





Abstract
Because they share several morphological and ecological characters, the North American sparrow (Emberizidae) genera Ammodramus, Passerculus, and Xenospiza have historically been considered members of a well-defined “grassland” sparrow assemblage. Relationships among the 11 members of this group have been the subject of much taxonomic debate, yet no comprehensive molecular assessment of relationships has been done. We investigated these relationships using mitochondrial DNA sequence data that included complete cytochrome-b and ND2 genes. Phylogenetic reconstructions derived via parsimony, likelihood, and Bayesian methods were congruent. The grassland sparrows, as presently configured, are polyphyletic. Pooecetes gramineus, Amphispiza belli (but not A. quinquestriata and A. bilineata), Oriturus superciliosus, and all three species of Melospiza are included in a reconfigured clade, whereas the traditional forms of Ammodramus savannarum, humeralis, and aurifons are placed well outside of these. Within the clade of interest, Ammodramus remains polyphyletic, with leconteii, maritimus, nelsoni, and caudacutus forming a well-resolved clade apart from henslowii and bairdii. The latter are in another strongly supported clade that also includes Passerculus and a Xenospiza-Melozpiza sister pairing. Pooecetes, Amphispiza (belli), and Oriturus represent early lineages in this clade that today have no close living relatives. The polyphyly of the genus Ammodramus is likely the result of morphological convergence attributable to similar adaptive responses to the occupation of similar habitats. In general, the morphological and ecological factors that have defined the grassland sparrows are poor indicators of relatedness. Taxonomic revisions are suggested.
Evaluación Molecular del Clado de Gorriones de Pastizales de Norte América
Resumen
Debido a que comparten varios rasgos morfológicos y ecológicos, los géneros norteamericanos Ammodramus, Passerculus y Xenospiza (Emberizidae) se han considerado históricamente como miembros de un ensamblaje bien definido de gorriones de pastizales. Las relaciones entre los 11 miembros de este grupo han sido objeto de muchos debates taxonómicos, pero no se ha hecho ninguna evaluación exhaustiva de sus relaciones con base en datos moleculares. Investigamos las relaciones entre estas aves utlizando datos de secuencias mitocondriales completas de los genes citocromo b y ND2. Las reconstrucciones filogenéticas basadas en métodos de parsimonia, verosimilitud y Bayesianos fueron congruentes. El grupo de los gorriones de pastizales tal como está configurado actualmente es polifilético. Pooecetes gramineus, Amphispiza belli (pero no A. quinquestriata ni A. bilineata), Oriturus superciliosus y las tres especies de Melospiza están incluidas en un clado reconfigurado, mientras que las formas tradicionales de Ammodramus savannarum, humeralis y aurifons se ubican bien afuera de las demás. Dentro del clado de interés, Ammodramus es aún polifilético, con leconteii, maritimus, nelsoni y caudacutus formando un clado bien resuelto, aparte de henslowii y bairdii. Estos dos últimos taxones están en otro clado fuertemente respaldado que también incluye a Passerculus y la pareja de taxones hermanos Xenospiza-Melozpiza. Pooecetes, Amphispiza (belli) y Oriturus representan linajes que se separaron tempranamente dentro de este clado y en la actualidad no tienen parientes cercanos. La polifilia del género Ammodramus probablemente se debe a convergencia atribuible a respuestas adaptativas similares a la ocupación de ambientes similares. En general, los factores morfológicos y ecológicos que han definido a los gorriones de pastizales son malos indicadores de las relaciones de parentesco. Se sugieren revisiones taxonómicas.
Many of the temperate-zone sparrows of North America have been divided into one of two traditional groups, the “grassland” and the “brushland” nesting sparrows (e.g., Paynter 1964, Dickerman et al. 1967, Robins and Schnell 1971). The former comprises the modern genera Ammodramus, Passerculus, and Xenospiza, whereas the latter includes Melospiza, Passerella, Zonotrichia, and Junco. Although formal phylogenetic analyses were lacking historically, members in each of these complexes were assumed to be closely related through “recency of common ancestry” (Robins and Schnell 1971). Several additional North American genera not assigned to one of these core “clades” include Aimophila, Oriturus, Spizella, Pooecetes, Chondestes, and Amphispiza. These taxa are typically listed before (American Ornithologists' Union [AOU] 1998) or after (Paynter and Storer 1970, Sibley and Monroe 1990) the grassland and brushland assemblages in modern linear taxonomies, and their systematic affinities remain poorly understood.
Although the “grassland” sparrow clade appears to be well circumscribed, little agreement exists regarding relationships among its constituents (the numerous taxonomic revisions attempted for this group of sparrows are reviewed by Murray [1968]). The genus Ammodramus, in particular, has a tumultuous taxonomic history, with its nine current members historically divided into anywhere from one (AOU 1998) to four (Ridgway 1901) to seven (Oberholser 1917) different genera (Table 1). Early taxonomies also listed Passerculus as a member of Ammodramus (e.g., AOU 1886, Chapman 1895) until it was raised to generic status by Ridgway (1901), where it has remained (but see Paynter and Storer 1970). Xenospiza, the montane Mexican endemic first described by Bangs (1931), is a relatively recent addition to the group. Although most workers place it among the grassland sparrow assemblage (Dickerman et al. 1967, Robins and Schnell 1971; but see Pitelka 1947), its “exact position within this group is less obvious” (Dickerman et al. 1967). The group, as currently configured, comprises 11 recognized (i.e., “biological”) species. It has been the focus of some formal systematic analyses, including a thorough phenetic analysis by Robins and Schnell (1971), who subdivided the complex into two clades, Ammodramus (the grassland sparrows, including the currently recognized forms sandwichensis, aurifrons, humeralis, savannarum, and bairdii) and Ammospiza (the “marshland” sparrows, which included the modern leconteii, caudacuta, nelsoni, maritimus, henslowii, and X. baileyi). Zink and Avise (1990) studied relationships among most members of the group (Xenospiza lacking) using allozyme and mitochondrial RFLP (restriction fragment length polymorphism) data. Both data sets identified a maritimus-caudacutus-nelson-leconteii clade and a probable henslowii-bairdii sister relationship. The allozyme evidence suggested a genetically distinct group consisting of savannarum and the South American forms humeralis and aurifrons, but their placement within the ingroup was equivocal. They concluded that the genus Ammodramus may not be monophyletic and that additional analyses using more distant outgroups are required. Here, we undertake such analyses.
Taxonomic treatment (abridged) of members of the grassland sparrow complex at various times over the past 120 years.
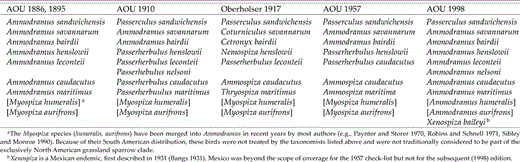
Taxonomic treatment (abridged) of members of the grassland sparrow complex at various times over the past 120 years.
A thorough systematic revision requires that all members of the clade of interest be identified. In addition to “known” ingroup taxa, genera with uncertain taxonomic affinities should be examined, as well as those taxa linked with members of the ingroup by earlier taxonomies. Because closely related taxa provide the optimal outgroups for rooting trees (Wheeler 1990, Smith 1994), these, too, need to be identified. In short, a modern phylogenetic hypothesis of generic relationships among all New World sparrows is required, and such a hypothesis does not yet exist. A few, less complete, systematic studies on higher-level sparrow relationships have been done. Patten and Fugate (1998) used morphological, behavioral, oological, and allozymic characters to investigate systematic relationships among “the emberizid sparrows.” They examined 18 genera, all from Paynter's (Paynter and Storer 1970) “first group” and considered by Paynter to represent the “typical” emberizines. Most of the characters used in that work were the same as those used to originally describe the genera under investigation. Not surprisingly, many traditional taxonomic groupings were recovered, including a grassland sparrow clade comprising Xenospiza baileyi, Passerculus sanwichensis, Ammodramus savannarum, A. henslowii, and A. bairdii. By contrast, Carson and Spicer (2003) used modern molecular methods to investigate sparrow relationships. Their results identified a well-supported clade that contained some members of the grassland sparrow complex, including P. sandwichensis, A. leconteii, and A. henslowii but also all members of the putative brushland sparrow genus, Melospiza. Placed as sister to this assemblage was an Amphizpiza belli-Pooecetes pairing. Their unusual result suggests a taxonomic arrangement that differs considerably from that found in recent classifications (e.g., Sibley and Monroe 1990, AOU 1998).
The goals of the present study are twofold. First, the work of Carson and Spicer (2003) is extended to include additional unsampled genera, with the goal of conclusively identifying all members of the grassland sparrow clade. Second, once ingroup membership is established, we will focus on discerning relationships within the clade. Given the checkered taxonomic history of this group (Table 1), a definitive revision is warranted.
Materials and Methods
Sampling strategy.—
All 11 currently recognized members (Sibley and Monroe 1990) of the traditional grassland sparrow assemblage were included in this work. To ensure that no potential ingroup members were omitted, we also examined at least one representative of each additional, currently recognized (Sibley and Monroe 1990) sparrow genus, excepting the monotypic Cuban form Torreornis. Analyses of these preliminary data (J. Klicka unpubl. data) identified a well-supported clade that included all genera traditionally considered members of the grassland sparrow assemblage but also representatives of the following additional genera: Oriturus, Pooecetes, Amphispiza, and Melospiza. Subsequently, all these taxa were considered a part of the ingroup for the present study. The preliminary analysis also identified a well-supported sister clade from which appropriate outgroups were chosen. The six outgroup taxa selected each represent a subclade within this larger sister clade. Complete ingroup and outgroup species representation is provided in Table 2.
Collecting dates and localities of specimens used in phylogenetic reconstructions.
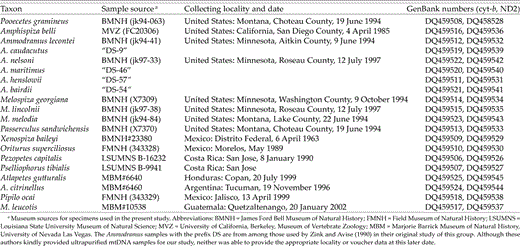
Collecting dates and localities of specimens used in phylogenetic reconstructions.
Laboratory protocols.—
Total genomic DNA was extracted from all specimens using a DNeasy tissue-extraction kit (Qiagen, Valencia, California), following the manufacturer's protocol. We amplified the mitochondrial DNA (mtDNA) cytochrome-b (cyt-b) gene using the primers L14764 (Sorenson et al. 1999) and H4A (Harshman 1996), and the NADH dehydrogenase subunit 2 (ND2) gene using L5215 (Hackett 1996) and H6313 (Johnson and Sorenson 1998). Because the X. baileyi sample was obtained from a study skin, DNA extraction and polymerase chain reaction (PCR) preparation were performed in a separate laboratory where no previous avian genetic work had been conducted. Amplifications of Xenospiza divided each gene into two fragments using internal primers: L 5758 and H5766 (Sorenson et al. 1999) for ND2 and LCBOB and H15299 (Klicka et al. 2001) for cyt-b. For all fragments, amplifications were done in 12.5-μL reactions under the following conditions: denaturation at 94°C, followed by 40 cycles of 94°C for 30 s, 54°C for 45 s, and 72°C for 1 min. This was followed by a 10-min extension at 72°C and a 4°C soak. Products were purified using a Qiaquick PCR purification kit (Qiagen) or ExoSAP-IT (USB Corporation, Cambridge, Massachusetts) purification following the manufacturer's protocols. Standard, 20-μL sequencing reactions were performed using 4 μL of BigDye (Applied Biosystems, Foster City, California) and 20–40 ng of purified and concentrated PCR product. Products of these reactions were purified using a magnetic-bead clean-up procedure designed by Agencourt Bioscience (Beverly, Massachusetts) and run on an ABI 3100-Avant automated sequencer. Complementary strands of each gene were unambiguously aligned using SEQUENCHER, version 4.2 (Gene Codes Corporation, Ann Arbor, Michigan). The veracity of the sequence data was supported in several ways. Both light and heavy strands were sequenced for all PCR fragments. No gaps, insertions, or deletions were apparent in the aligned sequences, and all data translated (using MEGA, version 3.01; Kumar et al. 2004) correctly into amino acid form. The resulting sequences include the complete cyt-b (1,143 base pairs [bp]) and ND2 (1,038 bp) genes for a total of 2,181 bp of concatenated data.
Phylogenetic protocols.—
Phylogenetic analyses were preceded by data exploration. Using PAUP*, version 4.0b4a (Swofford 2000), we constructed genetic-distance matrices using both inter- and intrageneric pairwise comparisons. The relatively low genetic distances uncovered suggested limited potential problems attributable to homoplasy. Nevertheless, we plotted pairwise comparisons of uncorrected genetic distances for each gene and gene partition (i.e., codon position) for all ingroup taxa to assess the degree of possible “saturation” effects. The evolutionary dynamics of each gene and gene partition were also investigated for all ingroup taxa. Parameters examined include transition: transversion ratio (Ts:Tv), relative rates of evolution, nucleotide composition (%), and the gamma-shape parameter (α). Potential nucleotide composition bias was assessed by performing a series of chi-square tests of homogeneity on the informative data of each gene and gene partition. For each gene, we also plotted the relative proportions of each nucleotide for each taxon used (e.g., C vs. T and A vs. G). Outliers in such plots likely indicate taxa that are problematic with respect to nucleotide composition biases. To ensure that the data sets for each gene contained congruent phylogenetic signal, we executed a partition homogeneity test (the incongruence length difference test [ILD] of Farris et al. 1995) in PAUP*. This test consisted of 100 replicates and considered only informative characters (Cunningham 1997). Given that no significant differences were identified (P = 1.00), we combined the data for all analyses.
We performed phylogenetic analyses using both maximum-parsimony (MP) and maximum-likelihood (ML) approaches. We conducted both weighted and equal-weighted parsimony analyses. In the former, transitions were downweighted in relation to transversions by one-third for both genes. Support for individual nodes was assessed using MP heuristic bootstrap (Felsenstein 1985) with 500 pseudoreplicates, each with 10 random-addition sequence replicates. We used MODELTEST, version 3.04 (Posada and Crandall 1998), to select the most appropriate model of sequence evolution for ML analyses. Akaike's Information Criterion (AIC) was used (see Posada and Buckley 2004) to identify the GTR + I + Γ model as the best fit to our combined data. We obtained an ML estimate of phylogeny using PAUP* (Swofford 2000) with parameter settings as determined by MODELTEST. Node support for ML analyses was determined via bootstrapping (Felsenstein 1985), with 100 pseudoreplicates and full heuristic searches using random addition of taxa. For both MP and ML bootstrap analyses, we considered values of >70% to indicate good node support. It is well understood that one of the shortcomings of MP is its inability to detect homoplasy on long branches, a potential source of bias in phylogeny estimation (Felsenstein 1978, Swofford et al. 1996). Because the model chosen is more resistant to error caused by homoplasy (Kuhner and Felsenstein 1994, Huelsenbeck 1995), we decided a priori to consider our likelihood topology as our best estimate of a phylogenetic hypothesis for the grassland sparrow assemblage.
Bayesian inference (Rannala and Yang 1996) was used primarily as a means of assessing support for nodes obtained via other (ML, MP) tree-building methods. Because the use of a single evolutionary model with data composed of differently evolving subsets may result in mismodeling and significant systematic error (Brandley et al. 2005), we partitioned our data by gene (cyt b and ND2) and ran each independently through MODELTEST to determine the AIC best-fit model. The program MRBAYES, version 3.1 (Huelsenbeck and Ronquist 2001) was then implemented using the TVM + I + Γ and TrN + I + Γ models of sequence evolution for the respective cyt-b and ND2 partitions. Specific nucleotide-substitution model parameters were left undefined and estimated as part of the analysis. All Bayesian analyses were initiated from random starting trees. Four Markovchain Monte Carlo chains were run for 1 million generations and sampled every 100 generations, yielding 10,000 trees. The first 100,000 generations (= 1,000 trees) were discarded to ensure that chain stationarity had been reached. To ensure that the Markov chain was sampling from the posterior distribution, this procedure was repeated two more times. Because all three runs converged on the same distribution, all trees (excluding those sampled before burn-in) were combined, yielding a total of 27,000 topologies from which a 50% majority-rule consensus tree was reconstructed. Nodes having posterior probability values of 95% or greater on this tree were deemed significantly supported.
Results
Sequence characteristics.—
As expected, the ND2 gene was slightly more variable than cyt-b (Table 3). Over the 2,181 bp of combined sequence, 610 (28%) sites were variable and, of these, 363 (16.6%) were potentially phylogenetically informative. Overall, slightly more than 47% of third-position sites varied. All plots of genetic distances (not shown) were linear, indicating that homoplasy is relatively low. In birds, cyt b approaches saturation between 8% and 9% divergence (Griffiths 1997). For our data, uncorrected cyt-b ingroup distances ranged from 1% (A. nelsoni-A. caudacutus) to 7.9% (A. maritimus-M. lincolnii) divergent, with a mean difference of 6.3% (Table 4). Corresponding values from ND2 distances are greater in all comparisons, ranging from 2% (A. nelsoni-A. caudacutus) to 13.2% (A. belli-M. georgiana), with an average of 10.8%. This latter value is approaching the known saturation point of the ND2 gene (10–12% [Hackett 1996], 12–13% [Johnson and Sorenson 1998]).
Overall and codon-position-specific dynamics of the cytochrome-b and ND2 genes for all ingroup taxa. Mean base composition is averaged over all sequences using PAUP*. Transition:transversion ratio (Ts:Tv) values are the average number of changes reconstructed on one of two topologies obtained with all sites having equal weight. The Ts:Tv and α values were estimated simultaneously for each partition.
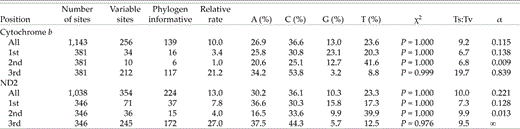
Overall and codon-position-specific dynamics of the cytochrome-b and ND2 genes for all ingroup taxa. Mean base composition is averaged over all sequences using PAUP*. Transition:transversion ratio (Ts:Tv) values are the average number of changes reconstructed on one of two topologies obtained with all sites having equal weight. The Ts:Tv and α values were estimated simultaneously for each partition.
Uncorrected divergences (%) for the cytochrome-b (above) and ND2 (below) genes. The outgroups have been averaged, yielding a single value for pairwise ingroup comparisons (column 1, below).
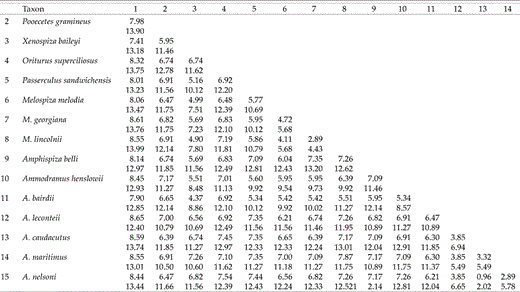
Uncorrected divergences (%) for the cytochrome-b (above) and ND2 (below) genes. The outgroups have been averaged, yielding a single value for pairwise ingroup comparisons (column 1, below).
Nucleotide composition and bias varies slightly between these two genes; both display a deficiency of guanine and an excess of cytosine nucleotides. Base composition biases (Table 3) recovered are similar to those reported from other avian studies. Tests of homogeneity of base frequencies across ingroup taxa were not significant for both genes combined (χ2 = 8.57, df = 57, P = 1.00), each individual gene, or any gene (codon) partition (Table 3). Codon-position-specific, gamma-shape parameter (α) estimates indicate that among-site rate heterogeneity is a likely problem in this data set. The problem is most acute at second-position sites where values for both genes (cyt-b = 0.009, ND2 = 0.013) lie outside the range (0.1–0.5; Yang 1996) typical of gamma-shape parameter estimates.
Phylogenetic analyses.—
Interpreting the results of alternative phylogenetic methods is straight-forward, because the weighted MP, ML, and Bayesian topologies were identical (Fig. 1). All methods support a Melospiza-Xenospiza sister relationship. Ammodramus is clearly shown to be polyphyletic, with its members distributed among three clades. Ammodramus bairdii and A. henslowii are embedded within a well-supported clade that also contains Passerculus, Xenospiza, and Melospiza. A separate, well-defined clade comprises the forms leconteii, caudacutus, nelsoni, and maritimus. The remaining members of Ammodramus (savannarum, humeralis, aurifrons) are only distantly related to the present clade and are instead most closely linked with members of the genus Arremonops (J. Klicka unpubl. data). Amphispiza belli is placed among the grassland sparrows as sister (though support is lacking) to the monotypic form Pooecetes. The genus Amphispiza is thus rendered polyphyletic, given that its congeners (bilineata and quinquestriata; Sibley and Monroe 1990) have strong affinities elsewhere in the emberizid phylogeny, closest to the monotypic forms Chondestes and Calamospiza (J. Klicka unpubl. data). This relationship was also suggested by Carson and Spicer (2003). The obscure Mexican endemic Oriturus is placed within the grassland sparrows, but its taxonomic affinities within the clade are uncertain. It likely represents a relict lineage with no close, extant “relatives.”
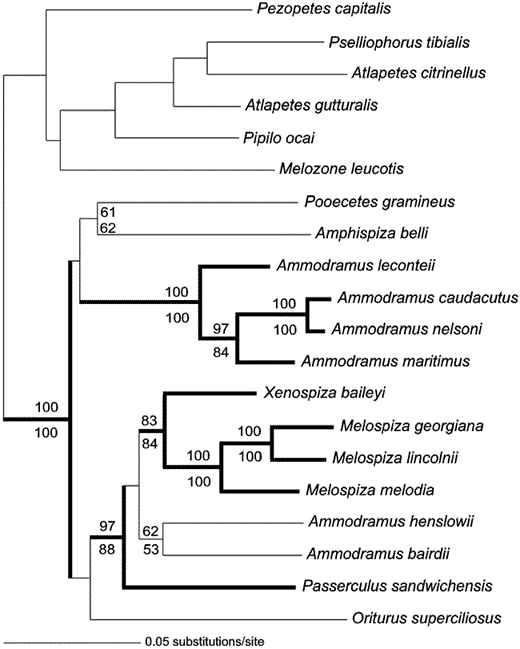
Phylogenetic hypothesis for the “grassland” sparrows as determined by weighted parsimony, maximum likelihood, and Bayesian methods. Branch lengths reflect likelihood estimates (GTR + I + Γ model of evolution, I = 0.5831, α = 2.2334; -Ln length = 10,910.2456). Nonparametric bootstrap support as determined via likelihood and parsimony methods are indicated above and below (respectively) the nodes. Bold lines indicate significant (>95%) Bayesian posterior support values.
Bayesian posterior probabilities have been criticized as too liberal (Suzuki et al. 2002) and prone to a high type-1 error rate (Erixon et al. 2003; but see Wilcox et al. 2002, Alfaro et al. 2003), whereas nonparametric bootstrapping proportions are known as conservative estimates of phylogenetic accuracy (Hillis and Bull 1993). In the present study, nodes identified as well supported via each of these methods are in complete agreement. Eight of 13 ingroup nodes (Fig. 1) have posterior probabilities of ≥95% and ML and MP bootstrap proportions of ≥70%. No nodes are supported by one confidence estimate but not the others. This consensus gives us a high degree of confidence in those supported portions of the tree and, overall, we submit that Figure 1 is our best estimate of phylogenetic relationships for this sparrow clade. However, caution should be used in interpreting those five nodes in the tree that are less well supported. Therefore, we consider Figure 2 our most reliable estimate of these relationships (after Lanyon 1993).
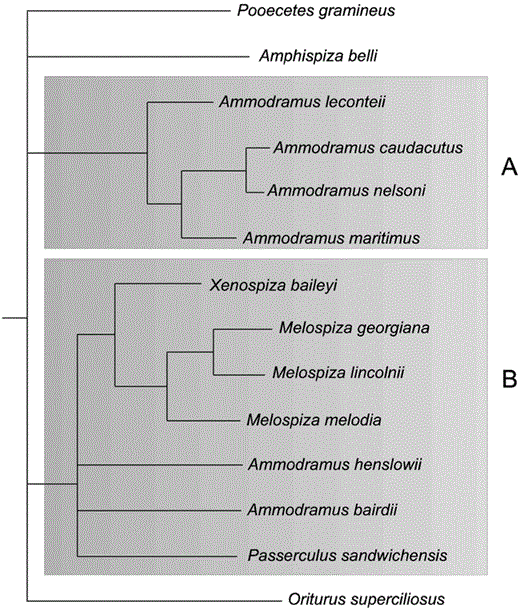
Consensus tree based on MP, ML, and Bayesian analyses with all weakly supported nodes collapsed. This tree represents our most “reliable estimate” (Lanyon 1993) of phylogenetic relationships among members of this clade. Subclades identified as (A) and (B) are discussed in the text.
Discussion
Systematics overview.—
We describe a well-defined clade that includes most of the elements of the historical grassland sparrow assemblage but also all members of the genus Melospiza, one member of the genus Amphispiza (belli), and the monotypic forms Pooecetes and Oriturus. The clade recovered was not expected; this particular taxon assemblage had not been predicted in any previous taxonomy. Relationships within the historically problematic genus Ammodramus are resolved here, as is the debate concerning the systematic position of Xenospiza. We identify both Amphispiza and Ammodramus as polyphyletic genera, corroborating the results of Carson and Spicer (2003). One of the strengths of this analysis is that it includes all likely members of this clade, enhancing the likelihood that the supported relationships shown reflect the true species relationships. We discuss below, in greater detail, some of our salient findings from a historical and taxonomic perspective.
Relationships within our Ammodramus “phylogeny” are novel with respect to earlier taxonomies (Table 1; also see review in Murray 1968), though some of the relationships we uncovered are reflected in those efforts. For example, the species leconteii, caudacutus (nelsoni) (considered a subspecies of caudacutus by most taxonomists at the time), maritimus, and henslowii were treated as a group, first as Ammodramus (Ridgway 1901), then as Ammospiza (Oberholser 1905), and later as Passerherbulus (Stone 1907, AOU 1910). Our analyses indicate that caudacutus, nelsoni, maritimus, and leconteii form a well-resolved clade (Fig. 2A). Ammodramus henslowii was paired with bairdii in all our analyses (but always with low support). Although henslowii and bairdii are typically placed near each other, linear taxonomies had not previously suggested a sister relationship, and none has suggested a closer relationship with Passerculus (and Melospiza) than with the other members of Ammodramus. That savannarum, humeralis, and aurifrons are not members of the reconfigured grassland sparrow clade is among our more striking findings, though similar results were obtained by Carson and Spicer (2003). The taxonomy of the genus has long been centered around savannarum, the type species for this genus (Swainson 1827). Our results with respect to the genus Ammodramus are similar to those obtained by Zink and Avise (1990), who analyzed mtDNA restriction fragments and allozymes. They also identified a well-resolved caudacutus-(nelsoni)-maritimus-leconteii clade and suggested that henslowii and bairdii represent a “relatively old sister species pair.” Their work, however, was hindered by incomplete taxon sampling. They recognized that savannarum, humeralis, and aurifrons were “genetically distinct from the others” and that the genus Ammodramus “is possibly not monophyletic,” but they lacked the data necessary to document correct generic limits.
Xenospiza baileyi, a Mexican highland endemic, occurs today only in sacaton bunch grass (Sporobolus sp.) habitat in La Cima Pass between Mexico City and Cuernavaca. Specimen records from a disjunct population in the Sierra de Bolaños region of Jalisco exist, but the species was not found there in recent surveys (J. Klicka unpubl. data). The correct placement of this sparrow has long been a topic of a taxonomic controversy, which continues to this day. Xenospiza evidently possesses a suite of morphological characters that suggest a variety of different generic-level assignments. In describing this species, Bangs (1931) emphasized color and plumage patterns and placed it among the grassland sparrows, believing that it was most similar to Ammodramus leconteii. Pitelka (1947) dismissed these characters as “superficial” and concluded that in characters of size, proportions, and wing and tail shape, Xenospiza was most similar to Melospiza lincolnii of the brushland sparrow group. Believing that data on nests, eggs, juvenal plumages, song, and behavior provide “a more reliable basis for evaluating the systematic relationship of this elusive sparrow,” Dickerman et al. (1967) considered it closest to other “grassland-nesting” sparrows, though these authors suggested that without more compelling evidence it should be retained as a monotypic genus. Each of these authors was partly correct. According to our results, Xenospiza is embedded within the grassland sparrow clade, within which it is placed as sister to the three members of the traditional brushland sparrow genus, Melospiza. The remaining members of the putative brushland sparrow complex, Zonotrichia, Junco, and Passerella, are elements of a well-resolved clade (J. Klicka unpubl. data) that lies outside of the grassland clade and its sister, from which outgroups were chosen.
The “brush-inhabiting” (Paynter 1964) or “brushland-nesting” sparrows (Dickerman et al. 1967) (Passerella, Melospiza, Zonotrichia, and Junco) have long been considered closely allied species. This was attributable, at least in part, to a number of well-documented intergeneric hybridizations (e.g., Dickerman 1961, Short and Simon 1965). Largely for this reason, Passerella, Melospiza, and Zonotrichia have previously been lumped into the genus Zonotrichia (Paynter 1964); Short and Simon (1965) went a step further, lumping all four of these genera into Junco. Within this assemblage, Melospiza and Passerella are most similar morphologically, which leads some authors to merge the two (e.g., Linsdale 1928, Mayr and Short 1970). Our results suggest that many of the morphological characters and ecological affiliations traditionally used to define sparrow assemblages are of questionable taxonomic utility (see below). These results also (once again) emphasize that hybridization events can be a poor measure of phylogenetic relatedness (Prager and Wilson 1975, Klicka et al. 2001).
The newly defined grassland sparrow clade includes three additional taxa, Pooecetes gramineus, Oriturus supercilisosus, and Amphispiza belli, of which the first two represent monotypic genera. These three are morphologically disparate taxa, bearing little resemblance to one another or to other members of this clade. Oriturus is a large, bulky sparrow that is endemic to the highlands of northwestern and central Mexico, where it occupies bunch grasses and open pine woods. In form, Oriturus resembles some members of the genus Aimophila. Pooecetes breeds widely across middle and northern latitudes of North America, in open habitats including grasslands, shrubsteppe, and fallow croplands. It is a medium-sized, rather typical “streaky-brown” sparrow that also possesses a unique combination of morphological characters including a white eye ring, white outer rectrices, and a chestnut-colored scapular patch. We suggest that both these species remain monotypic only because diagnostic morphological clues to relationships are lacking. The genus Amphispiza comprises three species, according to some authors (e.g., Sibley and Monroe 1990): belli, bilineata, and quinquestriata. Others have merged one (quinquestriata; Paynter and Storer 1970, AOU 1998) or all (Phillips et al. 1964, Mayr and Short 1970) of these species into Aimophila. Our results confirm polyphyly for this genus (Carson and Spicer 2003), because belli is without question a member of the clade of focus. Amphispiza quinquestriata does not belong among the Aimophila (contraPaynter and Storer 1970, AOU 1998, Patten and Fugate 1998), but is instead sister to bilineata and closest to Chondestes and Calamospiza (J. Klicka unpubl. data; see Carson and Spicer 2003).
Utility of morphological characters.—
The composition of our “grassland” sparrow clade, and the relationships among its component taxa, differ from all previous morphology-based taxonomic hypotheses. That is, morphological and genetic estimates of relationships for this group are incongruent. Generic-level relationships among sparrows have been the focus of a single modern, phylogenetic study (Patten and Fugate 1998) using the structural and plumage characters used traditionally by avian taxonomists. Not surprisingly, the results of that study supported the monophyly of all currently accepted genera. In that work, species of Ammodramus sampled included henslowii, bairdii, and savannarum. Our study indicates that the taxonomic affinities of the latter lie well outside this clade. Patten and Fugate's (1998) analysis also recovered other traditional groupings, such as the linking of Melospiza with Passerella, and Amphispiza belli with A. bilineata; both are incorrect, according to the molecular evidence.
It is difficult to fault earlier efforts that were unable to recover our phylogeny. The component species of our grassland sparrow clade are a morphologically heterogeneous assemblage, and we know of no nonmolecular taxonomic characters that would suggest that they form a group. Furthermore, from the perspective of traditional taxonomy, it is difficult to examine specimens and not come away with the impression that A. savannarum belongs somewhere near the other Ammodramus taxa and that Melospiza and Passerella are most similar to one another. Traditional comparative methods support these relationships. At some taxonomic levels and for some groups, morphological cues indeed provide adequate representation of taxon relationships. For example, both morphological and molecular characters identify a clade of Melospiza spp. and a group of “sharp-tailed” Ammodramus species (Fig. 2A). By contrast, however, the traditional placement of the savannarum-humeralis-aurifrons assemblage within the grassland sparrow group likely represents a case of morphological convergence, a potential pitfall for morphological analyses. Rather abrupt shifts in morphology can also lead to incorrectly defined relationships, particularly among genera. For example, the genus Melospiza had not previously been recognized as a member of this clade, despite the fact that it appears to be embedded within it. Morphologically, Melospiza departs from an otherwise uniform “Bauplan” that links the members of Ammodramus and Passerculus. Also problematic for morphological taxonomy are those taxa with no close relatives. In our clade, Oriturus, Pooecetes, and Amphispiza belli represent relatively basal lineages with no subsequent bifurcations. These “older” species are difficult to place within the context of genera whose members share suites of morphological characters. We do not doubt that some traditional taxonomic characters may be phylogenetically informative. The challenge before us is to identify those characters that are reliable indicators of relationships and those homoplastic characters that are not.
Utility of ecological characters.—
Robins and Schnell (1971) performed a thorough phenetic analysis of the grassland sparrow complex, measuring 48 skeletal features of multiple exemplars for each putative member of the group. They concluded that the assemblage should be divided into two genera. Their clade of Ammodramus comprised savannarum, humeralis (aurifrons), bairdii, and P. sandwichensis, whereas their clade of Ammospiza contained leconteii, caudacuta (nelsoni), maritimus, henslowii, and X. baileyi. These authors noted that all species of the latter “breed in or near marshes” and gave them the name “marshland sparrows.” Similarly, because of habitat association, members of their Ammodramus clade were designated the “grassland” sparrows. It is well known that trees derived from morphological comparisons can reflect a history of adaptive responses, which may or may not coincide with phylogenetic history (Endler 1982). Results of the present study and those of Zink and Avise (1990) indicate that the morphometric analyses of Robins and Schnell (1971) identified ecological associations rather than true evolutionary relationships. For clarity, we have used the terms “grassland” and “brushland” to describe putative ecological and evolutionary sparrow assemblages. The original configurations of both these groups are polyphyletic, which suggests that generalized and simplistic ecological associations may have limited value for phylogenetic inference. The single sparrow clade of study contains “grassland” and “brushland” forms but also species that are restricted to montane, lowland, mesic, or xeric environments. With respect to habitat preference, this is a heterogeneous group, and among-lineage habitat shifts occur regularly within the sparrows studied.
Taxonomic implications.—
Because our study benefits from thorough taxonomic sampling and identifies several well-resolved nodes, we believe that the following suggested nomenclatural changes are justified. The polyphyly of Ammodramus necessitates either the naming of new genera or the resurrection of former generic designations. The type species for this genus is savannarum (A. bimaculatus; Swainson 1827). This generic epithet must therefore remain with the savannarum-humeralis-aurifrons clade that lies outside of our clade of interest. We suggest that the genus Ammospiza (Oberholser 1905) be resurrected for the leconteii-caudacutus-nelsoni-maritimus clade (Fig. 2A). This name has taxonomic priority and, with the addition of leconteii, reverts to the AOU (1957) checklist. The Xenopiza-Melospiza-Ammodramus (in part)-Passerculus clade (Fig. 2B) is a bit more problematic. Likely, some taxonomists would favor merging Xenospiza and Melospiza, resurrecting the genus Passerherbulus (Stone 1907) for henslowii and bairdii, and retaining Passerculus as a monotypic form. Such a taxonomy would be consistent with the relationships depicted in Figure 1. However, Figure 2B indicates that precise relationships within the group remain equivocal. A henslowii-bairdii sister relationship is not certain, nor is the relationship of this putative pair with either Melospiza or Passerculus. To accommodate this uncertainty and reflect the known evolutionary pattern, we advocate merging all these taxa (Fig. 2B) into a single genus. Among the genera listed, Passerculus (Bonaparte 1838) has priority and should be used. The long branches of Pooecetes, Amphispiza, and Oriturus suggest a more distant relationship with other clade members and an uncertain placement within the group. This uncertainty should be reflected by the retention of their monotypic names. Thus, Pooecetes and Oriturus would remain as they are; A. belli, however, presents a more difficult taxonomic problem. The type species for the genus Amphispiza is bilineata (Coues 1874), which, along with A. quinquestriata, occurs well outside our study clade. With that name taken, belli is alone at the generic level and in need of a generic epithet. Throughout much of its breeding range, this sparrow is associated with sagebrush (Artemesia tridentata) habitats, and a descriptive name incorporating this association may be appropriate. The name for this genus of sage is derived from the Greek word Artemis. We therefore offer Artemisospiza as a new monotypic genus for the form belli.
Acknowledgments
We thank the curators, staffs, and collectors at the institutions that provided the tissue samples critical for the completion of this study. R. Banks and D. Nicolson kindly provided advice on nomenclatural changes. We thank C. Cicero and two additional reviewers for their constructive comments on an earlier draft of this manuscript. This work was funded, in part, by NSF DEB 0315469 (to J.K.) and the Barrick Museum Foundation.
Literature Cited

{kind=link}
{kind=link}