





Abstract
We used five years of mist-net-capture and point-count data to quantify avian diversity in four habitats along a 1750-m elevational gradient in the Dominican Republic. These habitats include desert thorn scrub, dry forest, pine forest, and montane broadleaf forest, which together comprise more than two-thirds of existing forest on Hispaniola. In midwinter samples we recorded 74 species of landbirds, including 22 species of latitudinal migrants and 19 endemics. The highest diversity and species richness were found in pine forest and dry forest, but the highest capture rate of individuals was in desert thorn scrub. Abundance of migrant individuals was highest in pine forest, whereas pine and montane broadleaf forest contained the highest proportion of endemic species and individuals, and more habitat specialists. Among mist-net captures, insectivorous species and individuals predominated in all habitats except in dry forest, where more omnivorous individuals were captured. A more complex pattern was found in point-count detections: insectivorous species and individuals predominated in most habitats; omnivorous species and individuals were most frequently counted in montane broadleaf and dry forest, respectively; and nectarivorous individuals were most common in desert thorn scrub. Data presented here represent the most complete quantitative record of avian abundance and distribution on Hispaniola. This study not only details the value of these four habitats to various suites of species, but also emphasizes the importance of montane broadleaf and pine forests to large numbers of Neotropical migrants and Hispaniolan endemics, some of which are narrowly restricted to these habitats.
Comunidad de Aves de Invierno en Cuatro Hábitats a lo largo de una Gradiente Altitudinal en la Isla Española
Resumen. Utilizamos cinco años de datos de captura con redes ornitológicas y de conteos en puntos para cuantificar la diversidad de aves en cuatro hábitats a lo largo de un gradiente altitudinal de 1750 m en la República Dominicana. Estos hábitats incluyen matorral espinoso seco, bosque seco, bosque de coníferas y bosque latifoliado montano, los que en conjunto ocupan más de dos tercios del área boscosa del país. En enero y febrero, registramos 74 especies de aves terrestres, incluyendo 22 especies migratorias latitudinales y 19 endémicas. La mayor diversidad y riqueza de especies fue encontrada en el bosque de coníferas y en el bosque seco, pero la mayor tasa de captura de individuos se presentó en el matorral espinoso seco. La abundancia de individuos migratorios fue mayor en el bosque de coníferas, mientras que los bosques de coníferas y latifoliado montano presentaron la mayor proporción de individuos y especies endémicas y el mayor número de especialistas de hábitat. Entre las capturadas, las especies e individuos insectívoros predominaron en todos los hábitats, excepto en el bosque seco dónde se capturaron más individuos omnívoros. Entre las aves registradas en los puntos de conteo, los insectívoros también predominaron en la mayoría de los hábitats, pero registramos más especies omnívoras e individuos omnívoros en el bosque latifoliado nublado y bosque seco respectivamente. Los individuos nectarívoros fueron más comunes en el matorral espinoso seco. Los datos presentados aquí representan la cuantificación más completa de la abundancia y distribución de aves en Española. Esta investigación no sólo describe en detalle el valor de estos hábitats para los distintos grupos de especies, sino que también enfatiza la gran importancia de los bosques latifoliado montano y de coníferas para un gran número de aves migratorias Neotropicales y endémicas de Española, algunas de las cuales se encuentran estrechamente restringidas a estos hábitats.
Introduction
Hispaniola harbors one of the most diverse assemblages of birds in the Caribbean with more endemic bird species than any other Caribbean island except Jamaica. The island's two nations, Haiti and Dominican Republic, contain 15 species considered critically endangered, endangered, or vulnerable, and another six species considered nearly threatened with extinction or requiring further study (Birdlife International 2000). Its contribution to global biodiversity has earned Hispaniola the highest ranking of importance in a worldwide assessment of bird protection priorities (Stattersfield et al. 1998). Neotropical migratory birds are also an important component of the avifauna during winter on Hispaniola (Wunderle and Waide 1993), and Hispaniola is the principal nonbreeding ground of the range-restricted Bicknell's Thrush (Rimmer et al. 2001; scientific names of all species appear in Table 1). Because most of Haiti is almost entirely denuded (Paryski et al. 1989), the most promising opportunities for conservation lie in the Dominican Republic, where a dwindling forest cover may be vital to the survival of many endemic and migrant bird species. However, the loss of these habitats in the last 20 years has been estimated as >90% (Rappole 1995, Stattersfield et al. 1998), and most currently forested areas are fragmented and under continuing heavy pressure (Rimmer et al. 1998, Latta et al. 2000). In response to this crisis, the Dominican government has established a number of protected areas. However, patterns of beta diversity, local endemism, and avian species turnover have not been quantified, even though these are some of the most critical areas of research needed to design reserve networks (Olson et al. 2002). Moreover, little is known of the habitat needs and biology of many Hispaniolan bird species, and management decisions are typically made without crucial biological knowledge.
Occurrence of birds in desert thorn scrub, dry forest, pine forest, and montane broadleaf forest in the Sierra de Bahoruco, Dominican Republic. P-values for among habitat comparisons are based on Kruskal-Wallis test. *Indicates significance at a tablewide alpha level of P < 0.05 using the sequential Bonferroni technique. Residency categories: R = permanent resident; ER = endemic resident; IR = introduced resident; M = Neotropical migrant. Diet categories: O = omnivore; F = frugivore; I = insectivore; N = nectarivore; G = granivore; C = carnivore. Status categories: S = habitat specialist, G = habitat generalist
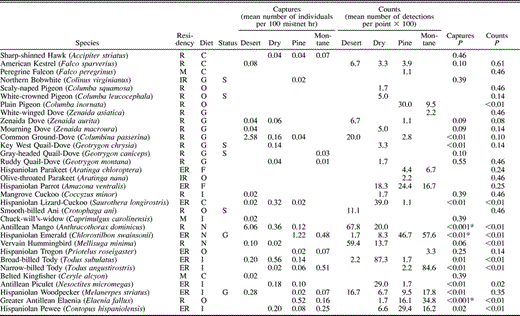
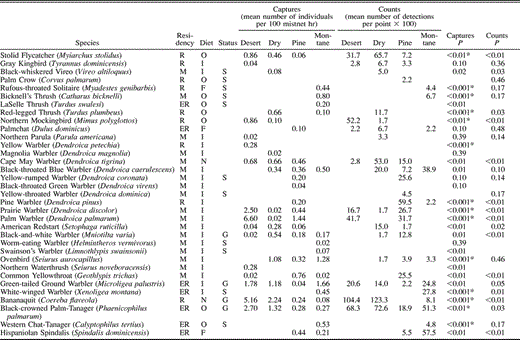
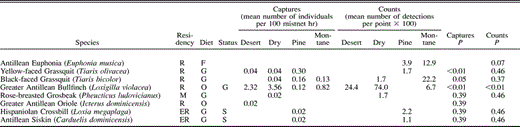
Occurrence of birds in desert thorn scrub, dry forest, pine forest, and montane broadleaf forest in the Sierra de Bahoruco, Dominican Republic. P-values for among habitat comparisons are based on Kruskal-Wallis test. *Indicates significance at a tablewide alpha level of P < 0.05 using the sequential Bonferroni technique. Residency categories: R = permanent resident; ER = endemic resident; IR = introduced resident; M = Neotropical migrant. Diet categories: O = omnivore; F = frugivore; I = insectivore; N = nectarivore; G = granivore; C = carnivore. Status categories: S = habitat specialist, G = habitat generalist
Few investigators in the West Indies have made a quantitative analysis of avian assemblages or taken a multihabitat approach to investigate both permanent resident and winter resident Neotropical migratory species. While the structure of West Indian bird communities has been the focus of a number of earlier studies (Ricklefs and Cox 1972, Lack 1976, Terborgh and Faaborg 1980, Case et al. 1983, Faaborg 1985), these primarily relied on the presence or absence of species on particular islands in order to elucidate how biotic factors, such as competition, resource abundance, vegetation structure, and habitat area; and abiotic factors, such as rainfall and temperature regimes, might influence abundance and distribution patterns of species or trophic groups of birds (Terborgh 1971, Beehler 1981). Several more recent studies have attempted to quantify avian abundance on various Caribbean islands (Faaborg et al. 1984, Askins et al. 1992, Wunderle and Waide 1993, Wallace et al. 1996), but these studies focused on Neotropical migratory birds and their integration into the resident bird community.
The lack of quantitative data from Hispaniola is especially striking. Beyond surveys of species occurrence and distribution (Wetmore and Swales 1931, Keith et al., in press), Terborgh and Faaborg (1980) used estimates of the proportion of migrants in local bird populations at eight sites across four forest types to assess factors determining migrant distribution during the winter months. Wunderle and Waide (1993) surveyed overwintering Neotropical migrants in a variety of habitats in the Dominican Republic, but they did not detail counts of permanent residents other than to report the number of resident species captured. In the most quantitative surveys to date, Wunderle and Latta (1996) sampled birds in sun and shade coffee plantations and in remnant pine forest in the Cordillera Central, and Latta and Wunderle (1998) characterized foraging behavior of birds in pine forests in the same mountains, but these surveys did not extend into other habitats.
Here we quantify avian diversity in four habitats along an elevational gradient in the Sierra de Bahoruco, Dominican Republic. Habitat-types studied represent 67% of 20 077 km2 of existing forest and “matorral” (brushland or scrub) in the Dominican Republic (Tolentino and Peña 1998), and include desert thorn scrub (18% of the total), dry forest (28%), pine forest (15%), and montane broadleaf forest (6%). The major objectives of this paper are to (1) describe species richness, avian diversity, and turnover along a 1750-m elevational gradient in Hispaniola, (2) describe patterns of change among trophic groups at different elevations, (3) determine the relative abundance of migrants and residents among the available habitats, and (4) determine the relative importance of each habitat to migratory and endemic species of highest conservation importance.
Methods
Study Sites
We studied bird communities in four habitats during the winters of 1996–1997 through 2000–2001. We established study sites near Cabo Rojo and the Sierra de Bahoruco, Dominican Republic (18°0′N, 71°38′W). The Bahorucos are an ancient chain of mountains that constitute a center of Hispaniolan endemism and are of extreme biological importance (Fisher-Meerow and Judd 1989). These mountains contain 25 of 26 endemic species of birds found on the island, including the threatened or endangered Hispaniolan Parrot, Hispaniolan Parakeet, LaSelle Thrush, Western Chat-Tanager, and White-winged Warbler. The Dominican Republic recognized the biological uniqueness of the Bahorucos by creating the 800-km2 Sierra de Bahoruco National Park in 1983. The area is ideal for field investigations because of its biological importance, the diversity of habitats available, and the proximity of large blocks of protected and relatively undisturbed habitat.
Study sites of 12–15 ha were established as follows: three study sites in low-elevation desert thorn scrub (20–50 m elevation; reduced to two study sites after 1999); two study sites in mid-elevation dry forest (300–365 m); three study sites in high elevation pine forest (1100–1475 m; reduced to two study sites after 1999); and three study sites in montane wet broadleaf forest (1675–1750 m; hereafter “montane forest;” reduced to two study sites after 1996). All study sites were >1.5 km from the next nearest site, were imbedded in a landscape of similar habitat, and were at least 150 m from habitat edges. While there was evidence that these habitats were variously affected by fire (pine forest), grazing (desert thorn scrub), timber cutting (pine forest), and tree-cutting for charcoal (desert thorn scrub, dry forest), the sites have been relatively undisturbed in the last 30 years.
Temperature and rainfall vary across the habitats. Daytime highs are similar across elevations (Latta 2000), but mean temperatures in January vary from 25°C at sea level to about 8°C at the highest elevations. Rainfall generally increases with elevation. At coastal levels rainfall averages <500 mm per yr, while at the highest sites rainfall exceeds 1700 mm per yr. These patterns are complicated, however, by differences in soil water-holding ability, lateral flow, evapotranspiration, and wind velocity (see Fisher-Meerow and Judd 1989 for a more complete discussion of the vegetation and ecology of these sites).
Desert thorn scrub sites
Low-elevation desert thorn scrub is found on a characteristic dogtooth limestone substrate with very little exposed soil (Fisher-Meerow and Judd 1989). Vegetation consists of widely scattered broadleaf trees that are partially deciduous, small broadleaf shrubs, and various cacti. Common broadleaf tree species include Capparis cynophallophora, C. ferruginea, Guaiacum officinale, Haitiella ekmanii, Metopium brownei, Phyllostylon brasiliensei, and Plumeria obtusa. A foliage height profile (Latta and Brown 1999) shows a low, open canopy with an understory dominated by broadleaf shrubs. Few forbs are present on the rocky floor, and while cacti and succulents are also sparse, they do occasionally extend into the canopy. Canopy cover is spare and averages 22 ± 28% (all means are reported ± SD) with greatest cover in the 2–4 m height categories and a maximum height of only 6 m. Mean and median broadleaf tree heights are 1.4 ± 1.0 m and 1.2 m, respectively.
Dry forest sites
The most abundant trees in classic dry forest in the Sierra de Bahoruco include Capparis ferruginea, Zizyphus rignoni, Bursera simaruba, Cameraria angustifolia, Cordia buchii, and Plumeria obtusa; in disturbed areas Acacia macracantha, Prosopis juliflora, and Cassia atomaria may be found (Fisher-Meerow and Judd 1989). A foliage height profile shows a moderately low, mostly closed canopy, a few emergent trees, and an understory dominated by broadleaf shrubs. Only low forbs are present, and cacti and succulents are sparse. Canopy cover averages 94 ± 17% with greatest cover in the 4–8 m height categories and a maximum height of 22 m. Mean and median broadleaf tree heights are 10.6 ± 4.6 m and 8.0 m, respectively.
Pine forest sites
Pine forest in the Sierra de Bahoruco is dominated by Hispaniolan pine (Pinus occidentalis); the only other common tree is the palm Coccothrinax scoparia. In some areas, particularly those disturbed by roadcuts, burning, or natural forces, Trema lamarckiana is prominent. A well-developed shrub layer is present, and common broadleaf species include Cestrum brevifolium, Chamaescrista glandulosa, Coreopsis buchii, Hypericum hypericoides, Lyonia truncata, L. microcarpa, Myrica picardae, and Senecio picardae, as well as the succulent Agave antillana (Fisher-Meerow and Judd 1989). The ground is covered by a thick layer of grasses. A foliage height profile (Latta and Sondreal 1999) shows a fairly open canopy, a sparse intermediate layer of pine, and a dense mixed-broadleaf and pine understory. Canopy cover averages 51 ± 26% with greatest cover in the 6–15 m height categories and a maximum pine height of 23 m. Mean and median pine heights are 17.7 ± 4.9 m and 19.0 m, respectively. The intermediate layer also consists solely of pine. Broadleaf trees and shrubs form a dense ground cover and understory, with broadleaf trees extending to 2.5 m in height. Young pine are also present in the understory.
Montane broadleaf forest sites
High-elevation moist broadleaf sites are very diverse hardwood forests that are humid and heavily shaded. The most distinctive feature of these sites is the abundance of lianas and epiphytes, including orchids, ferns, and bromeliads. Canopy cover is 100% with a well-developed canopy, subcanopy, and understory. The most abundant tree species include members of the genera Cupania, Dendropanax, Guarea, Mecranium, Miconia, Myrcia, Piper, Psychotria, and Trema (Fisher-Meerow and Judd 1989).
Sampling Birds
We used point counts and mist nets as complementary indices of abundance of birds (Ralph and Scott 1981, Ralph et al. 1993). We restricted our studies to landbird species. We sampled birds in midwinter because at this time of year there are significantly fewer nonterritorial migratory birds (“wanderers” or “floaters”) present in these habitats (Wunderle and Latta 2000, Latta 2000). In desert thorn scrub, dry forest, and pine forest, birds were sampled from 11 January–1 February 1997, 6–29 January 1998, 9–30 January 1999, 19 January–1 February 2000, and 27 January–8 February 2001. In montane forest, birds were sampled 6–13 December 1996, 5–12 November 1997, 8–16 November 1998, 24–27 January 2000, and 30 January–5 February 2001.
Birds were sampled with mist nets (12 m × 3 m × 30 mm mesh) set in three roughly parallel rows (100–150 m apart) of 10–16 nets each in desert thorn scrub, pine forest, and montane forest. Nets in the dry forest were placed in a single line of 31–39 nets over approximately 700 m to take advantage of existing trails in this very dense habitat. We did not view edge effects as a problem in these sites because the path was very narrow and only occasionally traveled by humans or domestic animals. In any given site, nets were opened mid-afternoon to dusk of day 1, dawn to dusk of day 2, and dawn to late morning of day 3, with an additional day of netting in montane sites. Because mist nets were set along transects, net placement was random with respect to physical features of the local environment that may have influenced capture rates of birds. All mist-netted birds were identified to species, age (second year or after second year), and sex by plumage characteristics (Pyle et al. 1987) when possible. Most birds were uniquely banded with both a numbered metal band and color bands for identification in the field. Birds that were too small to receive the smallest available metal band (i.e., hummingbirds, todies) had a rectrix or outer primary feather clipped so that recaptures within a netting session could be identified. Abundance of birds, or capture rate, was expressed as the number of birds captured per 100 mist-net hr, where one 12-m mist net opened for 1 hr = 1 mist-net hr. Recaptures from the same netting period were not included in capture rates. For analyses we calculated the mean capture rate of each species per habitat in each year.
Mist nets are subject to several familiar biases (Karr 1981, Remsen and Parker 1983), including unequal sampling of vegetation strata, and unequal capture rates among species and over time. This study minimized some of these problems: vegetation in these habitats is relatively low except in the pine forest; we limited analyses of net capture frequencies to within-species comparisons and assumed equal capture probabilities within species between habitats; and our mist-netting schedule (2–3 days every six weeks) minimized net shyness of birds.
We also conducted 10-min, 25-m fixed-radius point counts (Hutto et al. 1986) at ≥6 points in each study site each year. Points were situated in a grid except in the linear dry forest study sites, where points were placed along a transect. In all cases, each census point was 150 m from the next closest point. All point counts began at sunrise, and were completed by 09:30. No point counts were conducted in inclement weather. For analyses we calculated the mean number of detections of birds per point (×100) at each site and in each habitat in each year.
We classified birds captured in mist nets or recorded in point counts into groups based on diet and migratory status. Birds were grouped by diet on the basis of principal food items consumed in optimal habitats (Faaborg 1985, Wunderle and Latta 1996, SCL, unpubl. data). Groups included insectivores, nectarivores, granivores, carnivores (primarily predators of lizards or other birds), and omnivores (including birds that were primarily frugivores). Groups based on migratory status included residents (which are present throughout the year) and temperate (latitudinal) migrants which breed north of the tropics. One species, the Black-whiskered Vireo, which is mostly a migratory population (not present September–January) from Hispaniola to wintering grounds in northern South America (Keith et al., in press), was also included in the group of migrants.
Statistical Analyses
The software package SYSTAT Version 5.2.1 (Wilkinson 1992) was used to perform various statistical tests described by Sokal and Rohlf (1995). Data were tested for normality using normal probability plots and tests of skewness and kurtosis. When data were not normally distributed and could not be transformed to achieve normality, nonparametric statistics were used. A probability of Type I error of 0.05 or less was accepted as significant unless otherwise noted. We did not systematically analyze local variation in bird populations (within habitats), but rather pooled samples from different sites for each habitat type to increase sample sizes. Some analyses are based on presence/absence of species or proportions of birds captured or observed, while other comparisons are based on capture rates or mean number of individuals recorded in point counts.
We compared species diversity among habitats by calculating species richness and evenness. We used rarefaction (Simberloff 1972) to compare species richness in different habitats with data obtained from point count and mist-net samples. Rarefaction curves are essentially idealized species-accumulation curves that allowed us to compare the expected species richness of the different habitats for a constant sampling effort at each site. In addition, we also calculated an index of evenness for each habitat, and used a t-test to test the hypothesis that there is no difference in diversity of birds among habitats (Magurran 1988). We calculated numerical dominance of species captured or detected in point counts within each habitat by ranking by abundance all species combined, all latitudinal migrants, and all endemics, then calculating the percentage of total individuals represented by the five most abundant species, the three most abundant migrants, and the two most abundant endemics. In determining numerical dominance we sought to include as many species within a category (all species, latitudinal migrants, endemics) as was needed to find percent dominance in most categories >50% but <90%.
We used similarity measures to determine how habitats differed from one another. Jaccard's index (Magurran 1988) was used to compare similarity of habitats based on the presence or absence of species in netting and point-count samples among habitats, and Sorenson's measure (Magurran 1988) was used to compare the similarity of habitats based on the proportional abundance of species recorded in netting and point counts, and the proportional abundance of birds in each diet category. Spearman's rank correlation coefficient was used to quantify the association between migrant and resident abundance within each habitat.
The Kruskal-Wallis test was used to compare the abundance of individuals captured in mist nets (capture rate), and the abundance of individuals detected in point counts (detection rate) among habitats, with capture rates and detection rates averaged across years. These tests entailed 62 planned comparisons for each of the species captured in mist nets, and 61 planned comparisons for each of the species detected in point counts, so we used the sequential Bonferroni technique (Rice 1989) and decreased the tablewide level of alpha in order to reduce the probability of committing a Type I error (Sokal and Rohlf 1995). We used row × column tests of independence with a G-statistic to test for significant heterogeneity in (1) the proportion of species and individuals in a diet category among habitats, (2) the proportion of latitudinal migrant species and individuals among habitats, (3) the proportion of endemic species and individuals among habitats, and (4) the proportion of males of selected species among habitats.
Results
Patterns of distribution of species and individuals
We recorded 74 species of landbirds (Table 1) among 4582 net captures and 1403 point-count detections in the Sierra de Bahoruco, Dominican Republic, in midwinter surveys that included 29 876 net hr and 242 point counts across all four major habitat types (Table 2). Detections of birds were not equal among habitats. Capture rates were higher in the desert thorn scrub (33.2 birds captured per 100 mist-net hr) than in dry forest (14.6 birds captured per 100 mist-net hr), pine forest (8.3 birds captured per 100 mist-net hr), or montane broadleaf habitat (8.8 birds captured per 100 mist-net hr). A similar pattern was seen in point counts, with detection rates highest in desert thorn scrub (6.3 birds detected per point count) and dry forest (7.2 birds detected per point count), and lowest in pine forest (4.4 birds detected per point count) and montane sites (5.2 birds detected per point count).
Number of captures (mist nets) and detections (fixed-radius point counts) of Neotropical migratory and permanent-resident species in four habitats in the Sierra de Bahoruco, Dominican Republic, 1996–2001
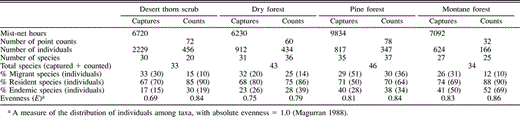
Number of captures (mist nets) and detections (fixed-radius point counts) of Neotropical migratory and permanent-resident species in four habitats in the Sierra de Bahoruco, Dominican Republic, 1996–2001
Among-habitat differences in species richness were similar whether based on mist-net captures or point counts, with richness highest in pine forest, followed by dry forest, montane forest, and desert thorn scrub (Fig. 1). Rarefaction curves of species richness based on mist-net captures indicated that some new species were being added in all habitats except montane forest. Curves of species richness based on point counts also suggested that a few new species were being added, although at a slower rate, as curves for each habitat (except montane) reached an asymptote.
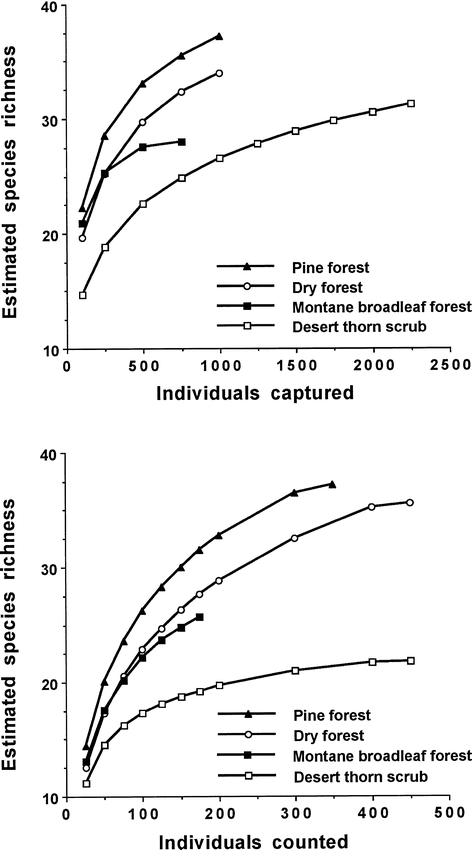
Species rarefaction curves based on number of mist-net captures or number of individuals detected in point counts in four habitats in the Sierra de Bahoruco, Dominican Republic, 1996–2001
Evenness was generally high and similar among habitats (Table 2) except for mist-net captures in desert thorn scrub, where evenness was a relatively low 0.69. In all other cases, evenness ranged from 0.75–0.83 for birds captured in mist nets and from 0.79–0.86 for birds recorded in point counts. The extent of numerical dominance of species within a habitat also suggests the degree of evenness within that habitat. In terms of mist-net captures (Table 3), five species accounted for 69% of captures in desert thorn scrub where evenness was relatively low, whereas five species accounted for 64% of captures in dry forest, 52% of captures in pine, and 57% of captures in montane forest; the pattern is similar in terms of point-count detections. Among latitudinal migrants (Table 3), numerical dominance of the three most abundant species was highest in desert thorn scrub (96% of captures, 100% of point count detections) and montane forest (91% of captures, 100% of point count detections), while among endemic species (Table 3), numerical dominance of the two most abundant species was much higher in desert thorn scrub (90% of captures, 80% of point count detections) than in any other habitat type.
Numerical dominance among all species, all migrants, and all endemic species in desert thorn scrub, dry forest, pine forest, and montane broadleaf habitats in the Sierra de Bahoruco, Dominican Republic, 1996–2001, as determined by mist-net captures and point counts. Numbers represent rank in abundance in captures (point counts) in each habitat
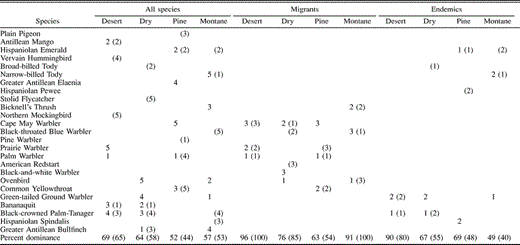
Numerical dominance among all species, all migrants, and all endemic species in desert thorn scrub, dry forest, pine forest, and montane broadleaf habitats in the Sierra de Bahoruco, Dominican Republic, 1996–2001, as determined by mist-net captures and point counts. Numbers represent rank in abundance in captures (point counts) in each habitat
Of the 74 species recorded, 22 (30%) were latitudinal migrants. The proportion of species detected that were migrants varied from 26–33% among habitats (Table 2). Although the ratio of migrant to resident species that were mist netted did not vary significantly among habitats (G3 = 2.0, P = 0.57), the ratio of migrant to resident individuals mist netted did (G3 = 22.5, P < 0.001). Migrants comprised 51% of the individuals mist netted in pine forest, whereas only 20% were migrants in dry forest. Similarly, there was a significant difference in the ratio of migrant to resident species (G3 = 12.5, P = 0.01; Table 2) and individuals (G3 = 30.2, P < 0.001) recorded in point counts among habitats, with most records of migrant species and individuals again from pine habitat.
Of the 74 species recorded, 19 (26%) were endemic residents. The proportion of endemic species and individuals also varied among habitats (Table 2). There was a significant difference in mist-net captures of endemic species (G3 = 21.4, P < 0.001) and individuals (G3 = 30.6, P < 0.001) among habitats, with the highest proportion of endemic species recorded in pine and montane forest, and the highest proportion of endemic individuals mist netted in montane habitat. Point-count data showed a similar pattern with a significant difference in the proportion of endemic species (G3 = 28.1, P < 0.001) and endemic individuals (G3 = 75.6, P < 0.001) counted among habitats, and the highest proportions recorded in montane forest.
In most cases we found low correlations between migrant and resident abundance within habitats. Using mist-net capture data, we found low correlations between the number of migrant and resident species (rs = 0.54), and individuals (rs = 0.20) captured. Point-count data also showed a low correlation between numbers of migrant and resident individuals within a habitat (rs = 0.20), but a higher correlation between migrant and resident species detected in point counts (rs = 0.74).
Our data suggest the presence of both habitat specialists and generalists in these sites. Of the 74 species of recorded landbirds, 30 (40%) were found in only one habitat (Table 1). Some of these (10 species) were locally uncommon and only recorded 1–2 times; others (2 species) were normally found in mangrove adjacent to desert thorn scrub, but mangrove was not extensive in area nor consistently sampled and so is not addressed here. However, 18 species were recorded multiple times in only one habitat suggesting that they are habitat specialists in winter. More habitat specialists were found in montane forest (8 species) and pine forest (6 species) than in dry forest (3 species) or desert (1 species).
Seven species were recorded across all four habitats and may be considered habitat generalists. Among these, two showed significant differences in abundance among habitats (Table 1). The Bananaquit (H = 17.0, P < 0.001) and the Black-crowned Palm-Tanager (H = 15.9, P < 0.01) were most common in captures in desert thorn scrub and dry forest sites. Five additional species showed large but nonsignificant differences in abundance among habitats (Table 1). The Hispaniolan Emerald was most abundant in counts in pine and montane forest sites, while captures suggested that the Black-and-white Warbler preferred dry forest and pine habitat, and the Greater Antillean Bullfinch preferred desert thorn scrub and dry forest sites. The Hispaniolan Woodpecker and the Green-tailed Ground Warbler both showed the most unusual distribution of these generalist species, being more abundant in low-elevation desert thorn scrub and high-elevation montane sites.
Beyond those species identified as habitat specialists or generalists, 37 species were found in 2–3 habitats. Within-species tests showed a significant difference in abundance in net captures of 11 of 33 (33%) species occurring in multiple (2–4) habitats, but there were no significant differences in abundance among habitats for the 35 species reported in multiple point counts (Table 1). Nevertheless, there were few cases in which the trend for differences among habitats in capture rates was not also seen in differences among habitats in point-count detections.
Broadly occurring Neotropical migratory species may segregate by sex on the wintering grounds. We were able to reliably determine sex in four species that were mist netted with adequate sample sizes for analysis (Fig. 2). There was a significant difference in the proportion of males among habitats for Cape May Warbler (G2 = 13.2, P < 0.001), Black-throated Blue Warbler (G2 = 54.2, P < 0.001), and Black-and-white Warbler (G2 = 17.1, P < 0.001), but not for Prairie Warbler (χ21 = 0.1, P = 0.89). Among Cape May Warblers and Black-and-white Warblers, males tended to use dry forest and pine forest, whereas among Black-throated Blue Warblers males predominated in dry forest.
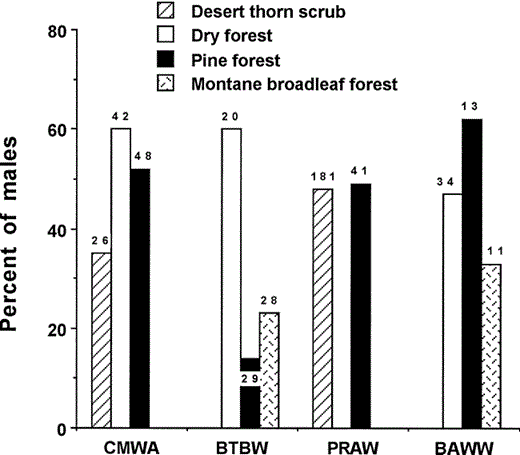
Percentage of male birds of four Neotropical migratory species captured in mist nets in four habitats in the Sierra de Bahoruco, Dominican Republic, 1996–2001. (CMWA = Cape May Warbler, BTBW = Black-throated Blue Warbler, PRAW = Prairie Warbler, BAWW = Black-and-white Warbler; scientific names are found in Table 1). Numbers above bars are sample sizes
Patterns of distribution of trophic groups
The proportion of mist-netted species (G12 = 23.6, P = 0.02) and individuals (G12 = 59.9, P < 0.001) in each diet category varied significantly among habitats (Fig. 3a). Similarly, the proportion of species (G12 = 46.2, P < 0.001) and individuals (G12 = 69.5, P < 0.001) in each diet category recorded in point counts varied significantly among habitats (Fig. 3b). Mist-net capture data (Fig. 3a) showed that insectivorous species and individuals were proportionately more common in all habitats except dry forest, where more omnivorous individuals were mist netted. Nectarivores were prominent in desert thorn scrub and pine forest, whereas omnivores were also common in montane forest. Data from point counts showed a more complex pattern (Fig. 3b) with insectivorous species proportionately more common in all habitats except montane forest, where omnivores were most frequent. Among individuals however, insectivores were proportionately more common only in pine forest and montane forest, whereas nectarivores were most common in desert thorn scrub. Omnivores were most common in dry forest, and were moderately common in desert and montane point counts.
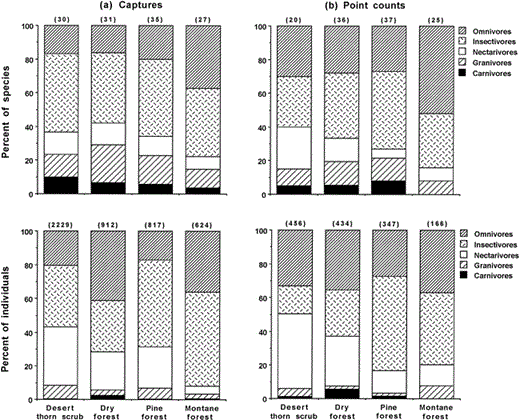
Frequency of five trophic groups of birds in four habitats of the Sierra de Bahoruco, Dominican Republic, 1996–2001, as measured by number of species (top graphs) or individuals (bottom graphs) that were (a) caught in mist nets or (b) detected in point counts. Numbers above bars are sample sizes
Similarities among habitats
Similarity indices based on species presence/absence in a habitat were generally low, with scores of 0.27–0.44 for most habitat pairs (Table 4). Scores tended to be higher between neighboring habitats. Similarity indices based on the proportion of individuals per species mist netted or recorded in point counts (Table 4) were somewhat lower than scores based on presence/absence, with most scores ranging from 0.17–0.42. In contrast, similarity indices based on trophic groups were quite high among habitats with all scores ranging from 0.57–0.93 (Table 5). Once again, scores tended to be highest among neighboring habitats.
Jaccard similarity indices based on species presence/absence, and Sorenson similarity indices based on proportion of individuals per species (in parentheses), in four habitats in the Sierra de Bahoruco, Dominican Republic, 1996–2001. Similarity indices range from 0 (entirely different) to 1 (complete overlap)
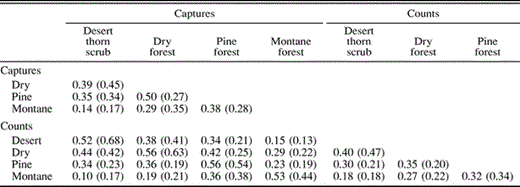
Jaccard similarity indices based on species presence/absence, and Sorenson similarity indices based on proportion of individuals per species (in parentheses), in four habitats in the Sierra de Bahoruco, Dominican Republic, 1996–2001. Similarity indices range from 0 (entirely different) to 1 (complete overlap)
Sorenson similarity indices for quantitative data based on proportion of individuals per trophic group (insectivores, granivores, carnivores, nectarivores, and ominivores; see Table 1) captured in mist nets or detected in point counts in the Sierra de Bahoruco, Dominican Republic, 1996–2001. Similarity index ranges from 0 (entirely different) to 1 (complete overlap)
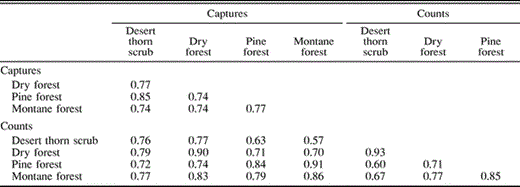
Sorenson similarity indices for quantitative data based on proportion of individuals per trophic group (insectivores, granivores, carnivores, nectarivores, and ominivores; see Table 1) captured in mist nets or detected in point counts in the Sierra de Bahoruco, Dominican Republic, 1996–2001. Similarity index ranges from 0 (entirely different) to 1 (complete overlap)
Discussion
Community Composition
Relative abundance data presented here for 74 species of Hispaniolan landbirds (including 22 Neotropical migrant species and 52 resident species) suggest that, in general, Hispaniolan habitats support a large number of avian species and individuals compared to other islands in the Bahamas and Greater Antilles (Lack 1976, Raffaele et al. 1998, Keith et al., in press). These data compare favorably to similar data from Cuba (23 migrant species, 39 resident species; Wallace et al. 1996), and Grand Bahama Island (19 migrant species, 33 resident species; Emlen 1977), the only other islands for which relative abundance data of commonly occurring winter and permanent resident landbirds have been published.
Patterns of species richness across these habitats indicate that our sampling was fairly complete, although some new species were still being recorded in most habitats at the end of the study. Like previous studies from other Neotropical sites (Blake and Loiselle 2000), we observed a general pattern of species richness increasing with elevation, except that our highest montane site had relatively low richness. Wunderle and Waide (1993) also found low richness in montane broadleaf forest, and relatively high species richness in pine forest, although not as high as reported here. Rarefaction curves in the Sierra de Bahoruco pine forest may be high in comparison to that of pine forest in the Cordillera Central because results from the latter were derived from smaller sample sizes collected over only one year, and pine forest sites in the Cordillera Central are fragments of the original pine forest. Fragmentation has been shown to negatively impact Hispaniolan Crossbill populations (Latta et al. 2000), and flycatchers and tanagers in the Dominican Republic (Robbins et al. 1987), and may affect other Hispaniolan populations as well.
Trophic Groups
Differences in avian community composition among habitats may be a result of differences in resources available to the birds and thus be reflected in the trophic groups present in each habitat. In low-elevation desert thorn scrub, insectivores and omnivores were the predominant species in point counts, but nectarivores were the most abundant individuals, owing to the numerical dominance of Bananaquit, Antillean Mango, and Vervain Hummingbird. Among net captures in desert sites, insectivores were proportionately most common while individual captures were dominated by insectivores (especially Prairie and Palm Warblers) and the nectarivores mentioned above. In dry forest, insectivores were most common in both point counts and mist-net samples, but omnivores were proportionately the most abundant individuals with large numbers of Greater Antillean Bullfinch, Black-crowned Palm-Tanager, and Stolid Flycatcher recorded. At pine forest sites, a suite of warblers including the resident Pine Warbler, and migratory Palm Warbler and Common Yellowthroat, contributed to insectivore domination of all measures of abundance. Finally, in montane forest, insectivores again dominated but omnivores had a strong presence especially in the proportion of species recorded. This is the result of the presence of numerous, mostly frugivorous endemic species, many with small populations, including the Hispaniolan Parakeet, Hispaniolan Parrot, Hispaniolan Trogon, LaSelle Thrush, Western Chat-Tanager, and Hispaniolan Spindalis.
The general domination of the avian assemblage by insectivores in most habitats is consistent with previous work in Hispaniolan pine forests (Wunderle and Latta 1996, Latta and Wunderle 1998), but is contrary to earlier suggestions that frugivores and omnivores dominate in other Caribbean habitats (Lack 1976). Our sampling indicates that in these habitats insectivory predominates during the nonbreeding period. However, frugivores and omnivores are likely to be proportionately more abundant during the breeding season. Because among wintering migrants in the West Indies, small gleaning insectivores are overrepresented while frugivores and granivores are conspicuously underrepresented (Faaborg and Terborgh 1980), proportional abundance indices shift toward insectivores in winter. Although the distribution of trophic groups among habitats should be quantified during the breeding season, our data are consistent with Faaborg's (1985) assertion that once the migrants depart, the frugivore guild is proportionately the most abundant and complex guild in the West Indies.
Endemic Species
The assemblage of birds in all habitats was dominated by residents as opposed to migrants. There was little indication that habitats that support large numbers and many individuals of residents also support similar numbers of migrants, as we found low correlations between most measures of migrant and resident abundance within a habitat. This suggests that migrants and residents require different resources, as suggested by the analyses of trophic groups. Among the permanent-resident species, most endemics were habitat specialists found in pine forest and montane forest. The relegation of endemic species to narrowly defined, high-elevation habitats has been observed previously by Ricklefs and Cox (1972), who suggested that these species are more ancient taxa (Ricklefs and Bermingham 1999) which have been pushed into progressively fewer habitats by more recent immigrants of superior competitive abilities. Although many endemic species also used desert thorn scrub habitat and dry forest, there were no endemic specialists confined primarily to these habitats.
Neotropical Migrants
Across all habitats, we found that migrants comprised 30% of the species encountered, and 32% of mist-netted individuals. These findings are similar to those of previous surveys of the West Indies that found that migrants constituted 30–47% of the species detected (Lack 1976, Arendt 1992, Wunderle and Waide 1993). The proportion of wintering migrants to residents results in part from the combined effects of island area and distance to continental North America (Terborgh and Faaborg 1980, Wunderle and Waide 1993). However, habitats on the same island often differ substantially in suitability, with migrant species abundance significantly related to elevation, rainfall, and vegetation complexity (Wunderle and Waide 1993). Differences in migrant abundance among habitats was clearly seen in this study, but patterns did not strictly follow elevational or moisture gradients. More than 50% of individual birds mist netted in pine forests were migrants, and pine forest also had a more diverse collection of migrants than other habitats. This is reflected in the high proportion of migrant species recorded in point counts in pine forests, and in the low numerical dominance of the most commonly captured and counted species. High avian diversity in the pine forest is undoubtedly related to the complexity of the vegetation, which includes a pine overstory and a well-developed broadleaf understory (Latta and Wunderle 1998). Pine forest has also been shown to support greater numbers of migrants than xeric habitats on a variety of other islands (Wunderle and Waide 1993), including Cuba (Wallace et al. 1996).
In contrast to pine forest, we found a low proportion of migrant species and individuals in point-count data from our most xeric site (desert thorn scrub) and our most mesic site (montane forest). Montane habitat was used by a diverse suite of species including a habitat generalist (Ovenbird), female Black-throated Blue Warblers, which have been shown elsewhere to primarily occupy shrubby second-growth and matorral (Wunderle 1995), a rare and range-restricted species (Bicknell's Thrush), and a habitat specialist (Swainson's Warbler). The low-elevation desert thorn scrub sites were also depauperate in terms of migrant species, but numerically were dominated by Palm Warblers and Prairie Warblers. Interestingly, although these species occupied daytime territories (Prairie Warbler) or large home ranges (Palm Warbler) in desert thorn scrub, they moved to a narrow coastal strip of mangrove in the evening where they formed communal nighttime roosts, a behavior not noted in other habitats where they occurred (Latta and Faaborg 2001). It may have been the proximity of these mangrove roost sites that allowed these species to be so abundant in the desert thorn scrub, because similar desert sites farther from mangrove contained extremely few Palm or Prairie Warblers (SCL, unpubl. data). Mangrove is a favored site for a number of migrant species in the West Indies (Lynch 1989, 1992, Arendt 1992, Lefebvre et al. 1992, Wunderle and Waide 1993, Wallace et al. 1996), but mangroves in the vicinity of our study sites were not extensive enough to allow sampling.
Beyond a consideration of habitats of importance to Neotropical migrants, this study highlights a number of migrant species that may be of conservation concern. Particularly abundant species included Bicknell's Thrush, Cape May Warbler, Black-throated Blue Warbler, Prairie Warbler, Palm Warbler, Black-and-white Warbler, and Ovenbird. All of these species except Black-and-white Warbler and Ovenbird have winter ranges largely restricted to the Greater Antilles and so are of special conservation concern and should be monitored (Wunderle and Waide 1994, Rimmer et al. 2001, Latta and Faaborg 2002). In addition, this study indicates a number of species which occurred in a single habitat and so may be considered habitat specialists of conservation importance, including Yellow-rumped and Yellow-throated Warblers in pine forest, and Bicknell's Thrush, Worm-eating Warbler, and Swainson's Warbler in montane forest. The Yellow-throated, Worm-eating, and Swainson's Warblers were also identified as habitat specialists by Wunderle and Waide (1994), but Yellow-rumped Warbler was labeled a habitat generalist as it was found in moderate numbers in a variety of other forest types on other Caribbean islands. The Bicknell's Thrush has also been shown to be a habitat specialist whose winter populations are largely concentrated in montane broadleaf forests (Rimmer et al. 2001).
Conservation of Birds on Hispaniola
The importance of structural habitat complexity and taxonomic diversity of vegetation to avian diversity has been pointed out by many authors (MacArthur and MacArthur 1961, Karr and Roth 1971, Holmes et al. 1979). Wunderle and Latta (1996) cited diversity of vegetation as a factor in the higher avian species richness found in Dominican pine forest in the Cordillera Central when compared to coffee plantations because complex habitats offered a wider variety of foraging sites. Our data support this conclusion in that most trophic groups were broadly represented in the most complex pine habitat. Montane and dry forest sites could also be considered relatively complex, but desert thorn scrub was the simplest habitat in terms of vegetation structure and had the lowest avian diversity. While habitat and structural complexity are important, both types of complexity may be negatively influenced by habitat alterations such as selective cutting, timber harvesting, or other disturbances. Latta et al. (2000) suggested that the historical cutting of the pine forests on Hispaniola had altered stand dynamics resulting in increased incidence of fire and, perhaps more importantly, hotter-burning fires which were severely threatening habitat for the endemic Hispaniolan Crossbill.
Although this study was not designed to examine elevational migrations, they are a distinct possibility. Birds may regularly move to lower elevations following food resources (Loiselle and Blake 1991), or sites at one elevation may become important to birds on an irregular basis such as after storm-related habitat alterations at other elevations (Winker et al. 1997). This may be of particular relevance to birds in the West Indies where large movements of highland birds to lower elevation habitats have been documented following hurricanes (Wunderle et al. 1992). The probability that elevational migrations are at times important to Hispaniolan bird species argues for the need to protect habitats along entire elevational gradients.
In terms of conservation and the design of reserve networks, this study not only details the value of all four habitats to various suites of species, but also emphasizes the importance of montane and pine forests to large numbers of Neotropical migrants, residents, and Hispaniolan endemics, some of which are narrow habitat specialists. Montane forest has been previously recognized as one of the most endangered habitats on the island (Latta and Lorenzo 2000), not only because it has been reduced to less than 1200 km2 islandwide (Tolentino and Peña 1998), but also because it is of critical importance to so many of the island's endemics and most severely threatened species. Pine forest, although currently more extensive in coverage (slightly more than 3000 km2 remaining; Tolentino and Peña 1998) is declining under severe disturbance from wildfire, agriculture, grazing, and cutting for fuelwood and building materials (Hartshorn et al. 1981, Lugo et al. 1981, FAO 1991), with a number of bird populations having been adversely impacted (Arendt 1992, Latta et al. 2000).
Data presented here represent the most complete quantitative record to date of avian abundance and distribution across a variety of widespread habitats on Hispaniola. Given that Haiti is almost entirely deforested (Paryski et al. 1989), these data on avian abundance and diversity, species richness, and turnover among permanent residents, endemics, migrants, and foraging guilds are representative of a significant proportion of Hispaniolan habitats. This study is an important step forward in our ability to understand patterns of habitat use, and it provides our best opportunity yet to assess habitat needs and conservation concerns of a variety of permanent resident and Neotropical migratory bird species on Hispaniola.
Acknowledgments
Field assistance was provided by J. Almonte, C. Brown, B. Cabrera, J. Chace, E. Cuevas, F. deLacour, J. Dolores, J. Farrington, H. Gamper, E. Garrido, B. Gibbons, J. Goetz, J. Hammond, M. Heredia, M. McFarland, D. Mejía, V. Mejía, R. Moore, B. Ryder, M. Sondreal, V. Stori, J. Tietz, T. Vargas, E. Vásquez, H. Walker, K. Wallace, and P. Wieczoreck. Funding or equipment was provided to SCL by the University of Missouri Research Board, National Fish and Wildlife Foundation, Wildlife Conservation Society, Association of Avian Veterinarians, and USDA Forest Service North-Central Forest Experiment Station. SCL was also supported by a STAR Graduate Fellowship from the US Environmental Protection Agency. Work of VINS in the Dominican Republic was supported by American Bird Conservancy, Blake Fund of the Nuttall Ornithological Club, Conservation and Research Foundation, National Geographic Society, National Fish and Wildlife Foundation, Wildlife Conservation Society, and U.S. Fish and Wildlife Service. An earlier version of this paper was reviewed by W. J. Arendt, R. Askins, J. Blake, D. S. Dobkin, J. Faaborg, and R. Ricklefs. Permission to work in the Sierra de Bahoruco was provided by the Dirección Nacional de Parques and the Departamento de Vida Silvestre, Dominican Republic.
Literature Cited
Author notes
Present address: PRBO Conservation Science, 4990 Shoreline Highway, Stinson Beach, CA 94970. [email protected]

{kind=link}
{kind=link}
{kind=link}