





Large mammalian herbivores made the world spiny. Over evolutionary time scales, the rise of physically defended plants – species that produce spines, thorns or prickles – has been linked to the diversification of large herbivores. In African savannas, for example, spinescence has arisen at least 55 times across diverse plant lineages, largely in response to, and concurrent with, the diversification of large-bodied browsers (Charles-Dominique et al., 2016; Fig. 1). The contemporary distribution of browsing mammals likewise drives conspicuous variation in plant defence phenotype across landscapes, such that plant communities in areas of high herbivore density tend to be more spiny than those lacking herbivores; these patterns are caused by both selection against undefended species where herbivores are abundant and the induction of defences in spinescent species following browsing (Ford et al., 2014; Coverdale et al., 2018). Ecological and evolutionary patterns thus suggest that mammalian herbivores have driven the evolution of spinescence, and that spines, thorns and prickles, in turn, are effective deterrents of large browsers. Defence phenotype can shift markedly over plant ontogeny, however, and many conspicuously spinescent species lack physical defences altogether for long stretches of their development (Dayrell et al., 2018); seedlings and saplings of these species, whose deployment of defences may be delayed by resource limitation or physiological constraints, are particularly vulnerable to browsers (Staver and Bond, 2014). In this issue of the Annals of Botany, Armani et al. (2019) explore the emergence of spines, thorns and prickles during this critical stage and find that not all defences are created equal.
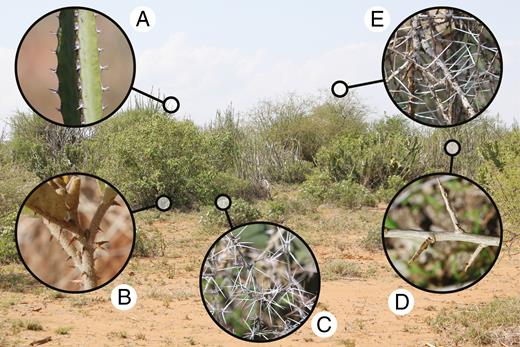
Diversity of physical plant defences in an East African savanna (Mpala Research Centre, Laikipia, Kenya). Generally considered to have evolved in response to large mammalian browsers, physical plant defences are often lumped into a single defence strategy (‘spinescence’). In a study published in this issue, Armani et al. (2019) demonstrate that the timing of defence emergence during plant ontogeny is strongly constrained by defence type, suggesting that spines, prickles and thorns are not as interchangeable as previously thought. Insets: (A) Euphorbia spp. (spines); (B) Solanum campylacanthum (prickles); (C) Barleria trispinosa (spines); (D) Lycium europaeum (thorns); (E) Acacia etbaica (spines). Physical defences range from approx. 2 mm (e.g. inset A) to approx. 80 mm (e.g. inset E) in length.
Traditionally, ecologists have not distinguished between spines, thorns and prickles, instead lumping them into a single defence strategy (‘spinescence’) based on their superficially similar morphology (Fig. 1). This broad treatment has been sufficient to test basic hypotheses about their evolutionary origins and efficacy against herbivores, but a more nuanced approach to the study of physical defences is long overdue. Indeed, the relative paucity of studies investigating potential ecological, evolutionary and/or physiological differences between spines, thorns and prickles is a key reason why our understanding of physical defences has lagged behind that of chemical defences: exploring the causes and consequences of phytochemical diversity has long been a cornerstone of chemical ecology. Given that spines, thorns and prickles arise from different plant organs (leaves, branches, and epidermal or cortex tissue, respectively), for example, it is reasonable to assume that the timing of their emergence during early plant ontogeny may be constrained by the development of those plant parts: spines may emerge earliest in conjunction with the first growth of leaves, prickles shortly thereafter due to a slightly longer developmental period, and thorns only after primary stem growth and branching. In their study, Armani et al. (2019) assessed the timing of defence emergence across 45 spinescent species grown in a common garden setting, and found strong support for different developmental trajectories across spinescent plants. As predicted, spine-bearing species produced defences first, followed closely by those producing prickles. Thorny species, in contrast, regularly experienced a long delay prior to defence emergence: more than half of thorn-bearing saplings did not produce a single thorn within 15 weeks of germination.
A key result from the study of Armani et al. (2019) then is the unequivocal demonstration that spines, thorns and prickles vary substantially in their regulation and expression during early ontogeny. In this regard, the authors’ findings complement earlier work on the ontogeny of chemical and biological (e.g. myrmecophily) defences, which show similar delays while necessary plant organs or tissues develop and grow. Collectively, these results suggest that ontogenetic shifts in defence expression are common (and perhaps unavoidable) across diverse defence strategies. Given the particular vulnerability of seedlings and sapling to browsers during this period, the authors suggest that variation in defence type (and thus the timing of defence emergence) may shape patterns of plant diversity and abundance in ecosystems with large herbivores. Whether differences in physical defence expression during the bottleneck of early ontogeny shape patterns of adult plant diversity and abundance remains an open question, but one deserving of further investigation, particularly in light of these results and ongoing, continental-scale shifts in herbivore communities. Large herbivore exclosure experiments in areas of high herbivore density and diversity such as East Africa (Goheen et al., 2018) may be particularly useful in this pursuit.
Another intriguing aspect of the study of Armani et al. (2019) is their assessment of the relative cost of different defence types. The authors collected and weighed all spines, thorns and prickles at the conclusion of their experiment and, based on these results, one may be tempted to draw several conclusions. For example, it appears that spines and prickles may represent two distinct strategies (or ‘syndromes’, sensuAgrawal and Fishbein, 2006) for deterring herbivores during early plant development: spines emerge rapidly but are more costly in terms of resource requirements and lost photosynthetic capacity, while prickles emerge more slowly but incur lower construction and opportunity costs. Given the ubiquity of trade-offs between growth and defence, this supposition seems both reasonable and testable (Dayrell et al., 2018). Comparative approaches across spinescent species or genera may be a particularly fruitful approach in the search for signatures of physical defence syndromes. More difficult to explain is the strategy of thorny species, which appear to endure the longest period without physical deterrents while incurring relatively high cost in terms of both thorn production and lost photosynthetic capacity. Broad comparisons of physical defence strategies – including estimates of the costs of producing and maintaining different defences and the vulnerability of individuals during undefended periods – is a promising area for future research (Staver and Bond, 2014). In particular, efforts in this area should follow the lead of earlier investigations of chemical defences by seeking to identify ecological, evolutionary and physiological trade-offs associated with different physical defence strategies (Agrawal and Fishbein, 2006).
Studies of plant defences against herbivores have been a mainstay of the field of ecology for decades (Burkepile and Parker, 2017). By clearly demonstrating that spines, thorns and prickles are not interchangeable – at least during critical seedling and sapling stages – Armani et al. (2019) have made a significant contribution to our general understanding of plant defence strategies. Theirs is among the first studies to document meaningful differences among physical defence types, and I hope that it will serve as a roadmap for future investigations of spinescent plants.

{kind=link}