





Abstract
The Aquatic Warbler (Acrocephalus paludicola) has a unique mating system. Males are free of any parental duties and can fertilize many females during a single breeding season. Females also usually copulate with more than one male, and nestlings in a single nest may be sired by as many as five males. Paternal success of 33 Aquatic Warbler males in a Polish population was determined by microsatellite polymerase-chain-reaction analysis. Males infected by blood parasites (trypanosomes) sired fewer offspring, weighed less, and probably arrived later to breeding grounds than uninfected males. Number of nestlings sired by a male correlated positively with its fat deposits and wing length. These findings indicate that male body condition directly influences paternal success. Number of sired nestlings per individual male ranged from one to eight. Reproductive success among males was uneven, with six males (18%) fathering 44% of nestlings in the study area. Males with high reproductive success arrived at the breeding grounds earlier in spring than males with low reproductive success.
Resumen
El Éxito Reproductivo de los Machos Está Correlacionado con los Niveles de Parásitos en la Sangre y la Condición Corporal en la Especie Promiscua Acrocephalus paludicota
Acrocephalus paludicola tiene un sistema de apareamiento único. Los machos se ven liberados de toda responsabilidad parentaly pueden fecundar a muchas hembras durante una época reproductiva. Las hembras también copulan generalmente con más de un macho, y los polluelos en un nido pueden haber sido engendrados por un máximo de 5 machos. Se determinó el éxito parental de 33 machos de A. paludicola en una población polaca, utilizando un análisis de microsatélites por medio de la reacción en cadena de la polimerasa. Los machos infectados por parásitos de sangre (tripanosomas) engendraron menos progenie, pesaron menos y probablemente Ilegaron más tarde a los territorios de cría que los machos no infectados. El número de polluelos engendrados por un macho se correlacionó positivamente con sus depósitos de grasa y longitud del ala. Estos hallazgos indican que la condición corporal del macho influencia directamente el éxito parental. El número de polluelos engendrados por cada macho varió entre 1 y 8. El éxito reproductivo entre los machos no fue parejo; seis machos (18%) fueron los padres del 44% de los polluelos en el área de estudio. Los machos con alto éxito reproductivo Ilegaron antes a los territorios de cría en la primavera que los machos con un éxito reproductivo menor.
Blood parasites have been shown to negatively affect reproductive success of birds (especially females) in several studies (discussed below). In the present study, we analyzed variation in reproductive success of individual males in a Polish population of Aquatic Warblers (Acrocephalus paludicola). Specifically, we asked whether male reproductive success is affected by blood parasites and body parameters. Hamilton and Zuk's (1982) hypothesis predicts that resistance to endoparasites is inherited. Data supporting such inheritance are scarce. Møller (1990), in his summary article, listed only three papers that proved intraspecific heritable resistance to parasites. In any case, resistance to parasites can be an important factor in female choice, and thus influence male reproductive success. Some male body parameters may have a similar influence.
The Aquatic Warbler inhabits Carex marshlands, which are particularly rich in arthropods, in fertile river valleys, mainly in Belarus, Poland, and Ukraine. Nests of the Aquatic Warbler are often clustered in places where food resources are abundant (Dyrcz and Zdunek 1993). Within clusters, females are aggressive toward one another, and their foraging grounds are exclusive during the nestling-feeding period. Males have defined singing posts; however, no distinct agonistic behavior among males was observed in the field. Telemetric data indicate that males regularly visit large areas throughout the population; as a consequence, their home ranges often overlap (Schmidt et al. 1999, Schaefer et al. 2000). The Aquatic Warbler has a unique mating system, which is classified as a kind of promiscuity (Heise 1970; Birkhead 1993; Schulze-Hagen et al. 1993, 1995; Dyrcz et al. 2002). Males can sire nestlings in the nests of many females, and eggs in one nest may be fertilized by more than one male. There are no leks or arenas, but males sing during the whole breeding season from more-or-less fixed singing posts. Males do not participate in parental duties. The high density of potential prey may explain and facilitate the uniparental rearing of offspring by the female. The percentage of broods with nestlings sired by more than one male is high, ranging from 54% to 92% (n = 62) across years; a maximum of five fathers per brood was detected in four broods (Dyrcz et al. 2002).
Materials and Methods
Field study and laboratory methods
Field studies were done in 1997 on a 44-ha plot of fen mires in the southern basin of the Biebrza River in northeastern Poland (53°20′N, 22°40′E). A large part of the study area was a bed of Carex tussocks, in which C. appropinquata was the dominant species. Characteristic features of the area were an extensive moss layer on the ground and scattered bushes of Salix spp., 1 m in height on average (for further details, see Dyrcz and Zdunek 1993). Water depth among tussocks varied between 1 and 5 cm during the study period. A grid of 100 × 100 m squares, marked by colored poles, was established to allow orientation in the field and mapping of nests. Females were mistnetted near their nests, and males near their singing posts. They were weighed on a Pesola spring balance (range 50 g, sensivity 0.5 g). Wing length was measured to the nearest millimeter by the flattened-wing method (Svensson 1992), and length of the bill was measured to the nearest tenth of a millimeter. Fat deposit was determined visually by one person, according to the six-degree scale (0–5) used by Operation Baltic in Poland (Busse 1970).
Blood samples
Blood samples were collected from 114 nestlings on their 8th and 11th days of life, from 27 breeding females, and from 33 males. A total of 27 broods were available for DNA analysis. Only the first brood of each female each year was examined. We tried to mistnet and individually color-band all the males present during the breeding season in our study area (44-ha sample plot) to take blood samples.
To detect blood parasites, blood smears were made from peripheral blood (one drop) taken from the brachial vein, air-dried, fixed in methanol, and stained with Haemacolor (Merck, Darmstadt, Germany). Whole smears were checked for large parasites, such as Trypanosoma spp. and Microfilaria spp., under 400× magnification without immersion oil. Then, 100 microscopic views were used to check for parasites from the Haemogregarina family under 1,000× magnification with immersion oil.
Molecular methods
DNA isolation: 10– 100 μL of blood was collected using a syringe and mixed 1:10 with a modified EDTA-Tris buffer (0.1 M Tris, pH 7.4, 10% EDTA, 1% NaF, 0.1% thymol) or ethanol. Samples were stored at 4°C for up to six months, with negligible effects on yield and quality of DNA.
Blood samples were incubated overnight at 37°C in 1 mL lysis buffer (10 mmol Tris [pH 7.5], 25 mmol EDTA, 75 mmol NaCl, 1% SDS) with 1 mg of proteinase K (Merck), followed by a standard phenol-chloroform protein extraction. DNA was precipitated with 0.8 vol of cold isopropanol, centrifuged, washed, dried, and resuspended in TE buffers (Sambrook et al. 1989, Swatschek et al. 1994).
Polymerase chain reaction and detection of its products
Microsatellite polymerase chain reaction (PCR): For amplifications, ∼30 ng of total DNA from Aquatic Warblers was used as a template, plus 20 pmol microsatellite primers HrU6A 5′-GCT GTG TCA TTT CTA CAT GAG-3′ and HrU6B 5′ ACA GGG CAG TGT TAC TCT GC-3′; AAr3F 5′-GCA TCT GGT CTC CGA TTG TT-3′; AAr3R 5′-TTT GGG TTA CAT CTG AGT GTG C-3′, and FhU2A 5′ GTG TTC TTA AAA CAT GCC TGG AGG-3′ and FhU2B 5′GCA CAG GTA AAT ATT TGC TGG GCC-3′ (Ellegren 1992, Primmer et al. 1996), 1.5 mmol MgCl2, 2.5 mmol of dGTP, dCTP, and dTTP, mmol dATP, 2 μCi alpha-33P-dATP, 2.5 μL 10× amplification buffer (100 mmol Tris HCl, pH 8.5, 500 mmol KCl, 5% Triton ×-100), and 0.8 units of Taq-Polymerase (Pharmacia, Freiburg, Germany), in a total volume of 25 μL. After an initial denaturation (180 s at 94°C), 39 cycles of 30 s at 94°C, 60 s at 55°C, and 60 s at 72°C were performed on a thermocycler (Biometra, Göttingen, Germany), followed by 72°C for 4 min and storage at 4°C. Products of PCR were electrophoretically separated on a Sequagel matrix (Biozyme, Oldenburg, Germany) (size 44 × 37 cm) at 65 W for 100 min. After drying, the gel was exposed to an x-ray film (BioMax MR2 Kodak) for two to six days and developed. Size of the microsatellites was determined with a sequence of a plasmid pGEMT (Stratagene, La Jolla, California), run on the same polyacrylamide gel (for details, see Dyrcz et al. 2002).
Analysis of microsatellite data
Approximately 22 separate alleles were found for the HrU6 locus, 17 alleles in the AAr3 locus, and 13 alleles in the Fhu2 locus. The percentage of birds with identical alleles (i.e. homozygous) was low; heterozygosity H accounts for 0.93 at the HrU6 locus. The combination of the three microsatellite primers allowed an unambiguous paternity exclusion in 99% of the young (see also Dyrcz et al. 2002).
Statistics
For statistical analysis, we used Spearman rank correlation and the Mann-Whitney U-test, two-tailed and one-tailed. We believe that the use of the one-tailed test in the case of blood parasites was justifiable, because we assumed that the presence of trypanosomes can only negatively affect male reproductive success.
Results
In the sample of 33 males, individual males sired 1–8 nestlings (x̄ = 2.7, variance = 4.59). Although most males sired only a single offspring, a few males fathered 5–8 young (Fig. 1).
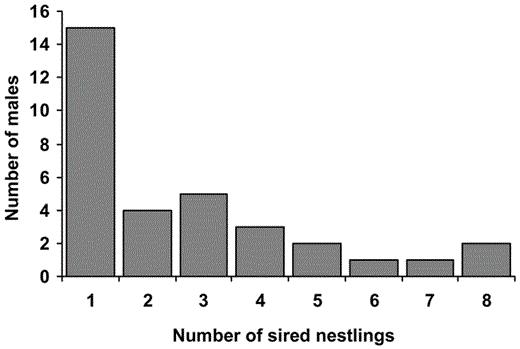
Reproductive success of individual Aquatic Warbler males. Six males sired 5–8 young, whereas 15 males sired a single young
Sixteen of 33 males (48.5%) were infected by Trypanosoma. Males infected with trypanosomes appeared to sire fewer offspring, weighed less, and arrived later at the breeding grounds (Table 1).
Correlation between fitness characters and trypanosome infection (statistics: Mann-Whitney U-test, one-tailed)
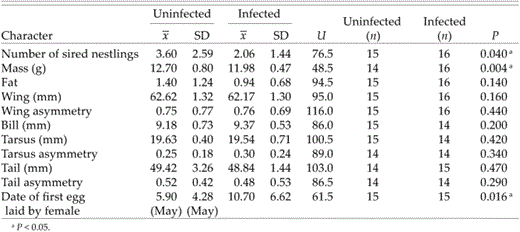
Correlation between fitness characters and trypanosome infection (statistics: Mann-Whitney U-test, one-tailed)
Number of nestlings sired by a single male correlated significantly with its fat deposits, wing length, and the date of the first egg laid in the nests in which it had sired one or more nestlings. Larger (in wing length) and fatter individuals tended to father more nestlings than smaller and leaner ones (Table 2). A negative correlation with the date of egg laying suggests that males with high reproductive success arrived earlier in spring on breeding grounds than males with low reproductive success. Number of nestlings sired by a male did not significantly correlate with his body mass, wing asymmetry, bill length, tarsus length, tarsus asymmetry, tail length, or tail asymmetry (Table 1).
Correlation between number of sired nestlings and variables describing morphological traits of males
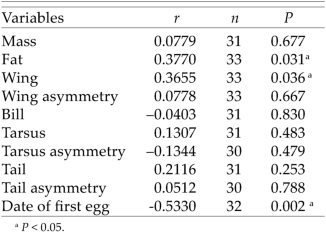
Correlation between number of sired nestlings and variables describing morphological traits of males
Discussion
Blood parasites are widespread among wildfowl (e.g. Shutler et al. 1999, de Jong and Muzzall 2000, de Jong et al. 2001), birds of prey and owls (e.g. Munoz et al. 1999), and especially passerines (e.g. Deviche et al. 2001). Although Trypanosoma spp. are not among the most frequently occurring blood parasites, they have been found in many species of passerine birds (Pruett-Jones et al. 1991; Dale et al. 1996; Rintamaki et al. 1997, 1999; Hatchwell et al. 2000; Blanco et al. 2001; Sehgal et al. 2001; Valkiunas and Iezvova 2001; Dusek and Forrester 2002; Valkiunas et al. 2003; Votypka et al. 2003). The essential question is whether blood parasites actually influence (probably negatively) the condition of their hosts. Populations of some intracellular microparasites probably benefit from the fact that their pathogenicity is minimal, in that they damage expendable cells. For example, although malarial parasites, such as Plasmodium relictum, destroy red blood cells, the effect of that destruction is limited, because those cells have a short lifespan and are regularly removed from circulation by the spleen (Crompton 1997). However, T. avium is a microparasite that lives freely in the plasma in blood vessels (Crompton 1997) and is likely to be more destructive. Several papers have shown that avian blood parasites are harmful to their hosts. In the Great Tit (Parus major), Haemoproteus blood parasites affected the health of their avian host; infected individuals differed from uninfected ones in respect to hematoserological general health and immune parameters (Ots and Horak 1998). It was also found that female Great Tits infected with Trypanosoma parasites laid smaller eggs, hatched proportionally fewer eggs, and raised nestlings with shorter tarsi and generally inferior body condition, as compared with females without trypanosomes (Dufva 1996). Merino et al. (2000), in their field experiments, have reduced through medication the intensity of infection by Haemoproteus spp. and Leucocytozoon spp. in Blue Tits (P. caeruleus) and demonstrated the detrimental effects of natural levels of infection by those parasites on the host's reproductive success and condition. Sanz et al. (2001) found that Pied Flycatcher (Ficedula hypoleuca) females infected with Trypanosoma spp. had a higher probability of deserting their clutches during the incubation period than noninfected females. In Purple Martins (Progne subis), subadult females provisioned nestlings significantly less when infected with Haemoproteus spp. (Wagner et al. 1997). Red-bellied Woodpecker (Melanerpes carolinus) males infected with H. velans had low mass and poor body condition in comparison with uninfected ones; no such association was found in females (Schrader et al. 2003). In their review, Hillgarth and Wingfield (1997) listed seven studies of different bird species in which it was found that females preferred the least-parasitized males; however, only two studies concerned blood parasites.
Male Aquatic Warblers with high reproductive success arrived and started copulating earlier and were infected less often by trypanosomes (Table 2), which suggests that one of the reasons for their early arrival may be that they are healthy and not infected by blood parasites. That finding implies that successful males have good immunocompetence, which in turn may reflect their better genetic quality. Although a trade-off between investment in immune system and reproductive effort has been found in birds (Gustafsson et al. 1994, Nordling et al. 1998, Cichoń 2000, Norris and Evans 2000), it mainly occurs in females, whose investment in reproduction is usually higher than that of males. Our results indirectly suggest that males with higher reproductive success should have a stronger immune system. On the other hand, Garamszegi et al. (2004) reported that in the Collared Flycatcher (F. albicollis), immunological treatment with sheep red blood cells triggered a decrease in body mass, testosterone level, and song rate; but other song traits were not significantly affected.
Female birds may assess the genetic quality of males on the basis of their resistance to endoparasities (Hamilton and Zuk 1982). Resistance is correlated with certain secondary sex characters, such as feathers or comb, which are less developed in parasitized individuals. In dull-colored species, one secondary sex character that may be correlated with parasite load is song complexity (Zuk 1984), which should be related to male condition as measured by body weight and fat deposition.
The Aquatic Warbler is not a sexually dimorphic species (and sexual monomorphism is quite unusual in promiscuous species), except that males are slightly but significantly larger than females (Dyrcz 1993). Therefore, coloration cannot be a clue for females to recognize whether a male is infested or uninfested by parasites. However, the most probable explanation is that females simply choose the larger and earlier-arriving males. Infected males may also differ in general behavior from healthy ones. Trypanosomes cause serious diseases in mammals; even if symptoms are weaker in birds, it could be speculated that Aquatic Warbler males with a trypanosome infection are less active and agile than healthy males.
Strong individual differences in mating success between males, as reported here, have also been found in many other birds (e.g. Hötker 1989, Newton 1989), including promiscuous species (e.g. Lill 1974, Alcock 1989). Such differences provide an opportunity for natural selection, and especially sexual selection. Generally, it is assumed that females select males on the basis of genetic quality, which is not directly observable (Kodric-Brown and Brown 1984). The difference in quality may manifest itself indirectly through biometrical characteristics. Here, we show that males with longer wings and larger fat deposits sired more offspring (Table 1). That finding may be attributable to higher paternal success of older males, because they have longer wings than first-breeding young males. Longer wings in older birds have been reported in Reed Warblers (Acrocephalus scirpaceus; Nowakowski 1994, Merom et al. 1999) and Clamorous Reed Warblers (A. stentoreus; Merom et al. 1999). Furthermore, young males are often less successful in obtaining mates than older males (e.g. Harvey et al. 1979, 1985; Perrins and McCleery 1985). We were unable to analyze that assumption directly, because it is difficult to distinguish first-year Aquatic Warbler males from older ones (Svensson 1992). Another possibility is that infected males are in poor condition, and therefore developed shorter wings, though they were not necessarily younger than uninfected birds. Such was the case in a population of Eurasian Blackbirds (Turdus merula) studied in England; adults parasitized with Leucocytozoon sp. were in relatively poor body condition and had shorter wings than unparasitized birds (Hatchwell et al. 2000). In the Sedge Warbler (A. schoenobaenus), males infected with blood parasites weighed less than uninfected ones (Buchanan et al. 1999). Mallards (Anas platyrhynchos) in poorer condition were more likely to be blood-parasitized (Shutler et al. 1999).
Larger fat deposits generally reflect the good physical condition of the male. Our data indicate that males that start breeding early father more offspring than those that start breeding late. Similarly, males of the Great Reed Warbler (A. arundinaceus) that arrive early on breeding grounds more often obtain a second and even a third female (simultaneous facultative polygyny) than males arriving later. Furthermore, polygynous males have higher reproductive success than monogamous ones (Dyrcz 1986, Hasselquist 1998).
In conclusion, our analysis shows strong differences in overall paternal success of individual male Aquatic Warblers. The observed correlations of paternal success with higher fat deposition, an early breeding start, and absence of blood parasites suggest that males in good physical condition have enhanced reproductive success.
Acknowledgments
We thank the Deutsche Forschungsgemeinschaft (Schwerpunktprogram: Genetik von Sozialsystemen) for funding, A. Backhaus for technical assistance, and W. Zdunek for field assistance. The study was also financially supported by a grant from the Polish State Committee for Scientific Research (KBN) and Forschungskommission of the Deutsche Ornithologen-Gesellschaft. We thank two anonymous referees for their constructive criticism.
Literature Cited

{kind=link}