





Abstract
We investigated patterns and causes of variation in clutch sizes in a guild of five species of temperate-nesting dabbling ducks (Mallard [Anas platyrhynchos], Northern Pintail [“pintail,” A. acuta], Gadwall [A. strepera], Blue-winged Teal [“teal,” A. discors], and Northern Shoveler [“shoveler,” A. clypeata]) during 1993-1995 in the Prairie Pothole Region of midcontinental North America. Clutch sizes (mean ± SE) were largest for teal (10.80 ± 0.03), followed in descending order by those of shoveler (10.31 ± 0.05), Gadwall (9.92 ± 0.04), Mallard (8.91 ± 0.04), and pintail (7.66 ± 0.06). In Mallard, pintail, and shoveler, predicted clutch sizes at onset of nesting exhibited minimal variation. Clutch sizes of Gadwall and teal displayed statistically significant variation among years at onset of nesting; pintail clutch sizes showed significant variation late in the nesting season. Clutch sizes declined seasonally in all species. Declines in clutch sizes of teal and shoveler were approximately linear; whereas clutch sizes of Mallard, pintail, and Gadwall usually declined at progressively decreasing rates. Linear declines in teal and shoveler clutches suggest that those species experienced greater difficulty securing lipids for egg production late in the nesting season than did Mallard, pintail, and Gadwall. That disparity may result because egg-laying female teal and shoveler feed almost exclusively on animal foods, which are primarily protein; whereas female Mallard, pintail, and Gadwall consume more carbohydrate-rich plant foods. Our findings, when examined in context with existing information, suggest that interspecific variation in clutch sizes results from innate differences in several traits—including body size, diet, timing of lipid acquisition, and nesting—all of which can affect the amount of lipid available for egg production. Temperate-nesting dabbling ducks have evolved traits that facilitate laying of large clutches early in the nesting season, because risk of mortality is lower among early-hatched young. Annual differences in clutch sizes of all five species were not significant when effects of annual variation in nest-initiation dates were accounted for, reflecting the key role of environmental influences on intraspecific variation in clutch sizes among years.
RESUMEN
Investigamos los patrones y las causas de la variación en el tamaño de la nidada en cinco especies de patos de la tribu Anatini que nidifican en la zona templada (Anas platyrhynchos, A. acuta, A. strepera, A. discors y A. clypeata) entre 1993-1995 en la región de Prairie Pothole en el centro de Norte América continental. Los tamaños de la nidada (media ± EE) fueron mayores para A. discors, seguidos en orden decreciente por A. clypeata (10.31 ± 0.05), A. strepera (9.92 ± 0.04), A. platyrhynchos (8.91 ± 0.04) y A. acuta (7.66 ± 0.06). En A. platyrhynchos, A. acuta y A. clypeata, los tamaños de las nidadas predichos al inicio de la nidificación presentaron variaciones mínimas. Los tamaños de las nidadas de A. strepera y A. discors mostraron variaciones estadísticamente significativas entre años en el momento del inicio de la nidificación, y los tamaños de las nidadas de A. acuta mostraron variaciones significativas tardíamente en el período de nidificación. Los tamaños de las nidadas disminuyeron estacionalmente en todas las especies. Las disminuciones en A. discors y A. clypeata fueron aproximadamente lineales, mientras que los tamaños de las nidadas de A. platyrhynchos, A. acuta y A. strepera generalmente disminuyeron progresivamente. La disminución lineal en las nidadas de A. discors y A. clypeata sugiere que estas especies presentaron una mayor dificultad para mantener los niveles de lípidos necesarios para la producción de huevos a fines de la época de nidificación, en comparación con A. platyrhynchos, A. acuta y A. strepera. Esta disparidad podría deberse a que las hembras de A. discors y A. clypeata que están poniendo huevos consumen casi exclusivamente alimentos animales, los cuales principalmente proveen proteína, mientras que las hembras de A. platyrhynchos, A. acuta y A. strepera consumen más alimentos vegetales ricos en carbohidratos. Si examinamos nuestros resultados en el contexto de la información existente, éstos sugieren que las diferencias interespecíficas en el tamaño de las nidadas son el resultado de diferencias innatas en varios caracteres - incluyendo tamaño corporal, dieta, momento en que se adquieren los lípidos, nidificación - que afectan la cantidad de lípidos disponibles para la producción de huevos. Los patos de la tribu Atini que nidifican en las zonas templadas han evolucionado caracteres que facilitan la puesta de nidadas grandes tempranamente en la época de nidificación, debido a que el riesgo de mortalidad de los juveniles eclosionados tempranamente es menor. Las variaciones anuales en los tamaños de las nidadas de las cinco especies no fueron significativas cuando se consideró el efecto de la variación anual en la fecha de iniciación del nido, reflejando el rol clave de las influencias ambientales en la variación interespecífica en el tamaño de las nidadas entre años.
Although much information has been published on clutch-size relationships among temperate-nesting ducks (Rohwer 1992), causes of interspecific and intraspecific variation remain poorly understood. Insights have been limited because clutch-size comparisons usually involve different geographic regions and environmental conditions. Therefore, it has been difficult to distinguish variation in clutch sizes caused by innate influences from variation caused by environmental influences. As a result, effects of major innate and environmental influences on clutch sizes remain unclear.
Female nutrition has long been suspected of influencing clutch sizes (Lack 1967, 1968); however, detailed knowledge concerning the influence of nutrition on clutch sizes in wild populations of dabbling ducks is lacking. Over the past two decades, much insight has been gained regarding sources of nutrients that female ducks rely on to produce clutches. Female dabbling ducks (genus Anas) breeding in temperate North America acquire most of the protein and calcium used in egg production from macro-invertebrates obtained from shallow wetlands on breeding grounds (Krapu and Reinecke 1992). Lipid for egg production is acquired from plant and animal foods and stored as an endogenous reserve either prior to or after arrival at the breeding site, or obtained directly from the diet during the egg-laying period. In several species of temperate-nesting dabbling ducks, amounts of endogenous lipid that females commit to egg production are related to the size of lipid reserves (Alisauskas and Ankney 1992). Female Mallard (Anas platyrhynchos) with large lipid reserves are capable of laying more eggs than those with small reserves, and clutch sizes of lighter females may be lipid-limited (Arnold et al. 2002). However, innate and environmental influences on amounts of lipid available for egg production, and thus on clutch size, are little understood. That lack of knowledge limits our understanding of the causes of inter- and intraspecific variation in clutch sizes.
We hypothesized that by comparing patterns of variation in clutch sizes among several species of prairie-nesting dabbling ducks across a range of wetland habitat conditions over several years, we potentially could distinguish between variation in clutch sizes caused by innate and environmental influences. To minimize sources of environmental variation within years, we measured clutch sizes of studied species in the same landscapes. We chose a guild of five species of prairie-nesting dabbling ducks: Mallard; Northern Pintail (A. acuta), hereafter “pintail”; Gadwall (A. strepera); Blue-winged Teal (A. discors), hereafter “teal”; and Northern Shoveler (A. clypeata), hereafter “shoveler.” All five species are common breeders in the Prairie Pothole Region (PPR) of midcontinental North America (Bellrose 1980). To evaluate the extent of interspecific variation in clutch sizes, we compared clutchsize relationships among the five species concurrently, in the same prairie-pothole landscapes, during the 1993-1995 nesting seasons. To assess influence of wetland conditions on intraspecific variation in clutch sizes, we compared clutch sizes by date within and among years as precipitation and pond conditions changed. Specifically, we (1) documented sizes of completed clutches and estimated dates of nest initiation from the onset to the end of most nesting for Mallard, pintail, Gadwall, teal, and shoveler in the PPR of North Dakota and South Dakota; (2) estimated mean clutch sizes by date and year for each of the five species; (3) compared patterns of inter- and intraspecific variation in clutch sizes within and among years; and (4) evaluated effects of innate and environmental influences on inter- and intraspecific variation in clutch sizes.
Study Areas and Methods
Location of study areas
Clutch-size data were collected during spring and early summer of 1993-1995 on 10-km2 study sites located in the PPR of North Dakota and South Dakota (Fig. 1; Kantrud et al. 1989a). Clutches were located by searching fields enrolled by private landowners in the Conservation Reserve Program (CRP) and on Waterfowl Production Areas (WPAs) owned and managed by the U.S. Fish and Wildlife Service. Each field searched contained ≤16.2 ha of CRP grassland cover. Numbers of plots contributing data per year were 37 (1993), 46 (1994), and 44 (1995). Nests were located and marked using standard nest-searching techniques (Klett et al. 1986). Each field selected for study was searched three times at ∼21-day intervals between 1 May and 2 July each year. Nests were revisited on subsequent searches or more frequently to determine fates (Reynolds et al. 2001). Only clutches monitored into incubation (Weller 1956) and showing no evidence of eggs having been removed by predators, accidentally broken by the female, or destroyed from other causes were included in the sample.
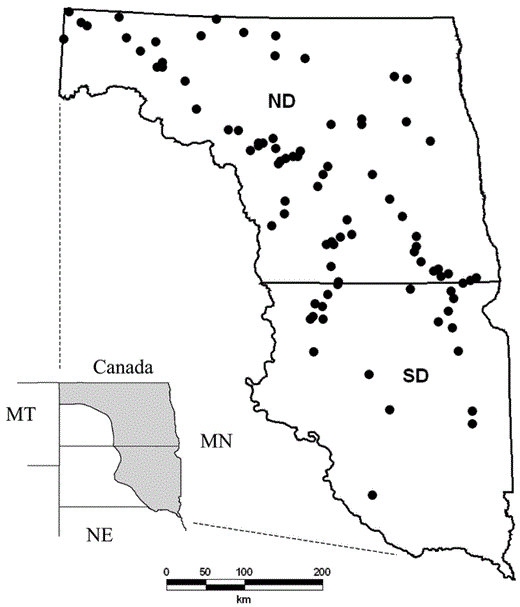
Black dots indicate study locations where data on completed clutch sizes of five species of temperate-nesting dabbling ducks were collected in prairie pothole landscapes east of the Missouri River in North Dakota and South Dakota during 1993-1995.
Statistical analysis
We used the lme() function of S-PLUS for Windows (version 6.1, Insightful Corporation, Seattle, Washington) to perform a mixed-model analysis of variance (ANOVA) and estimate mean clutch sizes for each species. Our model included species as a fixed classification factor and year as a random factor; the random effect of year nested within species served as the error term for testing the main effect of species (see Yandell 1997, Pinheiro and Bates 2000). After finding evidence of differences among species, we used one-way ANOVA, with year as a fixed factor, to compare clutch sizes among years for each species. When we found significant differences among years, we used Fisher's least-significant difference (LSD) test to conduct multiple comparisons. After fitting ANOVA models, we used residual plots to inspect distributions and variances of residuals. We considered model assumptions to be met adequately if residuals were symmetrically distributed about means and heteroscedasticity was not evident.
After testing for differences in mean clutch sizes, we used multiple-regression models to describe variation in mean clutch sizes within breeding seasons. Terms included in candidate models were date (linear and quadratic) and year, as well as interactions that allowed seasonal patterns to vary annually. Furthermore, because differences in nesting chronology are well documented (Krapu 2000), we fit a separate model for each species. Because clutch sizes are discrete, we also experimented with Poisson regression models; however, Poisson models were unsatisfactory because variances of clutch sizes were not proportional to means.
To assure that results were robust to criteria used for model selection, we compared models selected via Akaike's information criterion (AIC; Burnham and Anderson 1998) and via backward elimination with F-tests and an alpha level of 0.05 (Chatterjee and Price 1991). We present only the results of model selection by AIC because the two approaches, with a few exceptions that we note, identified the same “best” models. After selecting models, we examined residual plots to evaluate model fit, confirm homoscedasticity, and check for outliers or observations with undue influence. We used the lm() procedure of S-PLUS for model selection.
Mean clutch sizes at the onset and late in nesting are particularly informative, because they reflect the relative availability and sources of nutrients used in egg production. We therefore estimated a “nesting period” that was bounded by the three-year medians for earliest- and latest-observed nest-initiations. We then used model averaging (Burnham and Anderson 1998) to estimate mean annual clutch sizes and standard errors for beginning dates of those periods.
Annual differences in mean clutch size could have arisen from—or been obscured by—differences in nest-initiation dates, annual variation in other factors, or both influences acting in concert. To gain insights about consequences of annual changes in nest-initiation dates, we compared arithmetic-mean clutch sizes with model-averaged estimates of mean clutch sizes that would have resulted from constant nesting effort. To obtain constant-effort estimates for each model in our candidate set, we averaged the predicted values for dates in the nesting period. We used nonparametric bootstrapping (Efron and Tibshirani 1993) to estimate standard errors for average predicted values from each model.
Results
Precipitation and Wetland Habitat Conditions
The study began in spring 1993 at the onset of recovery from a five-year (1988-1992) drought in the northern Great Plains (Winter and Rosenberry 1998). Precipitation remained below average across North Dakota during April 1993 at the onset of nesting (National Oceanic and Atmospheric Administration [NOAA] 1992-1995). Runoff from snowmelt in early spring was sufficient to provide some water in many shallow basins, but most semipermanent and many seasonal ponds remained dry through April (G. Krapu pers. obs.). Precipitation increased in May, and June and July were extremely wet in North Dakota and South Dakota (United States Army Corps of Engineers 1994); rainfall averaged 4.9 and 13.6 cm above average across North Dakota during June and July, respectively (NOAA 1993). As a result, number of ponds increased from May to July 1993 across parts of North Dakota and South Dakota where study plots were located (Table 1). The increasing number of ponds from May to July in the PPR was unusual, because ponds typically decline during that period as ambient temperatures rise and precipitation declines from spring to summer (Kantrud et al. 1989a). Number of ponds continued to increase in spring 1994 and 1995 (Table 1), because precipitation remained well above average (NOAA 1994-1995). Landscape densities of settled pairs of dabbling ducks increased from 1993 to 1995 on the study area as number of ponds increased (Table 1). Fewer breeding pairs of all five species were present in May 1993 than in May 1994 and 1995 (Table 1)—presumably, in part, because pond densities in April, when pairs arrive in the region, were lower in 1993 than in 1994 and 1995.
Numbers of May and July ponds and numbers of dabbling ducks in strata 45 and 46 (North Dakota) and 48 (South Dakota) during the May Annual Breeding Pair and Pond Survey of the U.S. Fish and Wildlife Service (2001)
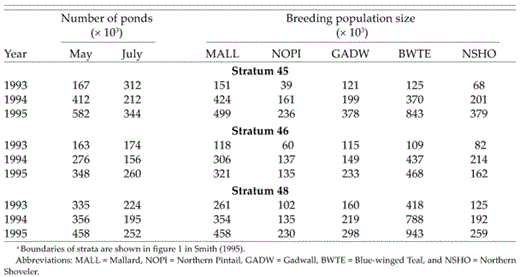
Numbers of May and July ponds and numbers of dabbling ducks in strata 45 and 46 (North Dakota) and 48 (South Dakota) during the May Annual Breeding Pair and Pond Survey of the U.S. Fish and Wildlife Service (2001)
Patterns of Variation in Clutch Sizes
Mean clutch sizes
Mean clutch sizes varied among species (F = 564, df = 4 and 8, P < 0.0001) and were largest for teal, followed by shoveler, Gadwall, Mallard, and pintail (Table 2). A majority of annual variation was additive (σ̂Year = 0.104, σ̂Year×Species = 0.022), reflecting a substantial degree of synchrony among species.
Arithmetic-mean clutch sizes (x̄ ± SE) of Mallard, Northern Pintail, Northern Shoveler, Blue-winged Teal, and Gadwall during the 1993-1995 nesting seasons in the Prairie Pothole Regions of North Dakota and South Dakota. Sample sizes are in parentheses.
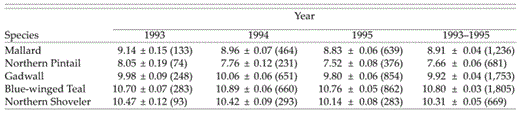
Arithmetic-mean clutch sizes (x̄ ± SE) of Mallard, Northern Pintail, Northern Shoveler, Blue-winged Teal, and Gadwall during the 1993-1995 nesting seasons in the Prairie Pothole Regions of North Dakota and South Dakota. Sample sizes are in parentheses.
Intraspecific differences in clutch sizes among years, which provide potential insight into possible causes of variation, were at least marginally statistically significant for every species (teal: F = 2.29, df = 2 and 1,802, P = 0.10; Gadwall: F = 4.61, df = 2 and 1,750, P = 0.02; Mallard: F = 2.29, df = 2 and 1,233, P = 0.10; pintail: F = 3.83, df = 2 and 678, P = 0.02; shoveler: F = 3.51, df = 2 and 666, P = 0.03). Arithmetic-mean clutch sizes of Mallard and pintail were largest in 1993 and smallest in 1995 (Table 2; Fig. 2). Arithmetic-mean clutch sizes of teal were similar in 1993 and 1995, and largest in 1994 (Table 2; Fig. 2). Gadwall clutches were larger in 1994 than in 1995, and intermediate in 1993 (Table 2; Fig. 2). Shoveler clutch sizes were similar in 1993 and 1994, and smaller in 1995 (Table 2; Fig. 2). In all five species, observed differences among years were substantially reduced and did not approach statistical significance, after we adjusted for the influence of annual differences in nest-initiation dates (Fig. 2).

Results of multiple comparisons of annual mean clutch sizes for five species of dabbling ducks nesting in the Prairie Pothole Region of North Dakota and South Dakota, 1993-1995. Symbols represent differences between arithmetic means (O), with associated 90% confidence intervals, and differences remaining after adjustment for influence of nest initiation dates (×). Confidence intervals that do not overlap 0 indicate statistical significance at P = 0.10.
In Mallard, pintail, and shoveler, predicted clutch sizes at onset of nesting exhibited minimal annual variation (Table 3). In Gadwall (z = −1.96; P = 0.05) and teal (z = −2.62; P = 0.008), predicted clutch sizes at the onset of nesting were smaller in 1993 than in 1995 and intermediate in 1994. Clutch sizes of teal were significantly smaller (z = −2.16; P = 0.03) in 1993 than in 1994.
Model-averaged estimates of mean clutch sizes (± SE) of Mallard, Northern Pintail, Gadwall, Blue-winged Teal, and Northern Shoveler at the median Julian date of the onset of nesting in the Prairie Pothole Region of North Dakota and South Dakota during 1993, 1994, and 1995.
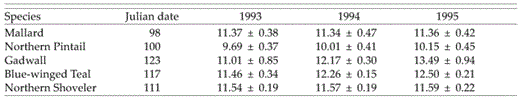
Model-averaged estimates of mean clutch sizes (± SE) of Mallard, Northern Pintail, Gadwall, Blue-winged Teal, and Northern Shoveler at the median Julian date of the onset of nesting in the Prairie Pothole Region of North Dakota and South Dakota during 1993, 1994, and 1995.
Seasonal variation in clutch sizes
Except for teal, AIC and backward elimination identified the same “best” models. For teal, AIC suggested similar support for three models, including the one selected by backward elimination. That fact, and symmetrically distributed, homoscedastic residuals suggest that our assessment of clutch sizes was robust to the method used for model selection. Because both methods produced equivalent results, we present only results based on AIC (Table 4). Patterns of seasonal mean clutch sizes estimated by our best-fitting models and model-averaged estimates of parameters representing main effects of initiation dates varied among species (Fig. 3).
Results of model selection describing seasonal changes in clutch sizes of Mallard, Northern Pintail, Gadwall, Blue-winged Teal, and Northern Shoveler during spring and early summer of 1993-1995 in the Prairie Pothole Region of North Dakota and South Dakota. Coefficients represent intercepts (α) and linear (β) and quadratic (γ) effects of nest-initiation dates. Coefficients with subscripts are year-specific (i.e. αi denotes a separate intercept for each year). Akaike weights (Burnham and Anderson 1998) used in model averaging are represented by wj: entries of 0 denote weights ≤0.005. Models with a dominant influence on results are in bold.
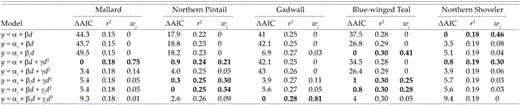
Results of model selection describing seasonal changes in clutch sizes of Mallard, Northern Pintail, Gadwall, Blue-winged Teal, and Northern Shoveler during spring and early summer of 1993-1995 in the Prairie Pothole Region of North Dakota and South Dakota. Coefficients represent intercepts (α) and linear (β) and quadratic (γ) effects of nest-initiation dates. Coefficients with subscripts are year-specific (i.e. αi denotes a separate intercept for each year). Akaike weights (Burnham and Anderson 1998) used in model averaging are represented by wj: entries of 0 denote weights ≤0.005. Models with a dominant influence on results are in bold.
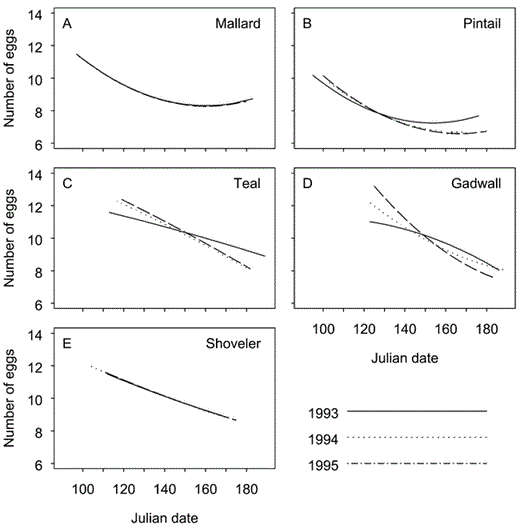
Model-averaged estimates of mean clutch sizes, by nest-initiation date, for (A) Mallard, (B) Northern Pintail (Pintail), (C) Gadwall, (D) Blue-winged Teal (Teal), and (E) Northern Shoveler (Shoveler) in the Prairie Pothole Region of North Dakota and South Dakota during 1993-1995.
Seasonal decline in clutch sizes of Mallard was best described by a simple quadratic relation. Simple linear models received virtually no support, and we found little evidence of annual variation (Table 4). In contrast, our data provided similar support for three models describing clutch sizes of pintail (Table 4). Those included a simple quadratic model and two models that allowed annual variation in relations between clutch sizes and nest-initiation dates.
For shoveler, our data provided similar support for simple linear and simple quadratic models. For Mallard, we found little evidence of annual variation in the relationship between initiation dates and clutch sizes. In contrast, competing models for teal and the best-supported model for Gadwall all included annual differences in slope coefficients and intercepts (Table 4).
For pintail and teal, relations between initiation dates and mean clutch sizes estimated by model averaging were similar for 1994 and 1995. Support for models accommodating annual variation thus resulted from relatively smaller mean clutch sizes early in 1993, and larger mean clutch sizes later. Results for Gadwall suggested similar differences between 1993 and 1995; however, results for 1994 were intermediate between estimates for 1993 and 1995 and likely contributed to the relatively strong support for a single, more complex model.
Discussion
Causes of Interspecific Variation in Clutch Sizes
Timing of nesting
Innate differences among the five species in timing of nesting (Krapu 2000) contributed to interspecific variation in mean clutch sizes. In North Dakota and South Dakota, female pintail and Mallard initiate nesting in early spring under lower ambient temperatures than Gadwall, teal, and shoveler (Hammond and Johnson 1984), resulting in higher percentages of endogenous lipids being used to meet maintenance energy requirements (Kendeigh et al. 1977). Low water temperatures in early spring also result in lower standing crops of macro-invertebrates in prairie wetlands than after conditions become more moderate (Murkin and Ross 2000). Consequently, highquality food is less available to female Mallard and pintail than to later nesters. Relatively small clutch sizes of Mallard and pintail may reflect the combined effects of relatively high maintenance requirements and low rates of lipid intake from diet during egg production. The progressively slower rates of decline in clutch sizes of Mallard and pintail as spring advanced were probably influenced by less energy being required for maintenance; also ambient temperatures rose from early to late spring (NOAA 1993-1995) (Kendeigh et al. 1977), causing a higher proportion of endogenous and dietary lipid to become available for egg production.
Body size
Of species we studied, female Mallard have the largest prelaying body masses and, thus, the lowest relative metabolic rate (Table 5). A higher ratio of body mass to clutch mass at the onset of nesting enables female Mallard to draw on endogenous lipids for a greater proportion of clutch lipid requirements. Predicted mean clutch sizes at onset of nesting were exceptionally consistent among years for Mallard, presumably reflecting that the amount of endogenous lipid females carried to the breeding grounds did not vary widely among years. Territorial behavior, large home ranges (Anderson and Titman 1992), and capacity of nesting females to acquire nutrients from a wide range of habitats (Krapu et al. 1997) may have contributed to Mallard having similar rates of decline in clutch sizes during 1993-1995, despite annual variation in wetland habitat conditions on their breeding grounds (also see Pietz et al. 2000). As expected, female pintail laid smaller early clutches than Mallard, probably because pintail have higher metabolic rates and greater maintenance energy requirements, relative to their body size and lipid reserves (Table 5). Female pintail thus use a relatively high percentage of their endogenous lipid to produce large early clutches (Esler and Grand 1994).
Prelaying body mass in relation to average mass of initial clutch and the body mass:clutch mass ratio for five species of temperate-nesting dabbling ducks.
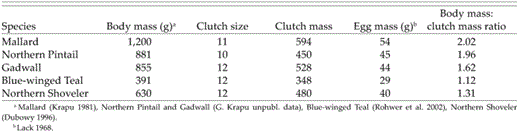
Prelaying body mass in relation to average mass of initial clutch and the body mass:clutch mass ratio for five species of temperate-nesting dabbling ducks.
Diet
Steady declines in clutch sizes of teal and shoveler during the nesting season probably occurred because egg-laying females forage almost exclusively on protein-rich animal foods—macro-invertebrates account for 99% of the diets of egg-laying female teal and shoveler (Swanson et al. 1979)—reducing their intake of lipid and lipid precursors. Carbohydrate-rich plant foods (important lipid precursors) form a significant part of the diets of egg-laying female Mallard, pintail, and Gadwall (Krapu and Reinecke 1992) and likely contributed to progressively decreasing rates of decline in clutch sizes.
Ages of females
Declines in arithmetic-mean clutch sizes of pintail and Mallard from 1993 to 1995 may have been influenced by higher ratios of adult to yearling females in their breeding populations in 1993. Lower ratios of immature to adult female Mallard and pintail in the harvest in the Central and Mississippi flyways in 1992 than in 1993 and 1994 (Sharp and Moser 2000) suggest lower percentages of yearling females in the breeding populations in spring 1993. Yearling female Mallard and pintail breeding in the PPR have lower body masses, initiate nesting later, and lay smaller initial clutches than adults (Krapu and Doty 1979, Duncan 1987, Young 1993). Although mean clutch sizes were ordered as we expected in 1993-1995 for Mallard and pintail, differences between annual estimates were relatively small and not statistically significant. Small differences may indicate that prolonged renesting in 1993 (Krapu et al. 2001) resulted in sufficient numbers of late (and smaller) clutches to offset larger clutch sizes early in the nesting season. Mallard nest longer in wet than in dry springs (Krapu et al. 1983), in part, because a greater abundance of high-quality food increases protein and lipid intake, increasing the potential for continued nesting after most endogenous lipid reserves are depleted.
Adaptations for Laying Large Early Clutches
Despite their diverse life histories, all five species consistently laid relatively large clutches early in the nesting season. Large early clutches are adaptive in the PPR because of higher survival of early-hatched ducklings in most years (Rotella and Ratti 1992, Dzus and Clark 1998, Guyn and Clark 1999). Seasonal ponds are usually most plentiful in spring and early summer (Kantrud et al. 1989b), and the risk of duckling mortality declines as the percentage of seasonal basins containing water increases (Krapu et al. 2000, Pietz et al. 2003), except where permanent water is plentiful (Krapu et al. 2004). Because young that hatch early survive at relatively high rates, traits have evolved that ensure females have the potential to acquire sufficient lipid to produce large clutches early in the nesting season. Female Mallard and pintail overcome constraints that would otherwise limit production of large clutches in early spring by acquiring lipid reserves before they arrive on the breeding grounds (Krapu 1974, 1981; Esler and Grand 1994). In midcontinental North America, Mallard and pintail acquire lipid reserves during stopovers at key sites along their spring migration routes, for example, the Rainwater Basin Area in south-central Nebraska (Jorde 1981, R. R. Cox, Jr. unpubl. data). Gadwalls begin arriving on their breeding grounds in North Dakota as early as late March (Dwyer 1974) but do not start nesting until about 1 May (Krapu 2000). Between arrival and onset of nesting, female Gadwall store endogenous lipid (G. Krapu unpubl. data), increasing their capacity to lay large initial clutches. Teal lay large early clutches, despite their small body sizes (and reduced capacity to store lipid), by settling in landscapes where high densities of recently filled temporary and seasonal ponds (Drewien and Springer 1969, Johnson and Grier 1988) provide exceptionally productive foraging habitat. Also, by nesting late, teal reduce the energy required for maintenance. Female shoveler produce large clutches in early spring primarily from endogenous lipid (Ankney and Afton 1988) gained from foraging on crustaceans and gastropods (Swanson et al. 1979, Ankney and Afton 1988). Cladocerans, the principal crustacean consumed by egglaying female shoveler (Swanson et al. 1979), are among the first macro-invertebrates to become abundant in spring (DuBowy 1988) and are at their peak caloric content early in the season (Wissing and Hasler 1971). Also, male shoveler aggressively defend territories during the laying period (McKinney 1973), helping females to forage undisturbed and thus increase their intake of nutrients needed for egg production.
Causes of Intraspecific Variation in Clutch Sizes
Within waterfowl populations, onset of nesting and sizes of initial clutches have been shown to vary with body mass (Reynolds 1972) and amounts of endogenous lipid that individual females carry at the onset of nesting (Alisauskas and Ankney 1992, Esler and Grand 1994). Females entering the nesting season with large lipid reserves apparently can mobilize sufficient lipids from reserves to initiate rapid follicular growth earlier and lay larger clutches than those with smaller lipid reserves. In springs when ambient temperatures are relatively warm, female dabbling ducks nest earlier (Hammond and Johnson 1984) and lay larger clutches. Similarly, as spring advances, increasing ambient temperatures and declining maintenance requirements allow females with less endogenous lipid to initiate nesting. Female Mallard and pintail that arrive on the breeding grounds with reduced amounts of lipid nest later, lay smaller clutches, and hatch fewer ducklings that are at higher risk of mortality in most years, because of declining quality of wetland habitat used for brood-rearing. As a result, foraging conditions that female Mallard and pintail encounter during spring migration influence the amount of lipid stored and, thus, reproductive success.
Although our results are based on observational data, they match prior expectations that took into account that Mallard and pintail produce early clutches mostly from endogenous lipid acquired before arrival on the breeding grounds, whereas Gadwall and teal acquire a major part of lipid for egg production after arrival. Related to that difference, water conditions on the breeding grounds had a greater influence on clutch sizes of Gadwall than on Mallard (also see Pietz et al. 2000). Pintail, like Mallard, experienced minimal annual variation in clutch sizes early in the nesting season across a wide range of water conditions. However, pintail clutches laid in late spring 1993 were larger than in following years, which suggests that after endogenous lipid is depleted, pintail clutch sizes are sensitive to local wetland conditions. Previous research has shown that rates of decline in pintail clutch sizes vary from temperate to Arctic regions—an apparent result (in part) of differences in maintenance energy costs and amount of lipid available from the diet during egg production (Krapu et al. 2002). Teal, like Gadwall, acquire a major part of their lipid for egg production, both early and late, from wetlands on the breeding grounds; that helps to explain why teal clutch sizes were smaller early and larger late in the nesting period in 1993 than in 1994 and 1995.
Seasonal declines in clutch sizes of pintail, Gadwall, and teal occurred at slower rates in 1993 than in 1994 and 1995, coinciding with the filling of temporary and seasonal wetland habitat in late spring of 1993 after a prolonged severe drought. Aquatic macro-invertebrates, the primary source of protein and calcium for females of the five species during egg production, increase rapidly following the filling of previously dry seasonal ponds (Neckles et al. 1990), and carbohydrate-rich seeds produced while wetlands were dry also become available, creating exceptionally productive foraging conditions. Teal, Gadwall, and Mallard usually terminate most nesting by the end of spring, but nested into late summer in 1993 (Krapu et al. 2001), reinforcing the conclusion that foraging conditions in late spring and summer 1993 were exceptionally productive for prairie-nesting ducks. For all five species, annual differences in clutch sizes were reduced and no longer significant when effects of annual variation in nest-initiation dates were accounted for, further supporting the hypothesis that environmental factors were a dominant influence in controlling intraspecific variation in clutch sizes of pintail, Gadwall, and teal during 1993-1995. Our conclusion that environmental factors serve a key role in determining intraspecific variation in waterfowl clutch sizes supports the prediction of Ankney and Afton (1988) that future studies of waterfowl would show that most intraspecific variation in mean clutch sizes is due to environmental variation.
Acknowledgments
Funding for the study was provided by the Central Flyway Council; Ducks Unlimited Incorporated's Institute for Wetland and Waterfowl Research; Mississippi Flyway Council; U.S. Bureau of Reclamation, Billings, Montana; U.S. Fish and Wildlife Service, Denver, Colorado; and U.S. Geological Survey, Northern Prairie Wildlife Research Center. The Wildlife Management Institute, Washington, D.C., managed fund accounts for some contributors. U.S. Fish and Wildlife Service managers M. D. Blenden, F. G. Geise, R. A. Gilbert, H. J. Hoistad, R. A. Hollevoet, R. L. Howard, S. J. Kresl, D. G. Potter, R. J. Vanden Berge, P. C. Van Ningen, and D. T. Walls, and their staffs provided logistical support. We thank D. A. Brandt and B. Euliss for assistance in preparing Figure 1, and D. A. Brandt for organizing data files for analyses. We are indebted to numerous field-crew personnel whose dedicated efforts made the study possible. We much appreciate constructive comments provided by C. D. Ankney, T. W. Arnold, R. R. Cox, Jr., D. A. Haukos, K. J. Reinecke, and W. E. Newton on earlier drafts of the manuscript. Finally, we are grateful to the many landowners who granted access to their lands.
Literature Cited

{kind=link}
{kind=link}
{kind=link}