beetles, green lacewings, and spiders. In general, estimated effects of fire ants on natural enemy abundances were lower in 2000 than in 1999 (Table 1). For example, in 2000, the coefficients estimating the correlation between fire ant abundance and the abundance of spiders, minute pirate bugs, Orius insidiosus (Say), and lacewings were all substantially lower than in 1999 (Table 1).
Pearson's product–moment correlation coefficients (r) and degrees of freedom (df) for correlations between red imported fire ant abundance and natural enemy abundance in cotton fields in 1999 and 2000
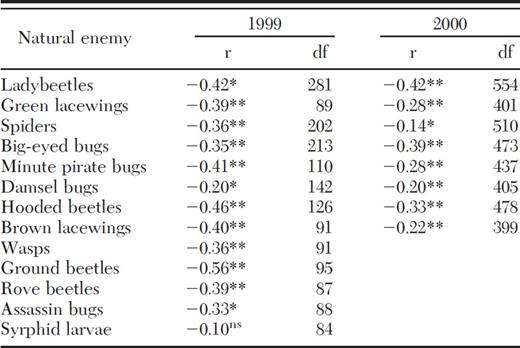
P < 0.05*, P < 0.01**.
Pearson's product–moment correlation coefficients (r) and degrees of freedom (df) for correlations between red imported fire ant abundance and natural enemy abundance in cotton fields in 1999 and 2000
P < 0.05*, P < 0.01**.
To a large extent, the results of our greenhouse experiment mirrored our field results. Fire ant workers inflicted significant mortality on lady beetles and green lacewing larvae (Fig. 1A and B). Lady beetles suffered the highest mortality from ant predation. Lady beetle survival was 50% lower in the presence of fire ants than in the absence of ants (Fig. 1A; G = 14.31, df = 2, P < 0.001). Green lacewing survival was 38% lower in the presence of fire ants than in ant-free controls (Fig. 1B; G = 6.1, df = 2, P < 0.05). Spider survival was unaffected by exposure to foraging fire ants (Fig. 1C; G = 0, df = 2, P = 1.00).
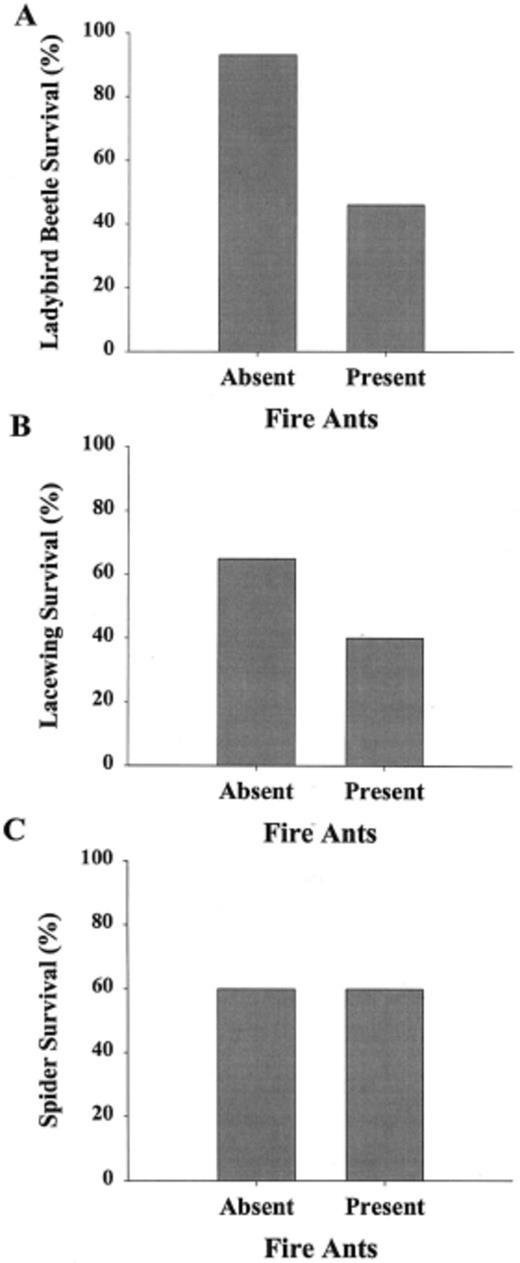
Percent survival of lady beetles (A), green lacew-ing larvae (B), and spiders (C) in the presence or absence of red imported fire ant workers on cotton plants in a greenhouse experiment. Survival of lady beetles and green lace-wing larvae were significantly lower (α = 0.05) in the presence of fire ants than in ant-free controls. Spider survival was unaffected by the presence of ants.
Baits were very effective at reducing fire ant populations during our field experiment. Fire ant populations were, on average, three times larger in untreated (control) plots than in treated plots (6.19 ± 1.25 ants per sample in control fields versus 2.03 ± 1.22 ants per sample in treated fields) (treatment: F1,3 = 64.94, P = 0.004). The effect of fire ant bait, however, was not consistent across fields and across dates (date × treatment interaction, F4,8 = 3.87, P = 0.046). In some fields, the effect was dramatic and persisted throughout the growing season. In other fields, the effect of the bait was relatively small, and fire ant suppression lasted only a few weeks. As a result, fire ant densities in bait-treated and control plots were approximately equal in some fields on some dates. To help ensure that we were testing for an effect of fire ant density on natural enemy abundance, we included a given field on a given sampling date in our analyses, if there was at least an 80% difference in fire ant abundance between the bait-treated and control plots in that field. There were not enough lacewings present in enough fields on enough dates to include them in the statistical analysis.
Four out of the six beneficial arthropods that we monitored responded to changes in fire ant density. Averaged across all dates, lady beetles were 2.9 times more abundant in plots of cotton with suppressed densities of fire ants than in control plots with relatively high densities of fire ants (Fig. 2A; Table 2). Likewise, spiders were 2.5 times more abundant in plots of cotton with suppressed fire ants (Fig. 2B; Table 2), and big-eyed bugs were 2.9 times more abundant (Fig. 2C; Table 2). Minute pirate bugs, however, were significantly more abundant in control plots than in bait-treated plots (Fig. 2D; Table 2). The abundance of damsel bugs (Fig. 2E; Table 2) and hooded beetles (Notoxus spp.) (Fig. 2F; Table 2) were unaffected by fire ant suppression.
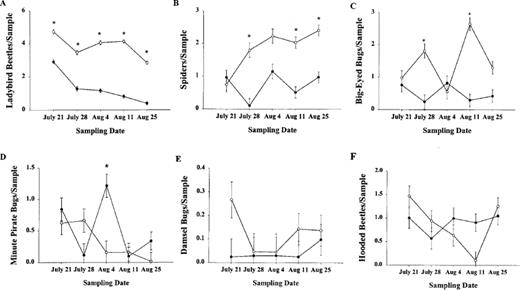
Mean (± SE) of lady beetles (A), spiders (B), big-eyed bugs (C) minute pirate bugs (D), damsel bugs (E), and hooded beetles (F) per plot in cotton fields where fire ants were suppressed or not suppressed during the 2000 growing season. An asterisk indicates that means were significantly different (α = 0.05) on that sampling date (Bonferroni test).
Results of ANOVA's examining the effects of treatment (fire ant suppression or not) on natural enemy abundance in cotton fields
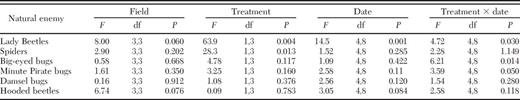
Results of ANOVA's examining the effects of treatment (fire ant suppression or not) on natural enemy abundance in cotton fields
Discussion
The results of this study suggest that intraguild predation in cotton involving red imported fire ants is pervasive and often intense. The density of fire ant workers was negatively correlated with almost all natural enemies sampled during 1999 and 2000, including lady beetles, green lacewings, and spiders (Table 1). In addition, foraging fire ant workers dramatically reduced the survival of lady beetles and green lacewings in greenhouse experiments (Figs. 1A and B). Most telling, populations of several important beneficial arthropods were two to three times larger in cotton plots with suppressed populations of fire ants (Fig. 2). Evidence of such pervasive intraguild predation by a single higher order predator is almost unprecedented.
Our greenhouse results suggesting that fire ants severely reduce the survival of lady beetles and lacewing larvae were not surprising given the strong negative correlation between fire ant and natural enemy abundance observed in the field. We were surprised, however, to find that spider survival was not affected by exposure to foraging fire ant workers in the greenhouse experiment (Fig. 1C). This was particularly unexpected given the strong effect of fire ant suppression on spider abundance in the field experiment (Fig. 2B).
While monitoring the greenhouse experiment, we observed several fire ant-spider encounters. Spiders were frequently able to avoid ant attack by quickly moving to another part of the plant, by dropping off the plant, or by stretching to get their abdomen and cephalothorax as far from the plant surface as possible as if they were standing "on their tip toes." We believe these behaviors allow spiders to largely escape predation by fire ants. We hypothesize that the negative correlation between fire ant and spider abundance observed in the field arises from fire ants' dramatically suppressing the density of shared prey in cotton and, as a result, spiders emigrating from areas of high fire ant densities (Eubanks 2001). Competition among fire ants and other predators might also partially explain the negative effects of fire ants on other predators as well. Future research will address this question.
Not all of the natural enemies responded positively to ant suppression during the field experiment. The effect of fire ants on minute pirate bugs was inconsistent (Fig. 2D). We hope future research will help us understand the lack of a strong effect of fire ants on this species.
Our results are consistent with studies conducted in pasture and pecan agroecosystems, which reported significant levels of predation on other natural enemies by S. invicta workers (Bugg and Dutcher 1989, Tedders et al. 1990, Hu and Frank 1996, Dutcher 1998). Our results, however, are not consistent with previous studies in cotton asserting that fire ants do not interfere with other predators. Sterling et al. (1979) suggested that most predator taxa were unaffected by fire ants, and Reilly and Sterling (1983a,1983b) found no evidence of negative interactions between fire ants and other predators when they examined predator distributions within cotton fields. More thorough examinations of the effect of fire ants (Lopez 1982, Vinson and Scarborough 1989, Vinson 1994), as well as the results of this study, indicate that fire ants are major intraguild predators in most, if not all, agroecosystems they inhabit.
Although more work is needed to fully document the direct and indirect effects of fire ants on other arthropods, the results of this study support the conclusion that red imported fire ants are major intraguild predators. Future work will determine if the intense levels of intraguild predation by red imported fire ants reported here disrupt the natural enemy guild's ability to exert strong top-down control of herbivore populations. Because of the high densities of red imported fire ants in many parts of the United States and their continued range expansion into California and other key agricultural states, an ongoing effort to understand the effect of fire ants on managed and natural systems is imperative.
Carol Lawson and Will Eubanks helped with fieldwork. Kathy Flanders and John Ruberson provided insightful suggestions during early stages of the study. We thank James Bannon and the staff of the E. V. Smith Agricultural Experiment Station for allowing access to cotton fields at the station. Jeannine Eubanks, Chad Harvey, Ian Kaplan, and John Styrsky provided helpful comments on an early draft. This work was funded in part by the Alabama Fire Ant Management Program and the Department of Entomology and Plant Pathology at Auburn University.
References

{kind=link}
{kind=link}