





Abstract
Motivation: The accurate discovery and annotation of regulatory elements remains a challenging problem. The growing number of sequenced genomes creates new opportunities for comparative approaches to motif discovery. Putative binding sites are then considered to be functional if they are conserved in orthologous promoter sequences of multiple related species. Existing methods for comparative motif discovery usually rely on pregenerated multiple sequence alignments, which are difficult to obtain for more diverged species such as plants. As a consequence, misaligned regulatory elements often remain undetected.
Results: We present a novel algorithm that supports both alignment-free and alignment-based motif discovery in the promoter sequences of related species. Putative motifs are exhaustively enumerated as words over the IUPAC alphabet and screened for conservation using the branch length score. Additionally, a confidence score is established in a genome-wide fashion. In order to take advantage of a cloud computing infrastructure, the MapReduce programming model is adopted. The method is applied to four monocotyledon plant species and it is shown that high-scoring motifs are significantly enriched for open chromatin regions in Oryza sativa and for transcription factor binding sites inferred through protein-binding microarrays in O.sativa and Zea mays. Furthermore, the method is shown to recover experimentally profiled ga2ox1-like KN1 binding sites in Z.mays.
Availability and implementation: BLSSpeller was written in Java. Source code and manual are available at http://bioinformatics.intec.ugent.be/blsspeller
Contact: [email protected] or [email protected]
Supplementary information: Supplementary data are available at Bioinformatics online.
1 Introduction
One of the major challenges in systems biology is gaining a full understanding of gene transcriptional regulation. Transcription factors, for which the binding sites are usually hidden in the promoter sequence of the gene, are in this respect of particular importance. Computational approaches for de novo motif discovery can be classified in (i) methods to identify binding sites in promoter sequences of co-regulated genes within a single genome and (ii) comparative approaches using homologous sequences from multiple related species (Das and Dai, 2007).
The first category uses clusters of co-expressed genes, which are assumed to be regulated by the same set of transcription factors. A drawback of these methods is that the relationship between co-expression and co-regulation relies on complex regulatory mechanisms, making it difficult to assemble reliable datasets since co-expression does not necessarily imply that there is a common binding site involved. Two different algorithmic approaches coexist: the statistical (Bailey etal., 2009; Hughes etal., 2000; Liu etal., 2001; Thijs et al., 2002; Wei and Yu, 2007) and the exhaustive, word-based algorithms. The latter contain graph-based approaches (Eskin and Pevzner, 2002; Liang etal., 2004; Satya and Mukherjee, 2004) and methods based on index structures (Marsan and Sagot, 2000; Marschall and Rahmann, 2009; Pavesi et al., 2001).
Due to the growing availability of genome sequences, a second category of algorithms based on phylogenetic footprinting emerged (Blanchette and Tompa, 2002): orthologous regulatory regions from multiple species are compared with the underlying assumption that functional elements evolve at a much slower pace, compared to the non-functional part of the genome, due to selective pressure (Berezikov etal., 2004). Most comparative motif discovery approaches rely in some way on multiple sequence alignments, in which regulatory signals are expected to be well-aligned. Pioneering algorithms in this category are Conreal (Berezikov etal., 2004), Phylonet (Wang, 2005) and Phyloscan (Carmack etal., 2007). More recent algorithms relying on alignments are used to study mosquitoes (Sieglaff etal., 2009), Fusarium (Kumar etal., 2010), vertebrates (Ettwiller etal., 2005) and mammals (Xie etal., 2005).
It has, however, been shown that known regulatory elements are not always correctly aligned (Siggia, 2005), an issue that is further complicated by the different alignments produced by various alignment programs (Pollard etal., 2004). Transcription factor (TF) binding sites are short, flexible against certain mutations and even mobile which explains why they are sometimes misaligned. Mechanisms have been observed that allow the modification of regulatory sequences without altering their function: divergence driving words and binding site turnover. Regulatory sequences can diverge freely if the divergence driving words, which are specific short words in the non-coding DNA, are not altered (Bradley etal., 2010). Since a TF can often bind to multiple similar sites, mutations turning one site into another should not affect regulation. Binding site turnover, on the other hand, is the mechanism where the gain of a redundant binding site allows the loss of a previously functional site (Venkataram and Fay, 2010). The corresponding TF can then bind to the new site, maintaining the regulatory interaction. This allows binding sites to relocate within the regulatory sequence, making it difficult for alignment algorithms to correctly align them.
Binding site discovery, especially in plants, has to deal with large divergence times and complex diversification mechanisms such as genome duplications. This makes approaches based on whole genome alignments, often used in de novo algorithms, impractical. Some of these problems have been addressed in earlier studies. Stark etal. (2007) used a mixed approach in a study with 12 Drosophila species, starting from whole genome alignments but allowing for limited motif movement within an alignment. Elemento and Tavazoie (2005) designed an alignment-free algorithm to discover overrepresented k-mers over the exact ACGT alphabet in pairs of related genomes. Finally, MDOS (Wu etal., 2008) is a new version of this algorithm with improved statistics.
In this article, four monocotyledonous plant species are studied using a phylogenetic footprinting approach: Oryza sativa ssp.indica (osa), Brachypodium distachyon (bdi), Sorghum bicolor (sbi) and Zea mays (zma). We adopt a gene-centric approach, where the promoter sequences of orthologous genes are grouped into gene families. A word-based discovery algorithm was designed to exhaustively report all genome-wide conserved motifs. The term conserved relates to the occurrence of the motif in multiple promoter sequences of a particular gene family. Genome-wide conservation relates to the fact that this conservation occurs in more gene families than what is expected by chance. Motifs are modeled as words (k-mers) over an alphabet that contains the four bases (ACGT) and (optionally) additional degenerate characters from the IUPAC alphabet (Cornish-Bowden, 1985). This degeneracy allows a motif to model a collection of binding sites. The algorithm can be run in both alignment-free or alignment-based mode. In case of alignment-free discovery, the conservation of a motif is scored irrespective of its orientation or position within a promoter sequence. This relaxed definition of conservation was previously used by Gordân etal. (2010) and is especially relevant when studying more diverged species for which accurate multiple sequence alignments are difficult to generate. Alignment-based discovery adds the constraint that motifs must be aligned, i.e. occur at the same position in the multiple sequence alignment.
Robust algorithms for comparative genomics are expected to gain in power when more related species are added. Most studies so far only consider motifs that are conserved within all organisms. The branch length score (BLS) was developed to quantify motif conservation in a biologically meaningful manner and ranges from 0% (not conserved) to 100% (conserved in all sequences). The BLS takes the phylogenetic relationships between the species into account by representing a relative evolutionary distance over which a candidate binding site is conserved within a gene family. The BLS was first used in a comparative study with 12 Drosophila genomes (Stark etal., 2007) and allows studying motifs only conserved in subsets of the organisms.
Whereas most current algorithms avoid exploring the full motif space by using greedy algorithms, our method is unique in the sense that it is exhaustive. MDOS (Wu etal., 2008) only processes promising k-mers and gradually adds degeneracy if this improves the conservation score. Kellis etal. (2003) and Stark etal. (2007) use the mini-motifs approach (van Helden etal., 2000) only processing promising trinucleotide duos before adding degeneracy. Here, every word that occurs in one of the input sequences, including their degenerate variants, is considered as a candidate motif. The only imposed restrictions are a prespecified minimum and maximum length and a maximum number of degenerate IUPAC characters. The advantage of such exhaustive approach is that the method yields globally optimal results. In order to strongly reduce the runtime and avoid excessive memory requirements, the MapReduce programming model (Dean and Ghemawat, 2004) was adopted as a means to take advantage of a parallel, distributed-memory cloud computing environment. By enabling disk I/O to store intermediate results, the current MapReduce implementation overcomes the memory bottleneck in a prototype implementation of this software that relied on the Message Passing Interface (MPI) for parallelization (De Witte etal., 2013).
2 Methods
2.1 Generation of gene families
The orthology relationships between the genes of the four different monocot plant species were inferred using the ‘integrative orthology viewer’ in the PLAZA 2.5 platform (Proost etal., 2009; Van Bel etal., 2012). Homologous (i.e. orthologous and paralogous) genes were grouped in gene families and their promoter sequences 2 kbp upstream from the translation start site were extracted. In its most simple form, a family consists of four orthologous genes: one from each organism. In that case, the phylogenetic tree by Reineke etal. (2011) is used. For gene families that comprise one or more paralogs, gene family-specific phylogenetic trees can be constructed that take into account the specific order in which the duplications and speciation events occurred. For simplicity, we assume that all paralogous gene duplications occurred recently. This is modeled by adding a bifurcation with a branch length of zero to the phylogenetic tree which means that only conservation between different species contributes to the branch length score. Note that besides promoter regions, additional homologous sequences of interest (e.g. intronic regions) could be added to the input dataset.
2.2 Intrafamily step: conservation within a gene family
For all gene families individually, all words with a length between and characters that occur in any of the sequences are exhaustively enumerated and their degree of conservation within that family is quantified. Words are spelled in the IUPAC alphabet or a subset thereof. Up to degenerate (i.e. non-ACGT) characters are allowed per word. The intrafamily phase can operate in alignment-free or alignment-based mode.
In the alignment-free approach, a generalized suffix tree (GST) is constructed (Giegerich etal., 1999) from the promoter sequences and their reverse complements in the gene family. Using Sagot’s Speller algorithm (Marsan and Sagot, 2000), the GST is used to efficiently and exhaustively report all words in the IUPAC alphabet along with the sequences in which they occur. Additional algorithmic details and runtime information are described in Supplementary Methods 1.1.
The alignment-based mode requires a pregenerated multiple sequence alignment (MSA) of the orthologous promoters in a gene family. Dialign-TX (Subramanian etal., 2008) was chosen to create these MSAs in view of good results on a non-coding alignment benchmark (Pollard etal., 2004). For every position in the alignment, a small GST is generated containing only the suffixes of the sequences that start at that position. The same Speller algorithm is run to report all words and the sequences in which they occur at aligned positions, again using the IUPAC alphabet.
For every word, the degree of conservation in each gene family is quantified using the branch length score (BLS). Given the sequences in which the word occurs, the BLS can be calculated by finding the minimum spanning tree that connects these sequences in the phylogenetic tree. The sum of the weights of the horizontal branches in the minimum spanning tree then represents the BLS (Stark etal., 2007). In alignment-based mode, the same motif can occur at multiple aligned positions within a single family; in that case only the highest BLS value is used. Only words for which the BLS exceeds a prespecified threshold T are retained. Such words are said to be conserved within the gene family.
2.3 Interfamily step: genome-wide conservation
The conserved words of all gene families are sorted according to base content and partitioned into permutation groups whose elements are permutations of each other. All words in a permutation group hence have the same length, base content and degeneracy. For example, the words AWTC, WTAC and CAWT belong to the same permutation group.
The number of occurrences for each distinct word within a permutation group is counted. This number corresponds to the number of gene families in which that word is conserved with a and is referred to as the conserved family count F(T). Genome-wide conserved motifs are selected based on the fact that they have a conserved family count F(T) that is (much) higher than the median conserved family count of the member instances of their permutation group. This median value, denoted as (bg = background) represents the expected conserved family count for a word in that permutation group. is approximated by randomly generating a large number (default = 1000) of instances of the permutation group, i.e. random words with the same length and base content and computing the median value for the conserved family count. Note that some of those random instances can have a conserved family count equal to zero.
2.4 MapReduce implementation
The method was implemented using the MapReduce (Dean and Ghemawat, 2004) programming model. The map phase corresponds to the intrafamily phase in which the gene families are processed in parallel by the different mappers. The reduce phase corresponds to the interfamily phase in which the permutations groups are processed in parallel by the different reducers. In between the map and reduce step, the candidate motifs are sorted according to length and base content in order to create the permutation groups.
3 Results and discussion
3.1 BLSSpeller algorithm
The workflow of BLSSpeller is illustrated in Figure 1. The input consists of gene families containing homologous promoter sequences from related species. The algorithm consists of an intrafamily and an interfamily step with a sorting step in between.
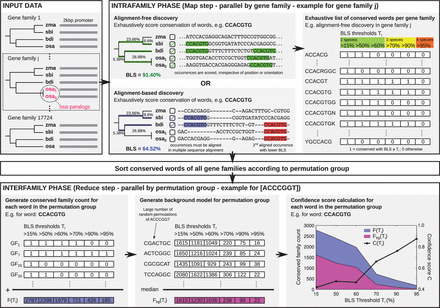
Overview of BLSSpeller. The input consists of homologous promoter sequences grouped into gene families. During the intrafamily phase, conserved words are exhaustively enumerated for each gene family individually. A word is considered to be conserved in a gene family if its branch length score (BLS) exceeds threshold T. Multiple BLS thresholds Ti can be used in a single run. In the alignment-free mode, the BLS of a word is computed irrespective of its orientation or relative position within the promoter sequences. Alternatively, in the alignment-based mode, words must appear aligned in the multiple sequence alignment. During the sorting phase, conserved words of all gene families are sorted according to permutation group, i.e. words with the same length and base content are grouped together. In the interfamily phase, permutation groups are handled individually. First, for each word, the conserved family count , i.e. the number of gene families in which the word is conserved with BLS , is established for all BLS thresholds Ti. Next, a background model is created by selecting the median value of the conserved family count of a large number of randomly generated instances of the permutation group, again for each threshold Ti. Finally, a confidence score is computed for each Ti. Words for which and for any threshold Ti are considered to be genome-wide conserved motifs and are retained
3.1.1 Intrafamily step
In the intrafamily step, for each gene family individually, all words with a length between and characters that occur in the promoter sequences of that gene family are exhaustively enumerated. Words are spelled in the restricted IUPAC alphabet that consists of 11 characters: 4 base pairs (ACGT), 6 twofold-degenerate characters (RYSWKM) and the ‘any’ character (N). A maximum of degenerate characters are allowed per candidate motif. The degree of conservation of a word within the gene family is scored using the branch length score (BLS). The intrafamily step can operate in either alignment-free (AF) or alignment-based (AB) mode. In case of AF discovery, the BLS of a word is scored irrespective of its orientation or relative position within the promoter sequences. AB discovery adds the constraint that the words must be aligned in the multiple sequence alignment of the promoter sequences. Words for which the BLS exceeds threshold T are considered to be conserved within the gene family and retained for further processing. Six BLS thresholds Ti (i.e. 15, 50, 60, 70, 90 and 95%) were used in this study. At the end of this phase an exhaustive list of conserved words has been generated for each gene family individually.
3.1.2 Interfamily step
Using the data from the intrafamily step, for each word, the conserved family count, i.e. the number of gene families in which the word is conserved with a BLS , is counted for each BLS threshold Ti. Next, a confidence score , adopted from Stark etal. (2007), is established for each candidate motif (see Section 2). Two thresholds apply: motifs are only retained when and for any of the BLS thresholds Ti. Here, represents a threshold on the conserved family count and is used to eliminate words that are conserved in only few gene families and hence typically do not correspond to TF binding sites. Additionally, ensures that the candidate motif is conserved in a much higher number of gene families than what is expected for such a word (i.e. a word with the same length, base composition and degeneracy) and can hence be considered a potentially functional element. Motifs that satisfy both thresholds are considered to be genome-wide conserved motifs.
Note that the branch length score thresholds Ti on the one hand and conserved family count threshold and confidence score threshold on the other hand are independent. The former provides information about the degree of conservation within a single gene family whereas the latter are indicative of the degree of genome-wide conservation. Certain motifs only show up as being genome-wide conserved for high BLS thresholds. This is typically the case for short and/or highly degenerate motifs, where also permutations of that motif are conserved with a moderate BLS in a rather large number of families, resulting in a low confidence score C. Conversely, a lower BLS threshold allows for the detection of longer motifs with genome-wide conservation in only a subset of the species. Using only a single BLS threshold would therefore limit the sensitivity of the method.
3.2 Exhaustive motif discovery in four monocot species
BLSSpeller was applied to four monocot species: O.sativa ssp.indica (osa), B.distachyon (bdi), S.bicolor (sbi) and Z.mays (zma). Based on conserved gene content and genome organization, these grass species are considered to be a single genetic system (Benntzin and Freeling, 1993), making a comparative motif discovery approach feasible. The dataset consists of 17 724 gene families each containing four orthologous genes (one from each organism). Additionally, 10 636 paralogs are taken into account. Hence, a total of 163 064 regulatory sequences (forward and reverse strands) with a length of 2 kbp each, were analyzed.
BLSSpeller was run on this dataset using both the alignment-free (AF) and the alignment-based (AB) discovery mode on the Amazon Web Services (Elastic MapReduce) cloud infrastructure using 20 nodes of the type m1.xlarge. On every node, 7 map tasks and 2 reduce tasks were run in parallel. The computational requirements are listed in Supplementary Results 2.1. Based on the Amazon pricing of 2014, the financial cost for performing these simulations amounted to 1080$ and 278$ for the AF and AB cases, respectively.
After the intrafamily step and using the AF discovery mode, an aggregated number of 537 billion words were found with a BLS (i.e. conservation in at least two species) over all 17 724 gene families. Note that these words are not necessarily unique as the same word can be conserved in multiple gene families. Using the AB discovery mode, only 82 billion words were found with a BLS . This is because the AB discovery mode imposes the additional constraint that words should appear aligned in the multiple sequence alignment. After the interfamily step and using and , the number of genome-wide conserved motifs amounted to 6.62 and 6.26 billion unique motifs, for the AF and AB discovery mode respectively.
The reason why the number of motifs is high is twofold. First, very relaxed thresholds and were used. It is computationally cheap to further filter this list using more stringent (and biologically meaningful) thresholds (see below). A second reason is the exhaustive, word-based nature of BLSSpeller. If a word is found to be genome-wide conserved, a large number of redundant, highly similar (e.g. slightly more degenerate) variants of that word may also appear in the final output of the method.
3.3 Estimation of the false discovery rate
The output of BLSSpeller consists of a list of motifs, along with the conserved family count and conservation score for the six different BLS thresholds Ti. This list was filtered using more stringent thresholds for (i.e. 1, 10 and 20) and (i.e. 0.5, 0.7 and 0.9). Additionally, the list can be filtered by considering only a (stricter) subset of the BLS thresholds Ti (i.e. all six thresholds , three thresholds corresponding to conservation in at least three species, a single threshold T6 corresponding to conservation in all four species). The number of genome-wide conserved motifs for all 27 parameter combinations is shown in Figure 2 for both AF and AB discovery. Clearly, each of the parameters has a strong influence on the final number of motifs in both the AF and AB discovery.
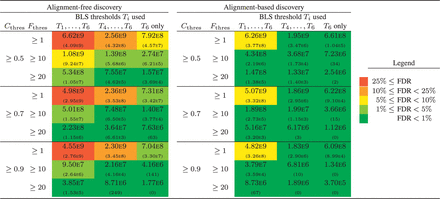
Number of genome-wide conserved motifs for both alignment-based and alignment-free discovery for different values of and and different subsets of the six BLS thresholds Ti ( and ). Top number: real Monocot dataset; bottom number between brackets: random dataset (zeroth-order Markov model). The colors represent the false discovery rate (see legend)
In order to assess the specificity of the method for the different parameter combinations, we estimate the false discovery rate (FDR) in an empirical fashion by running BLSSpeller on a random dataset generated using a zeroth-order Markov model (preservation of mononucleotide frequencies) as provided by RSAT (Thomas-Chollier etal., 2008). A more detailed version of Figure 2 is available as Supplementary Figure S4. Additional discussion of the limitations of the FDR analysis, higher-order Markov models and FDR analysis as a function of motif length and degeneracy is provided in Supplementary Results 2.2.
A number of observations can be made. First, for comparable parameter settings, AB discovery has a lower FDR compared to AF discovery. The multiple sequence alignment method increases the specificity for AB discovery as relatively few words will be aligned in random data purely by chance. Second, low values of result in a poor FDR. The reason for this is that in such case, the output consists of a large number of words that are conserved in only a single gene family. If these words are long and/or have low degeneracy, most random permutations of that word will not be conserved in any gene family, resulting in a confidence score . We therefore recommend to impose a certain threshold on the conserved family count. As functional transcription factors typically target multiple genes, this appears to be a biologically reasonable approach. Third, a reasonable threshold on the confidence score should be applied. Applying this threshold filters words for which their random permutations are conserved in a comparable number of gene families. This comprises low-complexity motifs and/or highly degenerate motifs. Finally, a more stringent definition of conservation results in an improved FDR. This can be obtained by imposing higher BLS thresholds Ti.
Even though there is a clear correlation between each of the parameters and the FDR, the exact FDR is hard to predict up front and likely also depends on the dataset that is used. We therefore recommend to run BLSSpeller with relaxed parameter settings on both real and random data, and to filter this output using more stringent parameters until a reasonable FDR is obtained.
For reasonably stringent parameter settings where the FDR , the AF discovery mode reports 3.1–6.8 times more motifs compared to the AB discovery. At first glance, this may seem to be a trivial consequence of the relaxed definition of conservation in the AF methodology. Indeed, a word that is found to be conserved in a gene family with BLS using the AB discovery will also be conserved in the AF method. Therefore, for each word. However, in order to establish the confidence score C(T), the conserved family count F(T) is compared to the corresponding median value of the background distribution (see Section 2). As is also computed using the relaxed, alignment-free definition of conservation, it holds that . Therefore, there is no reason to assume a priori that the AF mode will pick up more motifs than its AB counterpart, as can indeed be observed in Figure 2 for a few parameter combinations, e.g. and BLS thresholds . The reason that we do find more genome-wide conserved motifs for most parameter combinations (including those with good FDR) is because we found a significant number of known motif instances to be misaligned in this relatively highly diverged Monocot dataset. This is exemplified in Section 3.5.
3.4 Motif instance predictions correlate with experimental cis-regulatory datasets
The genome-wide conserved motifs discovered by BLSSpeller are highly redundant. High-scoring, motifs (AF discovery; BLS ; 38 462 976 motifs in total) were mapped back to the promoter sequences and were found to cluster around specific genomic regions (see Supplementary Figs. S8 and Supplementary Data). Certain loci are covered by thousands of highly similar motif variants. Nevertheless, the high-scoring motifs delineate distinct conserved genomic intervals on the promoter sequences. For these conserved regions, we investigated the accessibility for transcription factor binding in the promoter sequences of rice genes. DNase I hypersensitive sites are associated with regions of open chromatin where the DNA is accessible and as such provide a global perspective on possible protein-binding to the genome. Such regions were recently characterized by Zhang etal. (2012). We performed overlap analysis between conserved genomic regions (as determined by BLSSpeller) and open chromatin regions (see Supplementary Methods 1.2). We found a significant enrichment (3.005 fold) of conserved regions for open chromatin regions (P-value < 0.001) (see Table 1). For a stricter subset of motifs (AF discovery; BLS ; 1 769 963 motifs in total), the fold enrichment increased to 3.796.
Overlap between conserved genomic regions as identified by BLSSpeller and experimentally profiled open chromatin regions in rice and transcription factor binding sites inferred through protein-binding microarrays in rice and maize
| Overlap with experimentally profiled open chromatin regions (OCR) in O.sativa | |||||
|---|---|---|---|---|---|
| BLSSpeller | No. of conserved | No. of OCR regions | No. of conserved regions | No. of rand. conserved regions | enrichment |
| thresholds | regions | within OCR regions | within OCR regions | fold | |
| BLS 15%, 0.9, 20 | 754 205 | 77 247 | 121 026 | 40 277 | 3.005 |
| BLS 95%, 0.9, 20 | 464 229 | 77 247 | 98 681 | 25 996 | 3.796 |
| Overlap with experimentally profiled open chromatin regions (OCR) in O.sativa | |||||
|---|---|---|---|---|---|
| BLSSpeller | No. of conserved | No. of OCR regions | No. of conserved regions | No. of rand. conserved regions | enrichment |
| thresholds | regions | within OCR regions | within OCR regions | fold | |
| BLS 15%, 0.9, 20 | 754 205 | 77 247 | 121 026 | 40 277 | 3.005 |
| BLS 95%, 0.9, 20 | 464 229 | 77 247 | 98 681 | 25 996 | 3.796 |
Overlap with experimentally profiled TF binding sites (TBS) in O.sativa | |||||
|---|---|---|---|---|---|
| BLSSpeller | No. of conserved | No. of TBS regions | No. of TBS regions within | No. of TBS regions within | enrichment |
| thresholds | regions | conserved regions | rand. conserved regions | fold | |
| BLS 15%, 0.9, 20 | 754 205 | 442 506 | 159 542 | 42 522 | 3.752 |
| BLS 95%, 0.9, 20 | 464 229 | 442 506 | 37 093 | 5 689 | 6.520 |
Overlap with experimentally profiled TF binding sites (TBS) in O.sativa | |||||
|---|---|---|---|---|---|
| BLSSpeller | No. of conserved | No. of TBS regions | No. of TBS regions within | No. of TBS regions within | enrichment |
| thresholds | regions | conserved regions | rand. conserved regions | fold | |
| BLS 15%, 0.9, 20 | 754 205 | 442 506 | 159 542 | 42 522 | 3.752 |
| BLS 95%, 0.9, 20 | 464 229 | 442 506 | 37 093 | 5 689 | 6.520 |
Overlap with experimentally profiled TF binding sites (TBS) in Z.mays | |||||
|---|---|---|---|---|---|
| BLSSpeller | No. of conserved | No. of TBS regions | No. of TBS regions within | No. of TBS regions within | enrichment |
| thresholds | regions | conserved regions | rand. conserved regions | fold | |
| BLS 15%, 0.9, 20 | 828 400 | 482 317 | 156 929 | 66 564 | 2.358 |
| BLS 95%, 0.9, 20 | 454 221 | 482 317 | 35 710 | 10 755 | 3.320 |
Overlap with experimentally profiled TF binding sites (TBS) in Z.mays | |||||
|---|---|---|---|---|---|
| BLSSpeller | No. of conserved | No. of TBS regions | No. of TBS regions within | No. of TBS regions within | enrichment |
| thresholds | regions | conserved regions | rand. conserved regions | fold | |
| BLS 15%, 0.9, 20 | 828 400 | 482 317 | 156 929 | 66 564 | 2.358 |
| BLS 95%, 0.9, 20 | 454 221 | 482 317 | 35 710 | 10 755 | 3.320 |
Regions are required to fully overlap in order to be scored.
Overlap between conserved genomic regions as identified by BLSSpeller and experimentally profiled open chromatin regions in rice and transcription factor binding sites inferred through protein-binding microarrays in rice and maize
| Overlap with experimentally profiled open chromatin regions (OCR) in O.sativa | |||||
|---|---|---|---|---|---|
| BLSSpeller | No. of conserved | No. of OCR regions | No. of conserved regions | No. of rand. conserved regions | enrichment |
| thresholds | regions | within OCR regions | within OCR regions | fold | |
| BLS 15%, 0.9, 20 | 754 205 | 77 247 | 121 026 | 40 277 | 3.005 |
| BLS 95%, 0.9, 20 | 464 229 | 77 247 | 98 681 | 25 996 | 3.796 |
| Overlap with experimentally profiled open chromatin regions (OCR) in O.sativa | |||||
|---|---|---|---|---|---|
| BLSSpeller | No. of conserved | No. of OCR regions | No. of conserved regions | No. of rand. conserved regions | enrichment |
| thresholds | regions | within OCR regions | within OCR regions | fold | |
| BLS 15%, 0.9, 20 | 754 205 | 77 247 | 121 026 | 40 277 | 3.005 |
| BLS 95%, 0.9, 20 | 464 229 | 77 247 | 98 681 | 25 996 | 3.796 |
Overlap with experimentally profiled TF binding sites (TBS) in O.sativa | |||||
|---|---|---|---|---|---|
| BLSSpeller | No. of conserved | No. of TBS regions | No. of TBS regions within | No. of TBS regions within | enrichment |
| thresholds | regions | conserved regions | rand. conserved regions | fold | |
| BLS 15%, 0.9, 20 | 754 205 | 442 506 | 159 542 | 42 522 | 3.752 |
| BLS 95%, 0.9, 20 | 464 229 | 442 506 | 37 093 | 5 689 | 6.520 |
Overlap with experimentally profiled TF binding sites (TBS) in O.sativa | |||||
|---|---|---|---|---|---|
| BLSSpeller | No. of conserved | No. of TBS regions | No. of TBS regions within | No. of TBS regions within | enrichment |
| thresholds | regions | conserved regions | rand. conserved regions | fold | |
| BLS 15%, 0.9, 20 | 754 205 | 442 506 | 159 542 | 42 522 | 3.752 |
| BLS 95%, 0.9, 20 | 464 229 | 442 506 | 37 093 | 5 689 | 6.520 |
Overlap with experimentally profiled TF binding sites (TBS) in Z.mays | |||||
|---|---|---|---|---|---|
| BLSSpeller | No. of conserved | No. of TBS regions | No. of TBS regions within | No. of TBS regions within | enrichment |
| thresholds | regions | conserved regions | rand. conserved regions | fold | |
| BLS 15%, 0.9, 20 | 828 400 | 482 317 | 156 929 | 66 564 | 2.358 |
| BLS 95%, 0.9, 20 | 454 221 | 482 317 | 35 710 | 10 755 | 3.320 |
Overlap with experimentally profiled TF binding sites (TBS) in Z.mays | |||||
|---|---|---|---|---|---|
| BLSSpeller | No. of conserved | No. of TBS regions | No. of TBS regions within | No. of TBS regions within | enrichment |
| thresholds | regions | conserved regions | rand. conserved regions | fold | |
| BLS 15%, 0.9, 20 | 828 400 | 482 317 | 156 929 | 66 564 | 2.358 |
| BLS 95%, 0.9, 20 | 454 221 | 482 317 | 35 710 | 10 755 | 3.320 |
Regions are required to fully overlap in order to be scored.
Additionally, we investigated the enrichment of TF binding sites determined in vitro (Weirauch etal., 2014) towards conserved genomic regions in rice and maize. Transcription factor DNA binding specificities are the primary mechanism by which transcription factors recognize genomic features and regulate genes. Recently, a dataset containing a large number of these binding specificities was generated using protein-binding microarrays (PBM) (Weirauch etal., 2014). From this database, PWMs were downloaded for 481 TFs in rice and for 615 TFs in maize. These were mapped onto the respective rice and maize promoters and overlap analysis was performed (see Supplementary Methods 1.2). In rice, of the 754 205 constrained genomic regions (BLS ), 159 542 contain a PBM-based TF binding site, leading to 3.752 fold enrichment (P-value < 0.001). Again, for the stricter subset of conserved motifs (BLS ), fold enrichment increased to 6.520. Maize showed a fold enrichment of 2.358 and 3.320 (P-value < 0.001) respectively. Overall, these analyses revealed that a large part of the conserved non-coding sequences can be accessed by DNA binding proteins and as such can act as functional transcription factor binding sites, and that these conserved non-coding sequences show enrichment for the binding sites of a large number of TFs inferred using PBMs.
3.5 Conservation of the ga2ox1-like KN1 binding site
KNOTTED1 (KN1) transcription factors are involved in the establishment and maintenance of plant meristems and are thought to be conserved among the family of grasses (Bolduc and Hake, 2009). Bolduc etal. (2012) profiled KN1 binding sites in Z.mays using ChIP-seq experiments. The overlapping loci in two samples of immature ears were retained and assigned to the nearest gene within a range of 10 kbp. The ChIP-Seq peaks were found to be mainly situated in the 5′ en 3′ regions extending from the gene but also occur in introns and exons. Thus, a set of candidate KN1-regulated maize genes were identified. For approximately 7% of these genes, a binding site reminiscent of the intronic KN1 binding site in ga2ox1, was identified. For these so-called ga2ox1-like KN1 binding sites, a Position Weight Matrix (PWM) was derived by Bolduc etal. (2012). Translated to the IUPAC alphabet, this PWM corresponds to TGAYNGAYDGAY.
We investigate whether BLSSpeller is able to discover the ga2ox1-like KN1 motifs and binding sites through a comparative study of the four monocot species. From the BLSSpeller output, all genome-wide conserved motifs of length 12 that match the ga2ox1-like KN1 PWM identified by Bolduc etal. (2012) were retained. Using alignment-free discovery, and using and (FDR , see Fig. 2), 51 genome-wide conserved motif variants are identified. In total, these motifs are conserved in 165 gene families with a BLS (i.e. conservation in at least two species). From the 51 identified motif variants, only 19 are required to explain the conservation in all 165 gene families. These essential motifs are listed in Table 2 along with their respective metrics. In turn, these gene families contain 213 maize genes in total, 51 of which were also identified in Bolduc etal. (2012). These results were compared to those obtained by Fastcompare (Elemento and Tavazoie, 2005; see Supplementary Results 2.3), a method that also performs motif discovery in an alignment-free and exhaustive manner. However, Fastcompare is limited to the exact ACGT alphabet and pairwise species comparisons. Because of these limitations, Fastcompare could identify only 36 maize gene targets, 10 of which were also identified by Bolduc etal. (2012).
List of genome-wide conserved ga2ox1-like KN1 motif variants identified by BLSSpeller using both AF and AB discovery
Alignment-free discovery | Alignment-based discovery | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| KN1 motif variant | KN1 motif variant | ||||||||
| TGATNGATKGAY | 59 | 0.93 | 75 | 24 | TGATNGAYGGAY | 11 | 0.91 | 10 | 3 |
| TGATNGAYKGAT | 59 | 0.93 | 74 | 20 | TGATNGATKGAY | 11 | 0.82 | 11 | 3 |
| TGAYNGATKGAT | 54 | 0.93 | 68 | 21 | TGAYNGACKGAC | 10 | 0.90 | 11 | 3 |
| TGATNGAYWGAT | 40 | 0.88 | 50 | 11 | TGAYGGAYGGAY | 9 | 1.00 | 9 | 3 |
| TGAYNGAYTGAT | 36 | 0.89 | 48 | 11 | TGATNGAYRGAT | 9 | 0.89 | 10 | 3 |
| TGAYTGAYTGAY | 33 | 0.97 | 42 | 9 | TGAYNGAYTGAC | 8 | 0.88 | 9 | 2 |
| TGATNGAYTGAY | 32 | 0.88 | 40 | 7 | TGACNGAYTGAY | 8 | 0.88 | 10 | 3 |
| TGAYNGATWGAT | 31 | 0.84 | 42 | 12 | TGACNGACWGAY | 7 | 0.86 | 7 | 2 |
| TGATNGATWGAY | 30 | 0.83 | 36 | 9 | TGACAGAYRGAY | 3 | 1.00 | 4 | 0 |
| TGATNGATRGAY | 29 | 0.86 | 39 | 9 | |||||
| TGAYNGATRGAT | 27 | 0.85 | 37 | 9 | |||||
| TGATNGAYRGAT | 26 | 0.85 | 35 | 8 | |||||
| TGAYNGATTGAY | 25 | 0.84 | 34 | 7 | |||||
| TGAYNGATGGAY | 24 | 0.88 | 35 | 9 | |||||
| TGATNGAYGGAY | 24 | 0.88 | 31 | 8 | |||||
| TGAYTGAYWGAT | 22 | 0.91 | 27 | 6 | |||||
| TGAYNGACTGAY | 22 | 0.91 | 28 | 9 | |||||
| TGAYNGAYTGAC | 21 | 0.90 | 27 | 8 | |||||
| TGAYNGACKGAC | 20 | 0.90 | 25 | 10 | |||||
| Union (all variants) | 165 | – | 213 | 51 | Union (all variants) | 37 | – | 41 | 10 |
Alignment-free discovery | Alignment-based discovery | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| KN1 motif variant | KN1 motif variant | ||||||||
| TGATNGATKGAY | 59 | 0.93 | 75 | 24 | TGATNGAYGGAY | 11 | 0.91 | 10 | 3 |
| TGATNGAYKGAT | 59 | 0.93 | 74 | 20 | TGATNGATKGAY | 11 | 0.82 | 11 | 3 |
| TGAYNGATKGAT | 54 | 0.93 | 68 | 21 | TGAYNGACKGAC | 10 | 0.90 | 11 | 3 |
| TGATNGAYWGAT | 40 | 0.88 | 50 | 11 | TGAYGGAYGGAY | 9 | 1.00 | 9 | 3 |
| TGAYNGAYTGAT | 36 | 0.89 | 48 | 11 | TGATNGAYRGAT | 9 | 0.89 | 10 | 3 |
| TGAYTGAYTGAY | 33 | 0.97 | 42 | 9 | TGAYNGAYTGAC | 8 | 0.88 | 9 | 2 |
| TGATNGAYTGAY | 32 | 0.88 | 40 | 7 | TGACNGAYTGAY | 8 | 0.88 | 10 | 3 |
| TGAYNGATWGAT | 31 | 0.84 | 42 | 12 | TGACNGACWGAY | 7 | 0.86 | 7 | 2 |
| TGATNGATWGAY | 30 | 0.83 | 36 | 9 | TGACAGAYRGAY | 3 | 1.00 | 4 | 0 |
| TGATNGATRGAY | 29 | 0.86 | 39 | 9 | |||||
| TGAYNGATRGAT | 27 | 0.85 | 37 | 9 | |||||
| TGATNGAYRGAT | 26 | 0.85 | 35 | 8 | |||||
| TGAYNGATTGAY | 25 | 0.84 | 34 | 7 | |||||
| TGAYNGATGGAY | 24 | 0.88 | 35 | 9 | |||||
| TGATNGAYGGAY | 24 | 0.88 | 31 | 8 | |||||
| TGAYTGAYWGAT | 22 | 0.91 | 27 | 6 | |||||
| TGAYNGACTGAY | 22 | 0.91 | 28 | 9 | |||||
| TGAYNGAYTGAC | 21 | 0.90 | 27 | 8 | |||||
| TGAYNGACKGAC | 20 | 0.90 | 25 | 10 | |||||
| Union (all variants) | 165 | – | 213 | 51 | Union (all variants) | 37 | – | 41 | 10 |
denotes the number of gene families in which the motif is conserved with while denotes the corresponding confidence score. denotes the number of maize genes contained in the gene families while denotes the intersection with experimentally profiled maize genes.
List of genome-wide conserved ga2ox1-like KN1 motif variants identified by BLSSpeller using both AF and AB discovery
Alignment-free discovery | Alignment-based discovery | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| KN1 motif variant | KN1 motif variant | ||||||||
| TGATNGATKGAY | 59 | 0.93 | 75 | 24 | TGATNGAYGGAY | 11 | 0.91 | 10 | 3 |
| TGATNGAYKGAT | 59 | 0.93 | 74 | 20 | TGATNGATKGAY | 11 | 0.82 | 11 | 3 |
| TGAYNGATKGAT | 54 | 0.93 | 68 | 21 | TGAYNGACKGAC | 10 | 0.90 | 11 | 3 |
| TGATNGAYWGAT | 40 | 0.88 | 50 | 11 | TGAYGGAYGGAY | 9 | 1.00 | 9 | 3 |
| TGAYNGAYTGAT | 36 | 0.89 | 48 | 11 | TGATNGAYRGAT | 9 | 0.89 | 10 | 3 |
| TGAYTGAYTGAY | 33 | 0.97 | 42 | 9 | TGAYNGAYTGAC | 8 | 0.88 | 9 | 2 |
| TGATNGAYTGAY | 32 | 0.88 | 40 | 7 | TGACNGAYTGAY | 8 | 0.88 | 10 | 3 |
| TGAYNGATWGAT | 31 | 0.84 | 42 | 12 | TGACNGACWGAY | 7 | 0.86 | 7 | 2 |
| TGATNGATWGAY | 30 | 0.83 | 36 | 9 | TGACAGAYRGAY | 3 | 1.00 | 4 | 0 |
| TGATNGATRGAY | 29 | 0.86 | 39 | 9 | |||||
| TGAYNGATRGAT | 27 | 0.85 | 37 | 9 | |||||
| TGATNGAYRGAT | 26 | 0.85 | 35 | 8 | |||||
| TGAYNGATTGAY | 25 | 0.84 | 34 | 7 | |||||
| TGAYNGATGGAY | 24 | 0.88 | 35 | 9 | |||||
| TGATNGAYGGAY | 24 | 0.88 | 31 | 8 | |||||
| TGAYTGAYWGAT | 22 | 0.91 | 27 | 6 | |||||
| TGAYNGACTGAY | 22 | 0.91 | 28 | 9 | |||||
| TGAYNGAYTGAC | 21 | 0.90 | 27 | 8 | |||||
| TGAYNGACKGAC | 20 | 0.90 | 25 | 10 | |||||
| Union (all variants) | 165 | – | 213 | 51 | Union (all variants) | 37 | – | 41 | 10 |
Alignment-free discovery | Alignment-based discovery | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| KN1 motif variant | KN1 motif variant | ||||||||
| TGATNGATKGAY | 59 | 0.93 | 75 | 24 | TGATNGAYGGAY | 11 | 0.91 | 10 | 3 |
| TGATNGAYKGAT | 59 | 0.93 | 74 | 20 | TGATNGATKGAY | 11 | 0.82 | 11 | 3 |
| TGAYNGATKGAT | 54 | 0.93 | 68 | 21 | TGAYNGACKGAC | 10 | 0.90 | 11 | 3 |
| TGATNGAYWGAT | 40 | 0.88 | 50 | 11 | TGAYGGAYGGAY | 9 | 1.00 | 9 | 3 |
| TGAYNGAYTGAT | 36 | 0.89 | 48 | 11 | TGATNGAYRGAT | 9 | 0.89 | 10 | 3 |
| TGAYTGAYTGAY | 33 | 0.97 | 42 | 9 | TGAYNGAYTGAC | 8 | 0.88 | 9 | 2 |
| TGATNGAYTGAY | 32 | 0.88 | 40 | 7 | TGACNGAYTGAY | 8 | 0.88 | 10 | 3 |
| TGAYNGATWGAT | 31 | 0.84 | 42 | 12 | TGACNGACWGAY | 7 | 0.86 | 7 | 2 |
| TGATNGATWGAY | 30 | 0.83 | 36 | 9 | TGACAGAYRGAY | 3 | 1.00 | 4 | 0 |
| TGATNGATRGAY | 29 | 0.86 | 39 | 9 | |||||
| TGAYNGATRGAT | 27 | 0.85 | 37 | 9 | |||||
| TGATNGAYRGAT | 26 | 0.85 | 35 | 8 | |||||
| TGAYNGATTGAY | 25 | 0.84 | 34 | 7 | |||||
| TGAYNGATGGAY | 24 | 0.88 | 35 | 9 | |||||
| TGATNGAYGGAY | 24 | 0.88 | 31 | 8 | |||||
| TGAYTGAYWGAT | 22 | 0.91 | 27 | 6 | |||||
| TGAYNGACTGAY | 22 | 0.91 | 28 | 9 | |||||
| TGAYNGAYTGAC | 21 | 0.90 | 27 | 8 | |||||
| TGAYNGACKGAC | 20 | 0.90 | 25 | 10 | |||||
| Union (all variants) | 165 | – | 213 | 51 | Union (all variants) | 37 | – | 41 | 10 |
denotes the number of gene families in which the motif is conserved with while denotes the corresponding confidence score. denotes the number of maize genes contained in the gene families while denotes the intersection with experimentally profiled maize genes.
Similarly, using BLSSpeller’s alignment-based discovery mode, conservation with a BLS is observed in only 37 gene families, even with very relaxed thresholds ( and ) (FDR ). The nine essential motif variants required to explain this conservation are listed in Table 2. The 37 gene families contain 41 maize genes, 10 of which are also reported in Bolduc etal. (2012). Inspection of the promoter sequence alignments of the gene families reveals that the ga2ox1-like KN1 variants are often not aligned, either because the motif instances in the different species are located at entirely different positions in the promoter sequences or because they appear on different strands (see Supplementary Table 3). Therefore, alignment-based motif discovery approaches such as BLSSpeller in AB mode or the ‘mini motifs’ approach as used by Stark etal. (2007) suffer from reduced sensitivity on diverged datasets.
4 Conclusion
A novel phylogenetic footprinting approach was developed for the sensitive discovery of conserved cis-regulatory elements even in diverged sequences. Using IUPAC strings as motif model and using the MapReduce programming model to enable distributed computing, it was shown that it is feasible to compute all genome-wide conserved words in a large dataset in an exhaustive manner. For a given false discovery rate, it was demonstrated that an alignment-free approach detects more conserved words than an alignment-based approach. Even though millions of genome-wide conserved motifs were identified by our method, mapping of these motifs to the promoter sequences results in constrained conserved genomic regions. It was shown that these conserved regions were significantly enriched for experimentally profiled open chromatin regions in rice and for TF binding sites inferred through protein-binding microarrays in rice and maize. Finally, it was shown that the alignment-free approach shows an improved recovery of the ga2ox1-like KN1 binding site, compared to the alignment-based approach or competing methods.
Acknowledgements
We acknowledge the support of Ghent University (Multidisciplinary Research Partnership ‘Bioinformatics: From Nucleotides to Networks’) and Dries Vaneechoutte, Kenneth Hoste, Ewan Higgs and Stijn De Weirdt for technical assistance. Part of the computational resources (Stevin Supercomputer Infrastructure) and services used in this work were provided by the VSC (Flemish Supercomputer Center), funded by Ghent University, the Hercules Foundation and the Flemish Government department EWI.
Funding
J.V.D.V. is supported by the Institute for the Promotion of Innovation by Science and Technology in Flanders (IWT-Vlaanderen).
Conflict of Interest: none declared.
References
Author notes
Associate Editor: Alfonso Valencia
†The authors wish it to be known that, in their opinion, the first two authors should be regarded as Joint First Authors.
‡The authors wish it to be known that, in their opinion, the last two authors should be regarded as Joint Last Authors.


{kind=link}
{kind=link}